
NlMA related kinase 2 promotes gastric cancer cell proliferation via ERK/MAPK signaling
2019-07-10 06:27WeiDongFanTaoChenPengJunLiu
World Journal of Gastroenterology 2019年23期
Wei-Dong Fan, Tao Chen, Peng-Jun Liu
Abstract BACKGROUND NIMA related kinase 2 (NEK2) is closely related to mitosis, and it is currently considered to be over-expressed frequently in many poorly prognostic cancers.However, the effect of the up-regulated NEK2 on cellular signaling in tumors,such as gastric cancer (GC), is con-fusing.AIM To determine the role of the up-regulation of NEK2 in GC.METHODS To investigate the pathological significance of NEK2 in GC, the expression pattern of NEK2 in GC was investigated based on the “Oncomain” database and compared between 30 pairs of cancer samples and adjacent tissues. The coexpression of NEK2 and ERK in GC was analyzed using The Cancer Genome Atlas (TCGA) database and confirmed in clinical samples by quantitative realtime PCR (qRT-PCR), and the survival curve was also plotted. Western blot or qRT-PCR was used to analyze the effect of NEK2 on the phosphorylation levels of ERK and c-JUN in two GC cell lines (BGC823 and SGC7901) with NEK2 overexpression, and the expression of the downstream effector cyclin D1.Furthermore, CCK8, EdU incorporation assay, and flow cytometry were used to detect the proliferative ability of BGC823 and SGC7901 cells with stably silenced ERK.RESULTS NEK2 was significantly up-regulated in human GC tissues. ERK was significantly associated with NEK2 expression in human clinical specimens, and combined overexpression of NEK2 and ERK potentially forecasted a poor prognosis and Received: April 4, 2019 Peer-review started: April 4, 2019 First decision: April 11, 2019 Revised: April 27, 2019 Accepted: May 18, 2019 Article in press: May 18, 2019 Published online: June 21, 2019 P-Reviewer: Jones G, Tanabe S S-Editor: Ma RY L-Editor: Wang TQ E-Editor: Ma YJ survival in GC patients. NEK2 knockdown in GC cells inhibited ERK and c-JUN phosphory-lation and reduced the transcription of cyclin D1. More interestingly,NEK2 can rescue the inhibition of cellular viability, proliferation, and cell cycle progression due to ERK knockdown.CONCLUSION Our results indicate that NEK2 plays a carcinogenic role in the malignant proliferation of GC cells via the ERK/MAPK signaling, which may be important for treatment and improving patient survival.
Key words: NIMA related kinase 2; ERK/MAPK signaling; Cyclin D1; Cell proliferation;Gastric cancer
INTRODUCTION
Gastric cancer (GC) is a type of malignant tumor in the digestive system with high incidence, and it is the third cause of human cancer-related death[1-3]. Although surgery, radiation therapy, and chemotherapy can improve patient survival to some extent, the 5-year survival rate of GC patients is still low (25%-30%), partly because the complexity of clinical heterogeneity[4-6]. Due to high cell proliferation is associated with poor survival in patients with GC[7], exploring the mechanisms of malignant proliferation of GC cells is important for the treatment and improvement of patient survival[8].
In mammals, NIMA related kinase 2 (NEK2), as an S/G2 phase kinase, usually shows a high expression level in various forms of cancer, mediating the cellular mitosis during cell division[9-11]. In vivo, silencing NEK2 can significantly slow tumor growth in mice and result in reduced tumor burden in tumor-forming organs[12-14].Furthermore, some studies have shown that up-regulated NEK2 leads to malignant proliferation and drug resistance in vivo, and NEK2 siRNA can significantly reduce tumor formation ability in allogeneic tumor formation studies[15,16]. Although the exact molecular mechanism of NEK2 in the regulation of tumor formation and progression is largely unclear, the fact that NEK2 silencing induces the slower proliferation of cells suggests that NEK2 inhibition can resist tumor progression in vivo, and inhibition of NEK2 expression may become an effective treatment for GC[14,17-20].
The ERK/MAPK pathway communicates intracellular phosphorylation kinase signaling from a receptor on the surface of the cell to the DNA in the nucleus, and produces some changes in cell processes to respond to the extracellular stress[21,22].Overall, once the extracellular mitogen binds to the receptor on the membrane, ERK will be activated and its downstream effector c-Jun will also be switched by transferring a phosphate group[23], participating in the ERK/MAPK signal transduction[24,25]. Many studies show that activation of the ERK/MAPK pathway plays an important role in tumorigenesis, progression, survival in GC[26-28].
Here, we showed that NEK2 promotes gastric tumorigenesis by regulating the cell cycle. Based on the rise of NEK2 expression in GC and the application of bioinformatics and clinical sample analysis, our study focused on the combined expression of NEK2 and ERK in GC and demonstrated that up-regulation of NEK2 in GC cells by targeting ERK/c-JUN/cyclin D1 signaling effectively promoted the proliferation of GC cells and significantly affected the clinical prognosis of GC patients. Our study revealed the combined role of the NEK2 and ERK/MAPK pathways in the cell cycle, which is important for the GC treatment and improvement of patient survival.
MATERIALS AND METHODS
Human tissue specimens and cell lines
GC and paired adjacent tissues were obtained from 30 patients treated at Zhang jiagang First People's Hospital from 2016 to 2018. All patients had no history of chemotherapy or immunotherapy, and all samples were classified as GC by two pathologists. The sample use was approved by the patients and the Hospital Ethics Committee, and all patients provided informed consent.
The AGS, MGC803, BGC823, and SGC7901 cell lines and human normal gastric epithelial cell line (GES1) used in the study were purchased from the Chinese Academy of Medical Sciences Cell Resource Platform (Beijing, China). All cell lines were cultured in DMEM (Gibco) containing 10% FBS (Gibco) and 1% antibiotic(Gibco).
Cell transfection and lentivirus transductions
We transfected siRNA oligos into cells using Lipofectamine 3000 (Invitrogen). SiRNAs against NEK2 (siNEK2-1, 5'- GUCAGAUAUUGAGAAAAAUUACCAA-3'; siNEK2-2,5'- AAUGAGAAAUCAGAUAUCUGGUCAT-3') and control siRNA [siN.C, 5'-UUCUCCGAACGUGUCACGUd(TT)-3'] were obtained from GenePharma (Suzhou).After 48 h of transfection, cells were harvested for subsequent experiments.
The shRNA targeting the ERK gene (5'- AACCTTTAAAGACTGATATTCAAAT-3')was designed and cloned into the pGV307-RFP vector (Addgene). Lentiviral packaging, purification, and infection of the cells of interest were performed according to the protocol provided by the manufacturer. Cells with stable ERK knockdown were selected using 5 μg/mL puromycin (Sigma) at 3 d after infection.
RNA extraction and quantitative real-time PCR (qRT-PCR)
RNA was extracted from cells or tissue samples using TRIzol Reagent (Sigma), and 2 μg of RNA was transcribed into cDNA with a Prim primer RT Master Mix Kit(Takara) using random primers, followed by qRT-PCR using GoTaq qRT-PCR Master Mix (Promega, cat). The ΔΔCT method was used to calculate the expression level of the target mRNA. Experiments were performed in triplicate. The primers used in our study are listed in Table 1.
Immunohistochemistry
Paraffin-embedded tissue sections were deparaffinized and dehydrated, and immunohistochemical staining for NEK2 and ERK in tissues was performed using a DAB substrate kit (Maxin). The staining intensity and staining range were respectively scored using the standard method, and according to the final score(staining intensity × staining range), a score of less than 6 was considered to be low staining, and > 6 high staining.
Western blot analysis
Total cellular protein was extracted using RIPA lysis containing protease and phosphatase inhibitors. Protein concentration was measured according to the instructions of the BCA kit (Thermo Scientific). SDS-PAGE separation and immunoblotting were then performed according to standard methods. The primary antibodies used in our study are listed below: EK2 (Abcam), ERK1/2 (Cell Signaling Technology), c-JUN (Cell Signaling Technology), phospho-Erk1/2 (Thr202/Tyr204)(Cell Signaling Technology), phosphorylated c-JUN (Ser63) (Cell Signaling Technology), cyclin D1 (Abcam), and β-actin (Abcam).
CCK-8 assay
Cells with stable ERK knockdown or NEK2 overexpression were seeded in 96-well plates at a density of 1000 cells per well. After incubation for 24, 48, and 72 h, CCK-8 reagent (Dojindo) was added at a concentration of 10% to each well, and after incubation for 1 h, the optical density (OD) was measured at 490 nm. Experiments were performed in triplicate.
EdU incorporation assay
EdU incorporation assay was performed with an EdU Apollo 488 in vitro imaging kit(RiboBio). One thousand cells were seeded into each well of a 96-well plate. After incubation for 24 h, the medium was changed to a medium containing 20 mmol/L EdU, and incubation was continued at 37 °C for 1 h. After fixing the cells with 4%paraformaldehyde, Apollo staining was performed according to the standard instructions. Cell proliferation rates are shown as the percentage of EdU positive cells.Experiments were performed in triplicate.

Table 1 Primers used for quantitative real-time PCR
Flow cytometry
When the cells in the 6-well plate reached 80% confluence, they were harvested using trypsin and fixed overnight in pre-cooled 70% ethanol. After washing, cells were incubated in a dark environment with 500 μL of PI/RNase staining buffer (BD Pharmingen) for 15 min. Cell cycle changes were detected and analyzed using the FACSCalibur system (BD Biosciences). Experiments were performed in triplicate.
Statistical analysis
Data were analyzed using GraphPad Prism 5 (La Jolla, CA, United States). The chisquare test or Fisher's exact test was used to determine the significance of categorical data. The Student's t-test or one-way ANOVA was used to determine the significance of the mean. Differences were considered statistically significant at P < 0.05 (aP < 0.05,bP < 0.01,cP < 0.001).
RESULTS
Expression of NEK2 is significantly up-regulated in GC
The amplification of NEK2 has an oncogene effect in various types of tumors,regulating the process of tumor proliferation and invasion. To clarify the expression pattern of NEK2 in human GC, we searched the "Oncomine" database for GC data published by different researchers. It was surprised to find that NEK2 expression in GC tissues was significantly higher compared to that in normal tissues (Figure 1A). To confirm the high level of NEK2 expression in GC, we detected the NEK2 expression level in 30 pairs of GC and adjacent tissues. The results showed that the NEK2 mRNA level in GC tissues was significantly higher than that in non-cancerous tissues (Figure 1B).
In addition, in order to understand whether the high expression of NEK2 has an effect on the progression of GC, we divided 30 human GC samples into three grades(I, II, and III-IV) according to the WHO criteria for clinical GC tumors to evaluate the expression pattern of NEK2 in various stages of GC. Interestingly, high-grade tumors(Grade III-IV) showed a higher NEK2 expression level than low-grade tumors (Figure 1C), giving a suggestion that NEK2 expression is gradually increasing in the progression of GC. These results indicated that the progressively up-regulated expression of NEK2 may be a tumor-promoting regulator in the progression of GC.
To further verify that NEK2 is up-regulated in GC, the mRNA and protein expression levels of NEK2 were detected in four gastric tumor cell lines (AGS,MGC803, BGC823, and SGC7901) and gastric epithelial cell line (GES1). In the four GC cell lines, both the mRNA and protein expression levels of NEK2 were higher than those in the normal gastric epithelial cell line (Figure 1D and E), which is consistent with the expression pattern of NEK2 in the tissues. Taken together, these data can confirm that NEK2 expression shows a dynamic up-regulation pattern in GC tissues,and NEK2 plays a potential tumor-promoting role in the pathological process of GC.
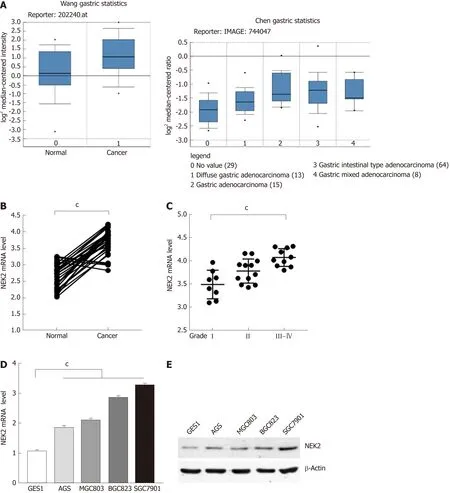
Figure 1 Expression of NlMA related kinase 2 in tissues and cells of gastric cancer. A: Relative mRNA expression level of NIMA related kinase 2 (NEK2) in gastric cancer and normal tissues (datasets from Oncomine). B: Relative mRNA expression level of NEK2 analyzed by quantitative real-time PCR in 30 pairs of human gastric cancer tissues (Cancer) and adjacent tissues (Normal). (Paired t-test, P < 0.001). C: The mRNA expression level of NEK2 in different grades of cancer samples. Data represents the mean ± SD of three replicates. D: Relative NEK2 mRNA expression level in four gastric cancer cell lines compared to gastric mucosal epithelial cell line (GES1). E: Relative NEK2 protein expression level in different gastric cancer cell lines. β-actin was used as the loading control. NEK2: NIMA related kinase 2.
Combined NEK2 and ERK expression may predict overall survival in patients with GC
Research has shown that there is a correlation between the activation of ERK1/2 and NEK2 during mitosis[29]. In order to further elucidate the molecular mechanism of the correlation of ERK1/2 and NEK2, we tried to detect the correlation of ERK1/2 and NEK2 expression in GC. First, we found that NEK2 and ERK had strong co-expression relationships in GC based on the TCGA database (Figure 2A). We next examined the ERK expression level in GC tissues, and the result showed that ERK was significantly up-regulated in GC tissue relative to normal tissue (Figure 2B). In addition, in order to confirm the expression relationship between NEK2 and ERK in GC, we further examined the expression relationship between NEK2 and ERK in clinical samples. The RT-qPCR result showed that NEK2 was positively correlated with ERK in GC tissues in terms of the expression level (Spearman, r = 0.594, P < 0.05) (Figure 2C).Collectively, all results suggest a correlation in expression patterns between NEK2 and ERK in GC.
Although the abnormal NEK2 expression is related to the therapeutic effect and prognosis in many cancers, the prognostic significance of NEK2 in GC patients remains unclear. We first classified 30 GC tissues into different groups according the expression of NEK2 and ERK (Figure 2D). The results showed that 56.67% (17/30) of the 30 GC samples showed strong NEK2 immunostaining, while in the high NEK2 expression samples, only 23.53% (4/17) of the tumors showed low ERK expression. In the samples with low expression of NEK2 (43.33%, 13/30), only five samples exhibited high REK expression. Whatever, the χ2 test revealed a significant expression relation between ERK and NEK2 (P = 0.035, Figure 2E). Analysis of the expression of NEK2 and ERK using the Kaplan-Meier method in GC patients indicated that patients with low NEK2 and low ERK expression levels had better overall survival than patients with high NEK2 and high ERK levels (Figure 3D). The above results suggest that co-expression of NEK2 and ERK in tissues may be involved in the pathological process of GC and significantly affect the prognosis of patients with GC.
NEK2 promotes the activation of ERK/MAPK signaling in GC
To elucidate the underlying molecular mechanisms by which NEK2 affects the pathogenesis of GC and the prognosis of patients with GC, we analyzed the effect of NEK2 on MEK/ERK signaling. Cell proliferation-related gene expression patterns were examined in BGC823 and SGC7901 cells after NEK2 silencing. We found that,when NEK2 expression in BGC823 and SGC7901 cells was significantly reduced(Figure 4B), the expression of key genes regulating cell proliferation was significantly down-regulated, including c-JUN and cyclin D1, which are considered to be the downstream molecule of the ERK/MAPK signaling pathway (Figure 4A).
In addition, NEK2 silencing resulted in decreased levels of c-JUN and ERK phosphorylation, as revealed by Western blot analysis, although c-JUN and ERK did not change significantly at the protein level. And grayscale comparison showed their phosphorylation levels were significantly reduced (Figure 4B and C). In addition,cyclin D1 protein expression was significantly reduced after down-regulation of NEK2 expression in BGC823 and SGC7901 cells (Figure 4B). These results revealed that the expression of cyclin D1 can be regulated by NEK2 through the ERK/c-JUN pathway in GC.
NEK2 promotes cell viability, proliferation, and cell cycle progression in GC via ERK/MAPK signaling
To evaluate whether NEK2 regulates potential phenotypic changes in GC cells via the ERK/c-JUN/cyclin D1 pathway, we used shRNA to establish BGC823 and SGC7901 cells with stable ERK knockdown. We found that cyclin D1 was significantly reduced both at the mRNA and protein levels in the NEK2 knockdown cell line compared to control cells (Figure 3A and B), while overexpression of NEK2 rescued the decreased cyclin D1 expression caused by ERK knockdown.
To explore how NEK2 mediates cell viability in GC, the CCK-8 assay was used to detect the viability of GC cells in BGC823 and SGC7901 cell lines with ERK knockdown. CCK8 assay showed that down-regulation of ERK significantly reduced the proliferation of BGC823 and SGC7901 cells after cell seeding, but cell viability was restored after stably expressing NEK2 in ERK-silenced BGC823 and SGC7901 cells(Figure 3C and D).
We next investigated the effect of silencing NEK2 on cell cycle progression by flow cytometry. Cell cycle distribution analysis showed that ERK silencing resulted in cell cycle arrest in the G0/G1 phase (Figure 4E and F), and the EdU incorporation assay showed that ERK silencing resulted in a decrease in the percentage of S phase cells,whereas both the cell cycle and DNA synthesis in BGC823 and SGC7901 cells were restored to some extent after overexpression of NEK2 (Figure 4G and H). Altogether,these data indicate that NEK2 can regulate GC cell cycle progression and cell proliferation via the ERK/c-JUN/cyclin D1 axis.
DISCUSSION
Previous studies have indicated that, unlike other kinases, NEK2 can promote cancer progression by participating in the regulation of centrosome activity during mitosis,leading to abnormal activation of mitosis. Therefore, the regulation of NEK2 is mainly involved in cancer during the advanced stage[30-32]. In the development of cancer at early stages, for example, NEK2 promotes tumor cell proliferation and participates in the invasion of tumor cells to epithelial tissues[9,16]. It is speculated that enhanced activity of centrosomes is a feature of advanced cell carcinogenesis, driving mutations to progeny cells and providing a source of genetic variation in tumorigenesis[33].

Figure 2 Combined NlMA related kinase 2 and ERK expression may predict overall survival in patients with gastric cancer. A: ERK co-expressed with NIMA related kinase 2 (NEK2) in gastric cancer (datasets from TCGA). B: Relative mRNA expression level of ERK analyzed by quantitative real-time PCR in 30 pairs of human gastric cancer tissues. C: ERK expression is significantly correlated to NEK2 expression in human gastric cancer tissues. D: Immunohistochemistry analysis of NEK2 and ERK expression in serial gastric cancer samples. E: Correlation between NEK2 and ERK expression. F: Overall survival of patients with gastric cancer calculated using Kaplan-Meier analysis according to the staining level. NEK2: NIMA related kinase 2.
In this study, we for the first time showed that NEK2 overexpression promoted the activation of c-JUN and the up-regulation of cyclin D1 through the abnormally activated ERK/MAPK pathway in GC. We also found that NEK2 overexpression promoted the cell cycle progression in GC cell lines and participated in the regulation of proliferation of GC cells. In human myeloma, NEK2 promotes myeloma resistance via activating the ERK pathway, indirectly supporting our conclusion[34-36]. Thus, our results suggest that NEK2 may have a new and important role in tumor progression.
In many mammalian cell types, the ERK pathway can integrate cellular external stimulation signals into cells, acting as a significant regulator that mediates cell growth and proliferation[37,38]. It is well known that uncontrolled growth or malignant proliferation is a necessary and initial step in the development of tumors[39,40].Furthermore, it has been shown that abnormal NEK2 expression obviously affected the process of mitosis of breast tumor cells, and in breast cancer, NEK2 overexpression was significantly associated with a poor prognosis, indicating that NEK2 may be an ideal target for therapy[9,41]. However, in our study, we demonstrated that NEK2 was highly expressed in GC and the up-regulated expression of NEK2 was associated with the activity of the ERK/MAPK pathway. NEK2 can promote phosphorylation of ERK and further promote cyclin D1 expression through the ERK/c-JUN pathway, regulating the mitosis of GC cells. In addition, the combined and elevated expression of NEK2 and ERK was found to be significantly associated with a poor prognosis in patients with GC, indicating that NEK2 is an essential factor that mediates the transmission of important signals in GC. Therefore, NEK2 has the potential to become an ideal target for GC therapy[41,42].

Figure 3 NlMA related kinase 2 mediates the activation of ERK/MAPK signaling. A and B: Relative mRNA expression of ERK/MAPK signaling key factors analyzed by quantitative real-time PCR in NIMA related kinase 2 (NEK2) silenced BGC823 and SGC7901 cell lines and negative controls. C: Western blot analysis of ERK/MAPK signaling key factors (ERK, c-JUN, and cyclin D1) and their phosphorylated forms (p-ERK and p-c-JUN) in NEK2 silenced BGC823 and SGC7901 cell lines and negative controls. D: Relative phosphorylation levels of ERK and c-JUN in C were calculated. β-actin was used as the internal control. Values are presented as the mean ± SEM. NEK2: NIMA related kinase 2.
In conclusion, our study reveals a novel mechanism by which NEK2 promotes the growth of GC cells by activating the ERK/MAPK pathway. Although we have demonstrated that NEK2 can regulate the phosphorylation of c-JUN and ERK and cell cycle progression in GC cells, there may be other targets that may also affect cancer cell proliferation. Nonetheless, our finding that NEK2 targets the ERK/c-JUN/cyclin D1 pathway provides new insights into NEK2 function and reveals that NEK2 may be a potential target for developing optimal treatments for GC patients.
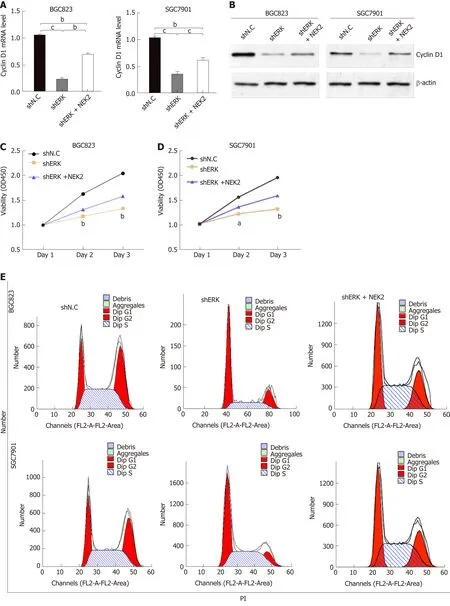
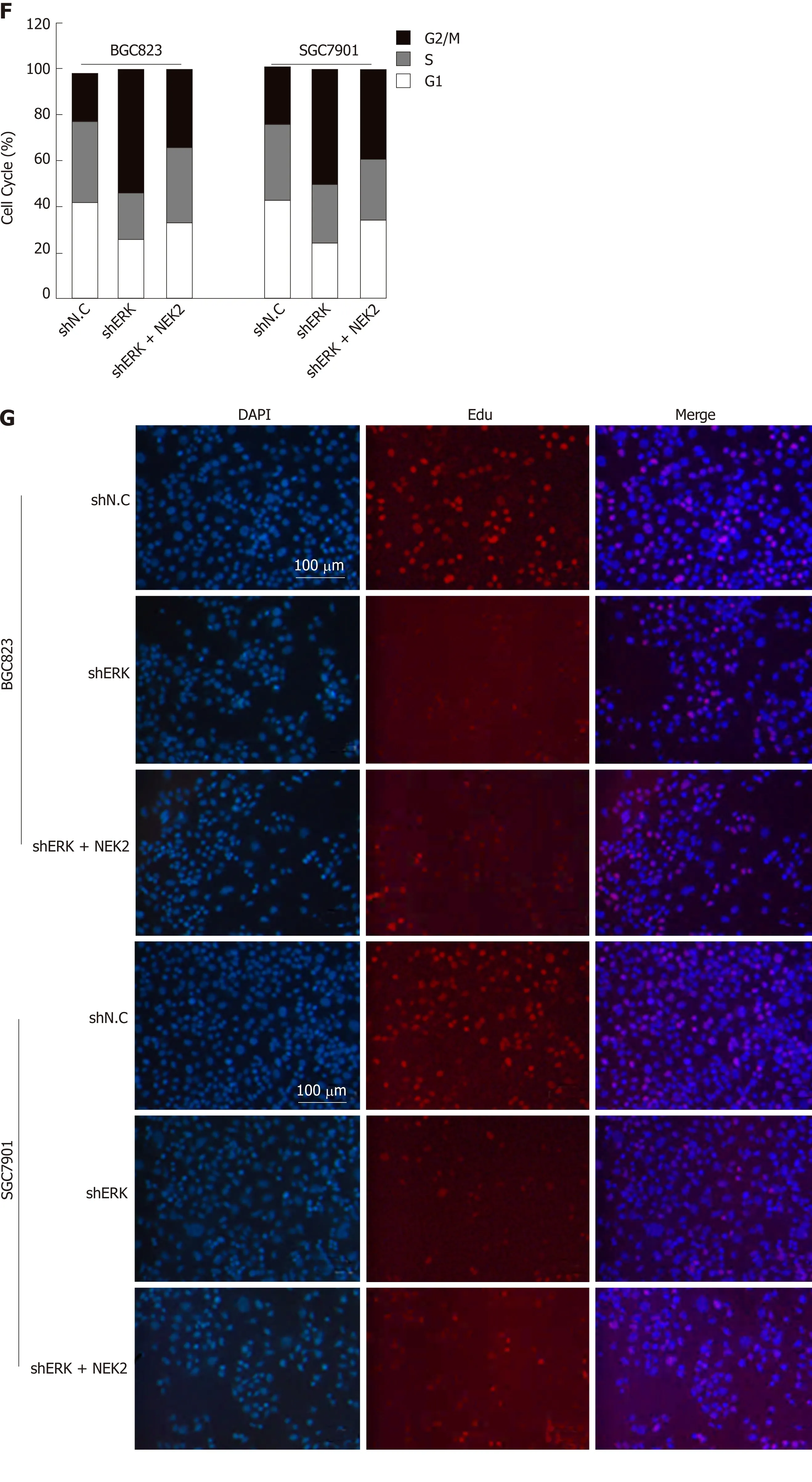
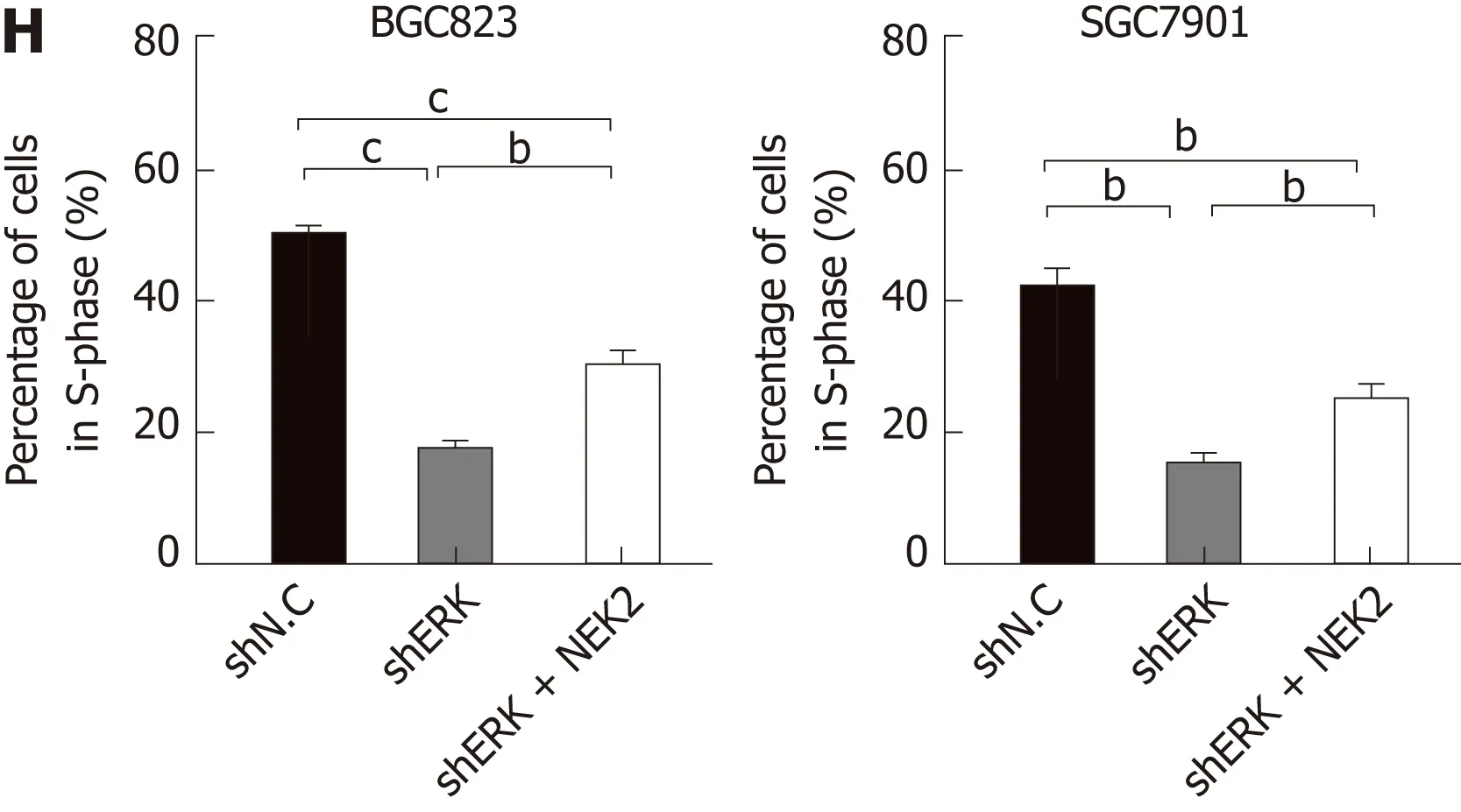
Figure 4 NlMA related kinase 2 promotes gastric cancer cell viability, proliferation, and cell cycle progression via ERK/MAPK signaling. A and B: Relative mRNA and protein expression of cyclin D1 in cell lines with ERK silencing and NEK2 overexpression. C and D: CCK8 assay showing cell viability in BGC823 and SGC7901 cell lines and negative controls. E: Flow cytometry analysis of cell cycle distribution in BGC823 and SGC7901 cell lines. F: Quantification of the data shown in E (n = 3). G: EdU incorporation assay showing the percentage of cells in S-phase. H: Quantification of the data shown in G (n = 3). NEK2: NIMA related kinase 2.
ARTICLE HIGHLIGHTS
Research background
NIMA related kinase 2 (NEK2) is closely related to mitosis, and it is currently considered to be overexpressed frequently in many poorly prognostic cancers. However, the effect of upregulated NEK2 expression on cellular signaling in tumors, such as gastric cancer (GC), is confusing.
Research motivation
To explore the treatment of GC and improve patient survival.
Research objectives
To determine the role of the up-regulation of NEK2 in GC.
Research methods
To investigate the pathological significance of NEK2 in GC, the expression of NEK2 in GC was investigated based on the “Oncomain” database and compared between 30 pairs of cancer and adjacent tissues. The co-expression of NEK2 and ERK in GC was analyzed based on the TCGA database and confirmed in clinical samples by quantitative real-time PCR (qRT-PCR), and the survival curve was also plotted. Western blot or qRT-PCR was used to analyze the effect of NEK2 on the phosphorylation levels of ERK and c-JUN in BGC823 and SGC7901 cells, and the expression of the downstream effector cyclin D1. Furthermore, CCK8, EdU incorporation assay,and flow cytometry were used to detect the proliferative of BGC823 and SGC7901 cell lines with stable silencing of ERK.
Research results
In this study, we found that NEK2 was significantly up-regulated in human GC tissues. In addition, ERK was significantly associated with NEK2 expression in human clinical specimens,and combined overexpression of NEK2 and ERK potentially forecasted a poor prognosis and survival in GC patients. NEK2 knockdown in GC cells inhibited ERK and c-JUN phosphorylation and reduced transcription of cyclin D1. More interestingly, NEK2 can rescue the inhibition of cellular viability, proliferation, and cell cycle progression due to ERK knockdown.
Research conclusions
NEK2 plays a carcinogenic role in the malignant proliferation of GC cells via the ERK/MAPK signaling, which is important for treating GC and improving patient survival.
Research perspectives
Future research may further reveal the mechanism of action of NEK2 to enhance the sensitivity of cancer cells and promote its application in anti-cancer treatments. And the identification of the molecular pathway related to ERK/MAPK signaling may further elucidate the underlying mechanism.
 World Journal of Gastroenterology2019年23期
World Journal of Gastroenterology2019年23期
- World Journal of Gastroenterology的其它文章
- Modified FOLFlRlNOX for resected pancreatic cancer: Opportunities and challenges
- Role of cytochrome P450 polymorphisms and functions in development of ulcerative colitis
- Role of epigenetics in transformation of inflammation into colorectal cancer
- Postoperative complications in gastrointestinal surgery: A “hidden”basic quality indicator
- The role of endoscopy in the management of hereditary diffuse gastric cancer syndrome
- Predicting systemic spread in early colorectal cancer: Can we do better?