
半干旱黄土区苜蓿退化对坡面草本植物分布及多样性的影响
2019-07-05 09:01王子婷柴春山张洋东张继强
生态学报 2019年10期
王子婷,杨 磊,李 广,柴春山,张洋东,陈 荣,张继强
1 甘肃农业大学林学院,兰州 730070 2 甘肃省林业科学研究院,兰州 730020 3 中国科学院生态环境研究中心城市与区域生态国家重点实验室,北京 100085 4 定西市水土保持科学研究所,定西 743000
人工植被恢复是黄土高原控制水土流失的关键措施之一[1- 4]。半干旱黄土区沟壑纵横、土壤侵蚀严重,严重制约了该区社会经济发展和区域环境改善,我国自20世纪80年代以来陆续开展了以小流域为单元的水土流失综合治理工程。尤其是1999年开始实施了大面积退耕还林还草工程,改变了这一地区植被覆盖状况,有效减少了土壤侵蚀,改善了区域生态环境[5-10]。然而,我们对该区域人工植被恢复演替过程的认识还存在一定的不足,尤其是对地形引起的土壤水分再分配对人工植被的影响及群落特征响应还缺乏系统的探讨。
紫花苜蓿(MedicagosativaL.)作为一种优良的牧草,具有较好的抗旱固土能力,在黄土高原被广泛种植[11- 12]。然而,紫花苜蓿根系较深,随着苜蓿生长其对水分的消耗亦在增加。罗珠珠等[13]在陇中黄土区的研究发现,人工种植苜蓿1年以后0—300 cm土壤水分出现不同程度的干旱且随种植年限的增加逐渐加剧。万素梅等[14]研究发现,黄土高原苜蓿随生长年限的增加土壤干层不断加深,苜蓿生长低于8年在250—350 cm土层出现轻度干层,超过8年干层范围延深至500 cm土层以下,超过18年则深层土壤通体干化且水分难以恢复。由此可见,在黄土区种植苜蓿虽然能够减少水土流失、增加土壤肥力,但其对土壤水分的过度消耗会引起不同深度土壤干化从而引起苜蓿草地退化。此外,有研究还发现苜蓿草地退化也在一定程度上促进了表层土壤水分恢复,这为其他草本植物生长提供了适宜条件,促进了人工苜蓿草地群落向天然草地群落的转变[15- 17]。土壤水分变化、苜蓿退化等驱动了人工苜蓿草地草本群落的恢复和演替过程,而这一过程又受坡面地形条件的影响。鉴于此,本研究以坡面退化苜蓿草地为研究对象,探索坡向及坡位变化对紫花苜蓿草地群落组成的影响,确定地形变化、土壤水分及苜蓿退化程度对坡面草地群落组成及分布的耦合影响和互馈作用特征,为半干旱黄土区坡面退化人工草地的恢复及可持续管理提供科学依据。
1 研究区概况
研究区位于甘肃省定西市龙滩小流域(104°27′—104°32′E,35°43′—35°46′N),研究区属典型半干旱黄土丘陵区,流域面积16 km2,平均海拔1900 m。研究区年平均气温6.8℃,1月份平均气温-7.9℃,极端最高温38.5℃,平均无霜期152 d,平均日照时数2052 h。多年平均降水量为386 mm,降雨主要集中在7—9月份,潜在蒸发量1439 mm。年平均相对湿度72%,干燥度1.9。流域内土壤以黄绵土为主,有机质含量低,土壤贫瘠且易侵蚀。天然植被以多年生草本为主,主要为长芒草(StipabungeanaTrin.)、赖草(Leymussecalinus(Georgi) Tzvel.)、阿尔泰狗娃花(Heteropappusaltaicus(Willd.) Novopokr.)等。因退耕还林(草)政策的实施,流域在2003年以来大面积人工种植的牧草为紫花苜蓿(MedicagosativaL.)。
2 研究方法
2.1 监测样点布设
在研究区内一个全部种植紫花苜蓿的典型子流域选择东向、西向和北向3个坡面,分别在每个坡面自上而下依次选取上、中、下3个坡位布设监测样点(表1),开展生长季土壤水分的定位监测,并开展苜蓿植被群落调查。研究区苜蓿均为2003年种植,种植年限14年,苜蓿在坡面上自上而下沿等高线呈条带状种植,条带宽3—5 m。研究区内苜蓿在种植初期存在较好的人工管理,根据降水状况每年收割1—2次,但近年来随着流域内人口逐渐迁出,苜蓿草地也疏于管理或无人管理,均出现不同程度的退化。
W1:西上坡位,West upper slope position;W2:西中坡位,West middle slope position;W3:西下坡位,West foot slope position;N1:北上坡位,North upper slope position;N2:北中坡位,North middle slope position;N3:北下坡位,North foot slope position;E1:东上坡位,East upper slope position;E2:东中坡位,East middle slope position;E3:东下坡位,East foot slope position
2.2 数据采集
2017年7月初—8月底期间在各监测样点上开展草本群落组成调查并测定草本地上生物量。每个监测样点选取3个1 m × 1 m的样方,统计样方内草本物种数、高度、多度、盖度、分布、生活力及物候,随后将紫花苜蓿和其他草本地上部分进行刈割。样品带回实验室称重后在105℃下杀青1 h后再在80℃下烘干72 h,分别获得紫花苜蓿和其他草本地上生物量。
利用GPS记录各监测样点的经纬度及海拔信息,利用坡度仪和罗盘分别测定坡度和坡向。在每个监测样地预先布设聚碳酸酯探管,利用Trime-FM土壤水分速测仪测定土壤水分含量。于2017年生长季(5月初—10月中旬)对各监测样点土壤水分含量进行测定,每15 d测定一次,土壤剖面0—40 cm每10 cm为一测层,40—200 cm每20 cm为一测层,每次每层重复测定并取平均值作为该次该层土壤含水量(%),生长季内剖面土壤水分变化见图1。根据该区草本及紫花苜蓿根系主要分布范围,本文分别统计植物生长季内0—20 cm、20—80 cm和80—200 cm 3个层次土壤含水量,分别命名为SW1、SW2和SW3用以分析不同土层深度土壤含水量与草本分布及多样性之间的关系。

图1 监测样点生长季0—200 cm剖面平均土壤水分含量Fig.1 Vertical distributions of temporal-averaged soil moisture content on different sample plots
2.3 数据统计
统计各监测样点内草本物种数,使用每种草本的相对高度和相对盖度计算其重要值(IV)[18]。利用草本重要值计算各监测样点内草本群落Simpson优势度指数、Shannon-Wiener多样性指数和Pielou均匀度指数,利用Sorenson相似性指数比较各监测样点内草本群落组成的相似性。计算方法见下述有关公式。
重要值(IV):IV= RHI+ RCO
Simpson优势度指数:D= 1-∑pi2
Shannon-Wiener多样性指数:H′= -∑pilnpi
Pielou均匀度指数:J=(-∑pilnpi)/lnS
Sorenson相似性指数:C=2m/(a+b)
式中,RHI为相对高度;RCO为相对盖度;pi为第i个物种的相对重要值;S为种i所在样方的物种总数;a和b为群落A和B的物种数;m为群落A和B共有的物种数。
利用方差分析检验坡向、坡位及二者的交互对草本地上总生物量、苜蓿生物量和其他草本生物量分布的影响是否显著。利用Pearson相关分析苜蓿地上生物量、其他草本地上生物量、草本地上总生物量、物种丰富度、多样性、均匀度和优势度指数及4个优势草本种(赖草、长芒草、茵陈蒿和阿尔泰狗娃花)与不同深度土壤水分含量(0—20 cm、20—80 cm和80—200 cm)及苜蓿地上生物量间的相关性。利用典范对应分析(CCA)确定地形(坡向和坡位)、苜蓿生长状况(地上生物量)和土壤水分与坡面草本群落分布之间的关系,采用蒙特卡洛置换检验(Monte-Carlo permutation test 999)第一排序轴和所有排序轴的显著性,然后通过CCA排序中逐步筛选(Interactive-forward-selection)过程和蒙特卡洛置换检验(Monte-Carlo permutation test 999)确定环境因子的相对贡献率。CCA分析使用Canoco for Windows 4.5软件包。地形、土壤水分和苜蓿的生物量等6个因子和4个优势草本种与排序轴1和轴2的关系通过Pearson相关分析确定。统计分析使用SPSS 21.0软件包。
3 结果
3.1 不同地形条件下草本群落物种组成
3条样带不同坡位苜蓿草地内共有草本26种,分属10科20属,其中菊科(8种)、禾本科(4种)、豆科(4种)和蔷薇科(4种)是主要的草本类群(表2)。西坡样带自上而下苜蓿重要值依次增加,草本物种数依次减少。西坡菊科茵陈蒿重要值在不同坡位间相差不大,阿尔泰狗娃花分布在中、上坡位且重要值相近;禾本科赖草和长芒草重要值上坡位高于中、下坡位;蔷薇科星毛委陵菜仅分布于上坡位。北坡样带不同坡位间苜蓿重要值变化不大,草本物种数相差不大。北坡菊科阿尔泰狗娃花重要值中、下坡位高于上坡位,茵陈蒿重要值上坡位高于中、下坡位;禾本科长芒草分布在上、下坡位且重要值上坡位大于下坡位,赖草分布在中、下坡位且重要值相近;蔷薇科二裂委陵菜仅分布在上坡位,伏毛山莓草分布在中、下坡位且重要值相近。东坡样带自上而下苜蓿重要值依次增加,草本物种数依次降低。东坡禾本科长芒草重要值在上坡位高于中、下坡位,禾本科赖草、蔷薇科二裂委陵菜、菊科阿尔泰狗娃花重要值在不同坡位间均相差不大。
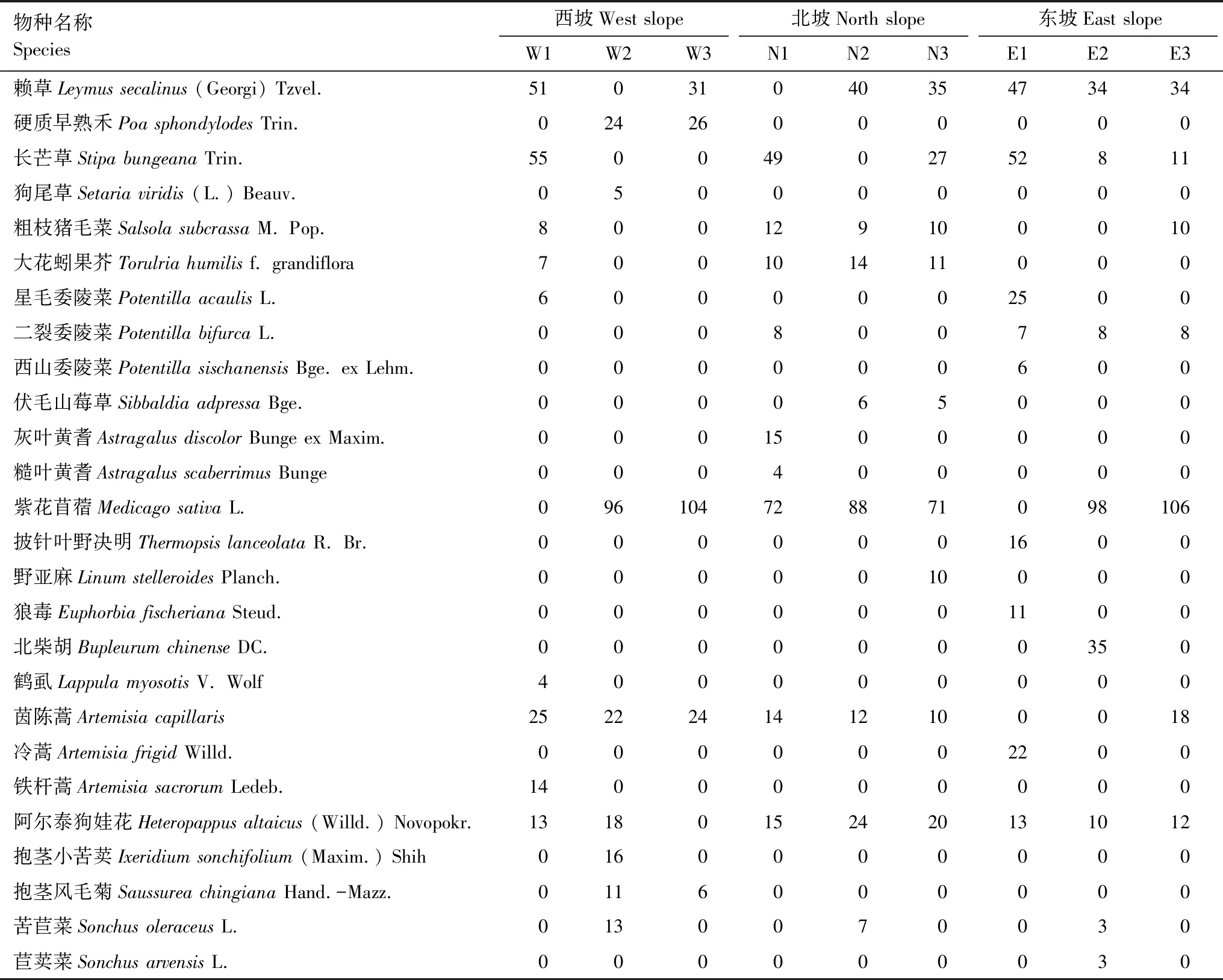
表2 不同坡位苜蓿草地草本的重要值
3.2 不同地形条件地上生物量比较
不同地形条件草本群落地上生物量的方差分析结果表明,坡向对苜蓿草地地上总生物量影响较小(P=0.524),坡位及坡向和坡位的交互均对草本地上总生物量有显著影响(P<0.001,P=0.006)。西坡和东坡样带,中、下坡位草本群落地上总生物量显著高于上坡位,其中西坡样带表现尤为明显(图2)。苜蓿地上生物量在不同坡位间的变化趋势与地上草本群落总生物量变化相近,坡向、坡位及二者的交互均对苜蓿地上生物量有显著影响(P<0.001,P<0.001,P<0.001)。然而,其他草本的地上生物量在不同坡位间的变化趋势与苜蓿地上生物量变化相反,即西坡和东坡样带上、中坡位其他草本地上生物量显著高于下坡位(图2)。

图2 不同坡位退化苜蓿草地苜蓿、其他草本和总草本地上生物量比较Fig.2 Comparison of above-ground biomass between alfalfa, other herbs, and total biomass in experimental sites
3.3 不同地形条件其他草本群落结构及相似性
3条样带内物种丰富度及多样性指数表现为西坡和东坡高于北坡,均匀度指数则表现为西坡和东坡低于北坡,优势度指数在不同坡向间变化较小(表3)。其中西坡不同坡位间,其他草本的物种丰富度自下而上逐渐升高,多样性和优势度指数上、中坡位高于下坡位,均匀度指数上坡位低于中、下坡位;北坡不同坡位其他草本多样性指数均相差较小;东坡样带其他草本的物种丰富度自下而上也逐渐升高,多样性和优势度指数上坡位略高于中、下坡位,均匀度指数不同坡位间相差较小(表3)。
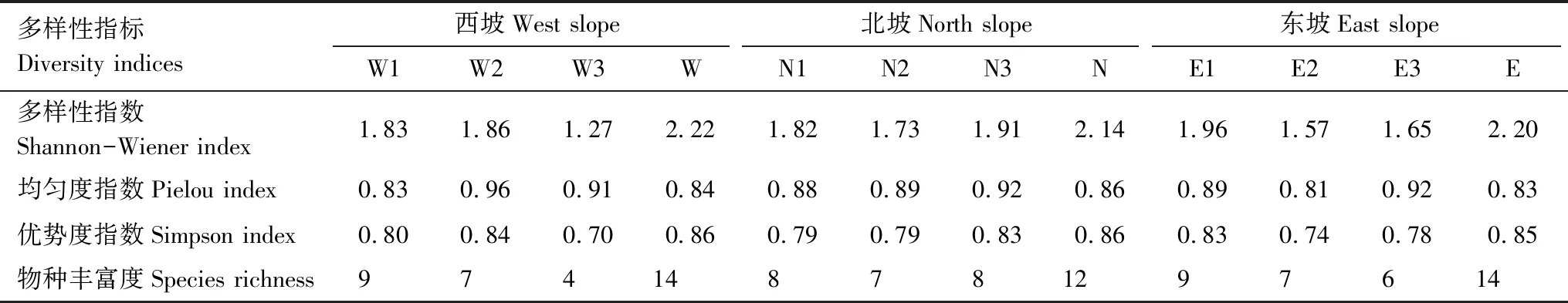
表3 不同地形条件草本多样性比较
W:西坡,West slope;N:北坡,North slope;E:东坡,East slope
不同坡向间其他草本物种相似度指数表现为,西坡与北坡相似度指数0.54、西坡与东坡0.57、北坡和东坡0.54。不同坡位间其他草本物种相似度指数表现为,上坡位与中坡位相似度指数0.48,上坡位与下坡位0.55,中坡位和下坡位0.79。
3.4 苜蓿退化、土壤水分及地形变化对草本群落分布的影响
相关分析表明,苜蓿地上生物量与其他草本地上生物量呈显著负相关(r=-0.36,P=0.062),与其他草本的物种丰富度、多样性和优势度指数呈负相关关系,而与其他草本的均匀度指数呈正相关关系;不同深度土壤水分与草本群落结构的相关性均较小(表4)。
表4 草本群落结构及4个优势种与不同深度土壤水分及苜蓿生物量的Pearson相关系数
Table 4 Pearson correlation coefficient between herbage community structure & four dominant species and different depths of soil moisture &alfalfa biomass
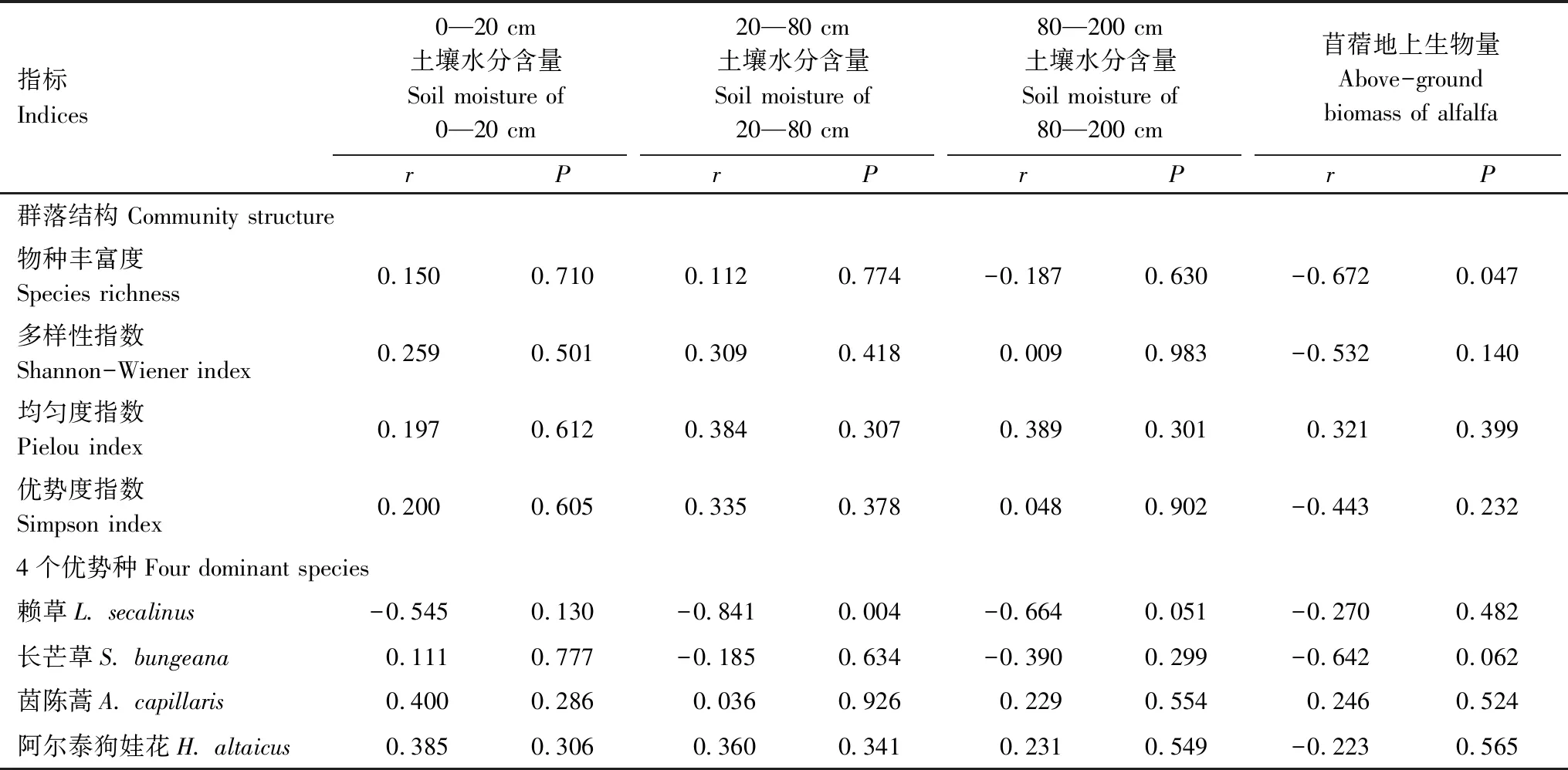
指标Indices0—20 cm土壤水分含量Soil moisture of 0—20 cm20—80 cm土壤水分含量Soil moisture of 20—80 cm80—200 cm土壤水分含量Soil moisture of 80—200 cm苜蓿地上生物量Above-ground biomass of alfalfarPrPrPrP群落结构 Community structure物种丰富度Species richness0.150 0.710 0.112 0.774 -0.187 0.630 -0.672 0.047 多样性指数Shannon-Wiener index0.259 0.501 0.309 0.418 0.009 0.983 -0.532 0.140 均匀度指数Pielou index0.197 0.612 0.384 0.307 0.389 0.301 0.321 0.399 优势度指数 Simpson index0.200 0.605 0.335 0.378 0.048 0.902 -0.443 0.232 4个优势种Four dominant species赖草L. secalinus-0.545 0.130 -0.841 0.004 -0.664 0.051 -0.270 0.482 长芒草S. bungeana0.111 0.777 -0.185 0.634 -0.390 0.299 -0.642 0.062 茵陈蒿A. capillaris0.400 0.286 0.036 0.926 0.229 0.554 0.246 0.524 阿尔泰狗娃花H. altaicus0.385 0.306 0.360 0.341 0.231 0.549 -0.223 0.565
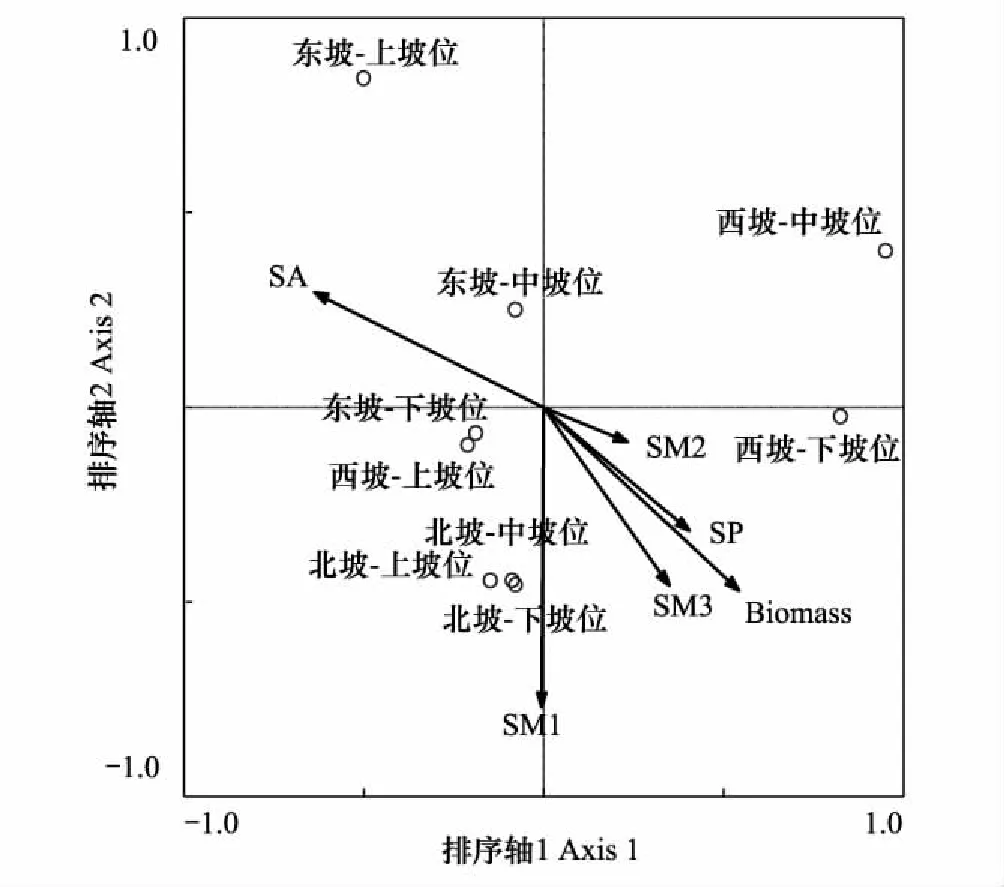
图3 地形、土壤水分和苜蓿地上生物量对其他草本群落分布影响的CCA排序图Fig.3 CCA analyses between topographical features, soil moisture content & above-ground biomass of alfalfa and herbage community distributionSA:坡向,Slope aspect;SP:坡位,Slope position;Biomass:苜蓿地上生物量,Above-ground biomass of alfalfa;SW1:0—20 cm土壤水分含量,Soil moisture content of 0—20 cm;SW2:20—80 cm土壤水分含量,Soil moisture content of 20—80 cm;SW3:80—200 cm土壤水分含量,Soil moisture content of 80—200 cm
苜蓿地上生物量与苜蓿重要值呈极显著的正相关关系(P<0.001),本文选用苜蓿地上生物量反映苜蓿的退化程度。苜蓿生物量、不同深度土壤水分及地形变化与其他草本群落的CCA排序结果表明,6个因子解释87.8%的草本群落变异(P=0.002)。第1排序轴与坡位(P=0.266)和苜蓿地上生物量(P=0.143)呈正相关,与坡向(P=0.044)呈显著负相关,它解释了30.1%的草本群落变异;第2排序轴与0—20 cm土壤水分呈显著负相关(P=0.039),与80—200 cm土壤水分和苜蓿的地上生物量也成负相关(P=0.219;P=0.268),它解释了20.4%的草本群落变异。从排序图中还可以看出,坡向、坡位及苜蓿地上生物量变化导致西坡草本组成与东坡和北坡不同,不同坡位0—20 cm、80—200 cm土壤水分和苜蓿地上生物量的变化导致西坡和东坡样带上坡位和中、下坡位草本群落组成相差较大(图3)。pCCA排序结果表明,坡向(解释量21.5%,P=0.053)、苜蓿地上生物量(21.2%,P=0.065)、0—20 cm土壤水分含量(19.1%,P=0.084)和20—80 cm土壤水分含量(19.1%,P=0.084)对草本群落分布的影响较大,而80—200 cm(10.5%,P=0.304)和坡位(10.3%,P=0.302)对草本群落分布的影响较小。
不同草本种对土壤水分及苜蓿退化的响应模式也不一致,赖草、长茅草、茵陈蒿和阿尔泰狗娃花是退化苜蓿草地常见的草本植物,在3条样带内均有分布且重要值较高。赖草与排序轴1负相关(P=0.239),与不同深度土壤水分和苜蓿地上生物量负相关(表4),它们解释了68.0%(P=0.068)的赖草重要值变化。长芒草与排序轴1负相关(P=0.007),与0—20 cm土壤水分正相关(表4),与20—80 cm、80—200 cm和苜蓿地上生物量负相关(表4),它们解释了11.0%(P=0.424)的长芒草重要值变化。茵陈蒿与排序轴1正相关(P=0.164),与排序轴2呈负相关(P=0.093),与不同深度土壤水分和苜蓿地上生物量均正相关(表4),它们仅解释了茵陈蒿9.0%(P=0.431)的重要值变化。阿尔泰狗娃与排序轴1和排序轴2均负相关(P=0.214;P=0.546),与不同深度土壤水分正相关(表4),与苜蓿地上生物量负相关(表4),它们解释了28.0%(P=0.295)的阿尔泰狗娃花重要值变化。
4 讨论
半干旱黄土区土壤水分是影响植被生长的关键限制因子[19-20],加之该区沟壑纵横,坡面地形变化改变了土壤水分的再分配,从而直接或间接地影响了植被生长。已有研究表明,坡面及微地形变化影响了土壤水分的有效性,在半干旱黄土丘陵区从坡顶到坡底不同土层深度土壤含水量变化趋势为先增加后持平或减小[21],同时塌陷、缓台、切沟、浅沟等微生境土壤水分要好于原状坡和陡坎[22]。此外,研究还发现植被恢复类型及年限不同,坡面土壤水含量亦存在差异[17,23-25]。总之,坡面人工植被恢复初期,随着植被盖度的增加会减弱土壤侵蚀,然而随着植被地上和地下生物量的增加,土壤水分出现亏缺将会抑制植物生长从而引起植被退化,进而影响人工植被群落组成及其群落物种多样性[15,26-30]。紫花苜蓿是典型的深根系、多年生草本,种植6—7a之后草地产量会迅速下降[31- 32],故半干旱黄土区大面积种植的紫花苜蓿因土壤水分的限制以及种植年限过长而逐渐衰退,加之后期缺乏合理的田间管理,加速了人工苜蓿草地退化。本文研究的苜蓿草地均处在退化阶段(栽植年限均为14a),但退化程度因坡向及地形部位不同而异。研究发现北坡样带属阴坡生长季内0—200 cm土壤水分(13.25%)条件好于半阳(阴)坡的西坡和东坡样带(11.72%和11.74%),其中北坡样带不同坡位间苜蓿的地上生物量相差较小,而西坡和东坡样带上坡位苜蓿地上生物量明显低于中下坡位。该结论与我们关于坡面地形及土壤水分变化对柠条生长影响的研究结果相类似[33],表明坡面地形引起土壤水分的差异进而影响了苜蓿草地退化过程。此外,研究还发现不同坡向苜蓿地上生物量与80—200 cm土壤水分呈显著正相关关系,而与生长季内0—80 cm土壤水分含量呈负相关关系,这在一定程度上说明苜蓿的退化有利于恢复0—80 cm土壤水分[16,34],同时苜蓿的生长会利用深层土壤水分,不同坡位深层土壤水分含量的高低会影响苜蓿的退化程度。Yang等[24]和杨永东等[35]针对半干旱黄土丘陵区苜蓿草地的研究发现,苜蓿的生长会导致100—200 cm间土壤水分亏缺,表明土壤水分持续减少是苜蓿退化的重要驱动要素,土壤水分含量在坡面上的变化与苜蓿退化程度密切相关。
坡面苜蓿的退化过程强烈影响了其他草本的分布及多样性。李裕元和邵明安[15]在陕西省神木县和王书转等[36]在甘肃省镇原县的研究均发现随着苜蓿草地的退化,草本的物种数开始增加,并且其群落也开始变化,草地退化前期物种丰富度、群落多样性在6a之前升高较快,6—10a达到最高,10a以上逐渐过渡到长芒草群落。本文的研究发现苜蓿地上生物量与其他草本地上生物量呈显著负相关,同时其他草本的物种丰富度、多样性和均匀度指数亦与苜蓿地上生物量呈负相关关系,这表明苜蓿的退化程度影响了其他草本的多样性,促进了草本群落从人工草地群落向天然草地群落的转变。此外,研究还发现不同坡向及坡位其他草本组成存在一定差异,这可能与不同草本种对地形及苜蓿退化引起的土壤水分变化有关。坡面退化苜蓿草地内赖草对土壤水分、长芒草对苜蓿地上生物量响应较为敏感,而茵陈蒿和阿尔泰狗娃花对土壤水分和苜蓿退化的响应均不敏感。茵陈蒿和阿尔泰狗娃花是流域内阳坡常见草本植物,常为长芒草等禾本科植物群落的主要伴生物种[32],其分布受地形和苜蓿退化的影响较小;而赖草和长芒草等禾本科植物则易受苜蓿退化引起的土壤水分变化的影响,苜蓿退化促进了人工苜蓿群落向长芒草群落的过渡[15,37]。该现象在研究区西坡样带表现最为明显,其上坡位苜蓿已完全退化,形成了以长芒草为优势种的次生天然草本顶级群落,而中、下坡位却未见长芒草,且苜蓿地上生物量下坡位显著高于中坡位,即紫花苜蓿与长芒草具有相互消长的种群动态关系,该研究结果与李裕元等[15]在陕北黄土高原的研究结果相似。总之,坡面苜蓿草地退化过程中,坡向和坡位等微地形可能改变土壤水分条件,其与苜蓿退化程度互馈互作影响了不同微地形下其他草本分布及多样性对苜蓿草地退化的响应模式[22,37-39]。
黄土区人工苜蓿草地退化改善了表层土壤水分,从而促进了一些浅根系草本的生长,驱动了天然草本群落的恢复演替过程。另一方面,地形变化改变土壤水分的时空分布格局,影响了人工苜蓿草地草本群落的恢复演替过程。本研究仅针对苜蓿草地退化后期地形和土壤水分变化对坡面草本分布及多样性的影响,后期的研究还需要借助控制试验和长期定位监测,以期系统地揭示退化人工植被生态系统中群落演替特征及其影响机制,为半干旱区人工植被恢复及可持续性管理提供科学依据。
5 结论
半干旱黄土丘陵区西坡、东坡和北坡人工苜蓿草地样带内苜蓿的地上生物量明显不同,西坡和东坡样带中、下坡位苜蓿地上生物量明显高于上坡位,而其他草本的地上生物量、物种丰富度和多样性指数的变化趋势则与之相反,这说明苜蓿的退化影响了其他草本的分布及多样性。土壤水分是影响苜蓿和其他草本生长的重要环境因子,80—200 cm土壤水分与苜蓿的地上生物量显著正相关;0—20 cm和20—80 cm土壤水分与其他草本地上生物量的相关性略高于80—200 cm土壤水分。地形、不同深度土壤水分和苜蓿地上生物量解释了其他草本群落变异的87.8%,其中坡向、苜蓿地上生物量、0—20 cm和20—80 cm土壤水分4个因子解释了79.3%的群落变异。此外,研究还发现不同草本物种对退化苜蓿草地地形变化、土壤水分及苜蓿地上生物量变化的响应模式不同,赖草对不同深度土壤水分的响应较敏感,长茅草对苜蓿地上生物量响应较敏感,而茵陈蒿和阿尔泰狗娃花对其响应不敏感。
猜你喜欢
文萃报·周二版(2022年23期)2022-06-11
现代畜牧科技(2021年4期)2021-07-21
农民致富之友(2019年33期)2019-12-20
中国交通信息化(2019年9期)2019-11-16
中国交通信息化(2017年6期)2017-06-06
小学生导刊(2017年14期)2017-05-17
绿色科技(2017年3期)2017-03-14
绿色科技(2017年1期)2017-03-01
植物营养与肥料学报(2012年3期)2012-10-26
