
唐古特白刺叶性状及叶片δ13C、δ15N沿降水梯度的变化特征
2019-07-05 10:59:48李永华辛智鸣刘明虎郝玉光刘丹一陈新均张正国
生态学报 2019年10期
董 雪,李永华,辛智鸣,刘明虎,郝玉光,刘丹一,陈新均,张正国
1 中国林业科学研究院荒漠化研究所, 北京 100091 2 中国林业科学研究院沙漠林业实验中心, 磴口 015200 3 库姆塔格荒漠生态系统国家定位观测研究站, 敦煌 736200 4 甘肃敦煌荒漠生态系统国家定位观测研究站, 敦煌 736200 5 内蒙古磴口荒漠生态系统国家定位观测研究站, 磴口 015200 6 中国治沙暨沙业学会,北京 100091
叶片性状是反映植物对环境生存策略的重要因子[1- 2]。近20年来,围绕植物适应性、竞争、演替等内容国内外进行了大量研究[3- 4]。总结现有研究表明,叶片形态(长、宽、长宽比、面积、厚度等)、比叶面积(比叶重)、叶氮含量之间存在显著的相关关系[5- 8]。同时,叶性状因子随水分、养分、辐射、温度等环境因子变化而变化[5,7]。最终,通过叶性状的调整实现对限制资源的优化利用,获取较高的光合生产能力。例如,随水分有效性减少或辐射(温度)的增加,叶片变小、变厚、比叶面积降低、叶氮含量增加,从而实现在减少单叶蒸发面积的同时提高光合速率,增加水分利用效率[4,9-10];在高水分环境、低光、低养分环境下,叶片变薄、比叶面积增加、叶氮含量降低,增加氮在光合组织投资比例,提高养分或光的利用效率[8,11];在低温环境下,叶片也出现变小、变厚、比叶面积降低、叶氮含量增加的特征,这一特征将有利于叶片保持较高的温度、减少低温的伤害,从而提高光合生产能力[12-13]。有关干旱区植物的研究仍存在滞后现象,现有研究结果表明,干旱区植物叶性状之间以及叶性状与环境因子(降水、温度)耦合关系变弱甚至消失[14-15],叶性状对植物的影响(如长期水分利用效率,δ13C)有别于其他区域的变化趋势[10,12]。这一混乱的特征可能与干旱区植物生境异质性的增加有关,从而无法应用单个环境因子或叶性状因子解释植物的生存适应特征[15-16]。
叶片稳定碳氮同位素,受生长环境的影响较大,用于指示和评估植物生理代谢与水分利用效率[17-18],可记录物源信息和环境变化[19]。13C同位素在一定程度上可以反映植物所受的水分胁迫程度和水分利用效率,荒漠区植物个体生长及对水分生理响应较明显,且在干旱年份随着地下水位的下降而下降[20],而环境变化和植物生理调节对植物个体、品种、群落之间的影响存在较大差异,叶片形态变化必将影响叶片瞬时或长期水分利用效率[21]。 植物δ15N除受其本身对氮的生理代谢过程控制外, 很大程度上还受各种环境因素的影响,因而δ15N值在植物种内和种间存在显著差异,其同位素组成在形成过程中能响应植物所处的环境变化[22]如气候、土壤类型等以海拔分异研究为主但变化趋势并不完全一致。因此, 探讨当前环境因子与植物碳氮同位素组成的关系,有助于揭示和预测未来全球变化对植物水分生理和生态系统氮素走向的影响, 同时也可以将获得的植物δ13C、δ15N与环境因子关系的结果应用于植被恢复重建。
为此,本研究选择干旱区常见优势种群唐古特白刺为研究对象,沿300—40 mm的降水梯度,设置7个区域进行调查取样,测定了不同降水环境下唐古特白刺叶片性状(长, 宽、长宽比、叶面积、比叶面积、叶氮含量)、叶片δ13C和δ15N值,同时结合环境因子如地下水埋深、降水、温度、温度等,系统分析了不同降水环境下唐古特白刺叶片性状、叶片δ13C和δ15N变化特征及影响其变化的环境因子。
1 材料与方法
1.1 样地设置与特征
在唐古特白刺群落天然分布区,沿降雨梯度(300—40 mm),设置了7个调查区(表1)。2013年至2015年的8—9月,我们在每个调查区内选择地势较为平坦的区域,设置了3个10 m×10 m调查样方,每个样方间隔50 m以上,并保证每个样方内至少包含1个没有明显退化特征的唐古特白刺沙包。通过实地勘测调查,获取样地周边地下水埋深数据,地下水取样点与调查点的距离不超过0.5 km。同时,利用国家气象局气象共享网站,分年度获取了7个研究区2013—2015年逐日气象数据,并依此计算了7个研究区采样当年的年平均降雨量,年平均气温、年平均相对湿度及年最高气温大于35 ℃日数。7个调查样地位置及区域环境特征见表1。
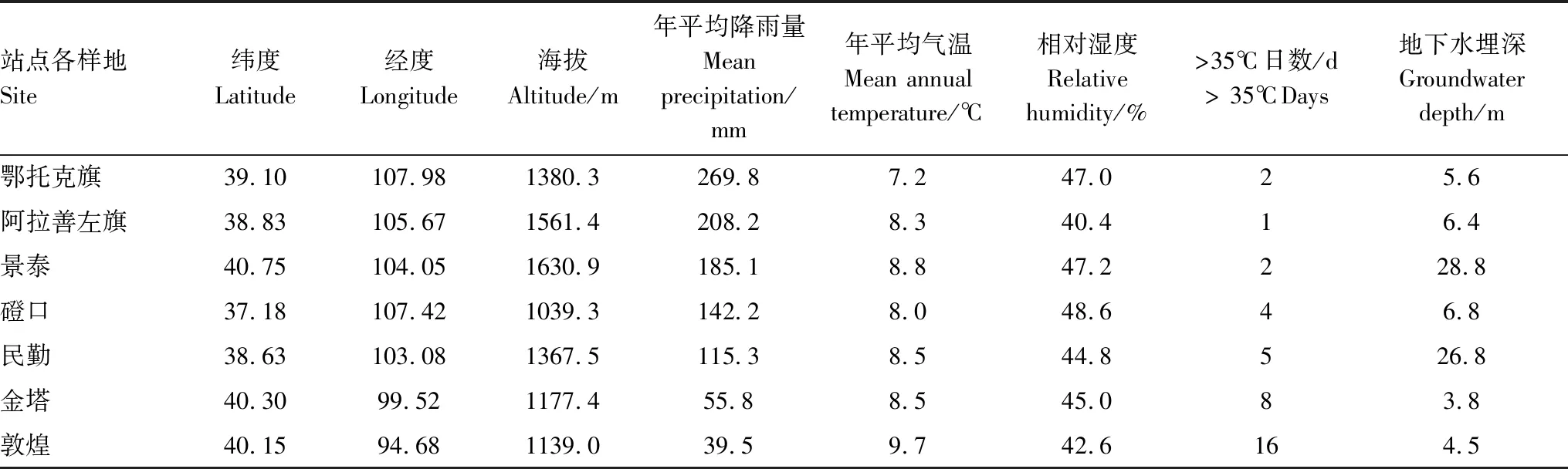
表1 样地位置与环境特征
所选择的7个样地以封育为主,人为干扰较少。地貌类型以固定或半固定沙地为主。随着降水减少,从鄂托克旗到敦煌,除唐古特白刺以外,其他优势物种从草本逐渐变化为半灌木、灌木;常见物种从多年生植物、沙生一年生短命植物逐步变化为沙生或盐生一年生短命植物;同时,随降水减少样地内物种数量也急剧减少(表2)。

表2 研究区7个样地特征
唐古特白刺NitrariatangutorumBobr.、白沙蒿Artemisiasphaerocphala、沙拐枣Calligonummongolicum、油蒿Artemisiaordosica、柽柳TamarixchinensisLour. 、白草PennisetumflaccidumGriseb、猪毛菜Salsolacollina、虫实Corispermumhyssopifolium、沙生针茅Stipacapillata、雾冰藜Bassiadasyphylla、沙鞭Psammochloavillosa、沙米Agriophyllumsquarrosum、芦苇Phragmitesaustralis、盐生草Halogetonglomeratus、狗尾草Setariaviridis(L.) Beauv. 苦马豆Sphaerophysasalsula(Pall.) DC、牛心朴子Cynanchumkomarovii、稗草Echinochloacrusgalli(L.).Beaiv.、三芒草AristidaadscensionisLinn
1.2 样品采集与测定
我们在每个样方内选择成熟且形态完整的35—45个叶片,应用测厚仪(Exploit-033004, Exploit, Yiwu, China)活体测定叶片厚度。同时,在每个样方内选择并采集成熟且形态完整的50—150个叶片,通过图片扫描(Canon LiDE120, Canon, Tokyo, Japan)、分析(Image-Pro Plus 6.0, Olympus, Tokyo, Japan), 获取叶片长、宽、面积等形态参数,并计算长宽比。叶片扫描完成后,带回实验室在75 ℃环境下烘干至恒重称其干重,并通过叶片重量与对应的叶面积计算每个样方内叶片的比叶面积。称重完成后,将叶片粉碎并过100目筛,最后将叶片送至厦门大学同位素实验室分析叶片δ13C、δ15N及单位重量的叶氮含量。
长宽比(L/W)=叶片长度/叶片宽度
比叶面积=叶片面积(cm2)/叶片干重(g)
1.3 数据处理
本研究利用SPSS 18.0 (SPSS Inc., Chicago, USA) 软件分析了叶片长、宽、长宽比、叶面积与叶片厚的平均值及其分布范围。由于比叶面积、叶氮含量、叶片δ13C和δ15N分析数据每个样区仅为3组,这里仅分析了4个指标在同一样区的平均值,而没有给出其分布区间。同时应用双变量相关分析检查了7个研究区环境因子(年平均降雨量,年平均气温、年平均相对湿度,年最高气温大于35 ℃日数、样地地下水埋深)与叶片性状(长, 宽、长宽比、叶面积、比叶面积、叶氮含量)、叶片δ13C和δ15N之间的相关关系;基于不同样区3个样方的测试数据,应用单因素AVONA分析检查了不同样区叶片性状、叶片δ13C、δ15N之间的差异性。应用Origin 8.5 (OriginLab Corp., Northampton, MA, USA)线性回归拟合了环境因子及叶性状对叶片δ13C、δ15N的影响。
2 结果与分析
2.1 不同区域唐古特白刺叶片性状、δ13C、δ15N 的分布特征
数据分析表明,唐古特白刺叶片形态在同一样区及不同样区间均表现在出较大的变异性。同一样区叶片长度变化幅度在2.4—4.7倍之间,全部样品叶片长度分布区间为0.8—5.1 cm,变幅为6.4倍;同一样区叶片宽度变化幅度在2.6—4.4倍之间,全部样品叶片宽度分布区间为0.3—1.7 cm,变幅为5.4倍;同一样区叶片长宽比变化幅度在2.0—3.3倍之间,全部样品叶片长宽比分布区间为1.5—5.7,变幅为3.8倍;同一样区叶面积变化幅度在4.6—15.0倍之间,全部样品叶片面积分布区间为0.2—3.7 cm2,变化幅度为19.4倍;同一样区叶片厚度变化幅度在1.6—2.4倍之间,全部样品叶片厚度分布区间为0.5—1.7 mm,变化幅度为3.4倍(表3)。单因素方差分析表明叶片宽度、长宽比、叶面积与叶片厚度在不同样区间差异显著,而叶片长度差异不显著。样区数据分析表明,7个样区比叶面积、叶氮含量、叶片δ13C和δ15N值的平均值分布区间分别为69.9—85.5 cm2/g,2.3%—3.6%,-25.1‰—-27.4‰,3.6‰—11.5‰(表3),其中以阿拉善左旗的比叶面积和叶氮含量最高,显著高于其他分布区域,而敦煌最低。由于δ13C值由大到小的顺序是:民勤>景泰>磴口>阿拉善左旗>鄂托克旗> 敦煌>金塔,而δ15N值是:敦煌>景泰>金塔>阿拉善左旗>民勤>鄂托克旗 >磴口,说明7个区域的唐古特白刺叶片δ13C和δ15N值的变化趋势不同。单因素方差分析表明,不同样区间比叶面积、叶氮含量、叶片δ13C和δ15N值差异显著。总之, 荒漠植物唐古特白刺不同分布区自然种群中叶片特征显著不同。
2.2 环境因子对唐古特白刺叶性状的影响
相关分析表明,7个样区环境因子间,除温度与降水关系较为密切外,降水、温度与空气相对湿度、地下水埋深之间没有显著的相关关系,但与年最高气温大于35℃日数呈显著相关(表4)。除叶片长度和叶片宽度与叶面积呈极显著正相关,叶片厚度与单位重量的叶氮含量呈显著负相关,其他叶性状间均无显著相关性。环境因子对叶片性状的影响方面,叶片厚度与温度显现出显著正相关关系(P<0.05),尤其与年最高气温大于35℃日数的关系更为密切,达到极显著正相关(P<0.01),相关系数最大达到0.96;同时随温度的升高,单位重量的叶氮含量呈降低趋势,两者间呈显著负相关(P<0.05);可以说年最高气温大于35℃日数是影响唐古特白刺叶总体特征的主要限制因子。除此之外,环境因子对叶片性状影响并不显著(表4)。
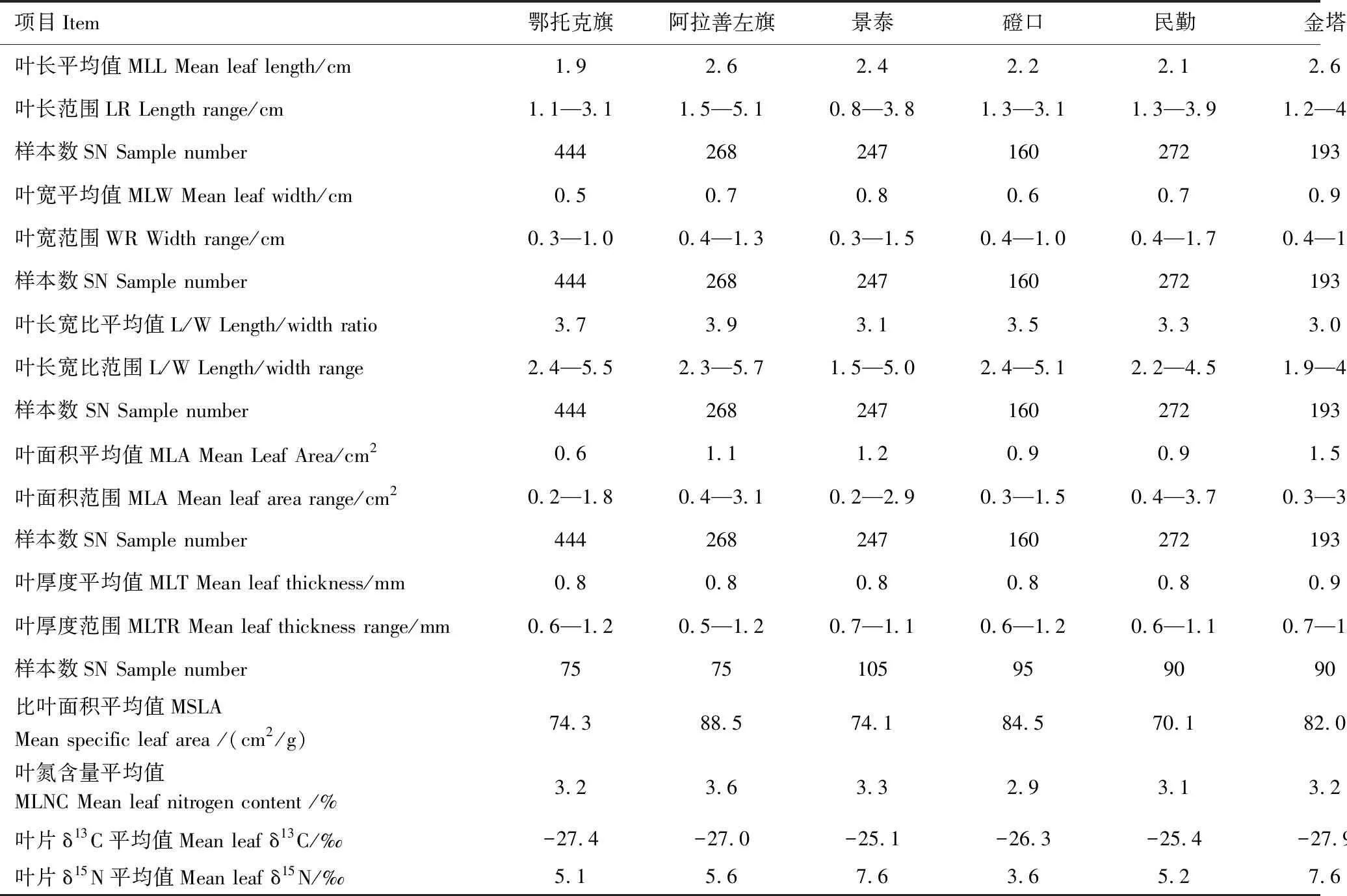
表3 不同地区唐古特白刺叶片性状、叶片δ13C、叶片δ15N 分布特征
*:<0.05;**<0.01

表4 环境因子与叶性状间的相关分析
MAP:年平均降雨量,Mean Annual Precipitation;MAT:年平均温度,Mean Annual Temperature; MAH:年平均相对湿度,Mean Annual Relative Humidity;Tmax- 35:年最高气温大于35 ℃日数,> 35 ℃ Days;DG:地下水埋深,Groundwater Depth;LT: 叶片厚度,Leaf Thickness;LL:叶片长度,Leaf Length;LW:叶片宽度,Leaf Width;L/W:叶片长宽比,Length to Width Ratio; LA:叶面积,Leaf Area;SLA: 比叶面积,Specific Leaf Area;Nmass:叶氮含量,Leaf Nitrogen Content;*,P< 0.05; **,P< 0.01
2.3 叶片δ13C、δ15N的变化特征
相关分析显示(表5),除地下水埋深外与叶片δ13C值相关关系极显著(P<0.01)外,其他环境因子及叶片性状因子对叶片δ13C值变化影响不明显(P>0.05);而叶片δ15N值仅与年平均气温、年最高气温大于35 ℃日数和叶片厚度呈显著正相关(P<0.05),与其他环境因子及叶片性状因子无关。叶片δ15N值与叶片厚度对温度的响应显著且一致。

表5 叶片δ13C、δ15N与环境因子、叶性状间的相关分析
δ13C:碳同位素比值,Leaf δ13C Content; δ15N:氮同位素比值,Leaf δ15N Content; *,P< 0.05; **,P< 0.01
线性回归分析显示,地下水埋深是导致叶片δ13C值变化的关键因子,二者呈现显著的正相关关系(R2=0.84,P<0.01)(图1)。由于磴口有黄河和众多湖泊的影响, 存在多种生态系统,唐古特白刺沙包土壤中有明显的粘土层分布,造成对地下水分利用的影响,因此如果排除磴口的数据,地下水埋深变化能够解释唐古特白刺叶片δ13C变化的97% (P<0.001)。另外,高温环境下,叶片增厚的同时叶氮含量显著降低(表4),而这一过程中叶片δ15N值却有增加趋势(R2=0.62,P<0.05)(图2),7个样区的唐古特白刺通过权衡叶片厚度、叶氮含量与δ15N值的关系来消除或者降低不适宜温度对自身的伤害作用。
3 讨论与结论
3.1 环境对唐古特白刺叶片性状变化的影响
叶片性状随着环境(温度、降水、相对湿度)变化而变化,反映了植物对不同气候状况下环境选择的适应性[23- 25]。不同区域同一种植物,因年均温度(MAT)和年均降水量(MAP)等气象因子的差异,会产生叶片形态变异,形成不同的叶性状[25- 27]。区域尺度的光照、土壤及降水等环境因子各不相同,造成热量和水分重新分配,使植物生长条件更为复杂,也使叶片性状对环境条件产生复杂的适应性变化[28-29]。本研究中,唐古特白刺功能性状主要受降水和温度的影响,其中年最高气温大于35℃日数是主要的影响因素,这与孟婷婷等[30]认为温度和降水是植物功能性状变化的主要驱动因素的结论一致。唐古特白刺群落7个天然分布区,沿着年均降雨量逐渐降低的梯度,气候干旱加剧,温度升高,进而影响唐古特白刺的生长和发育,为了更好地适应干旱的环境,通过叶片的可塑性以形成对水分和养分的吸收利用策略和对环境的防御策略。7个唐古特白刺天然分布区沿降雨梯度,叶片功能性状属性值(除叶片长度以外)均具有显著差异(P<0.05),变化较大叶片厚度和叶氮含量与植物利用资源的能力密切相关,唐古特白刺通过叶片各功能性状间的调节来适应环境的变化,并形成性状间的最佳功能组合。
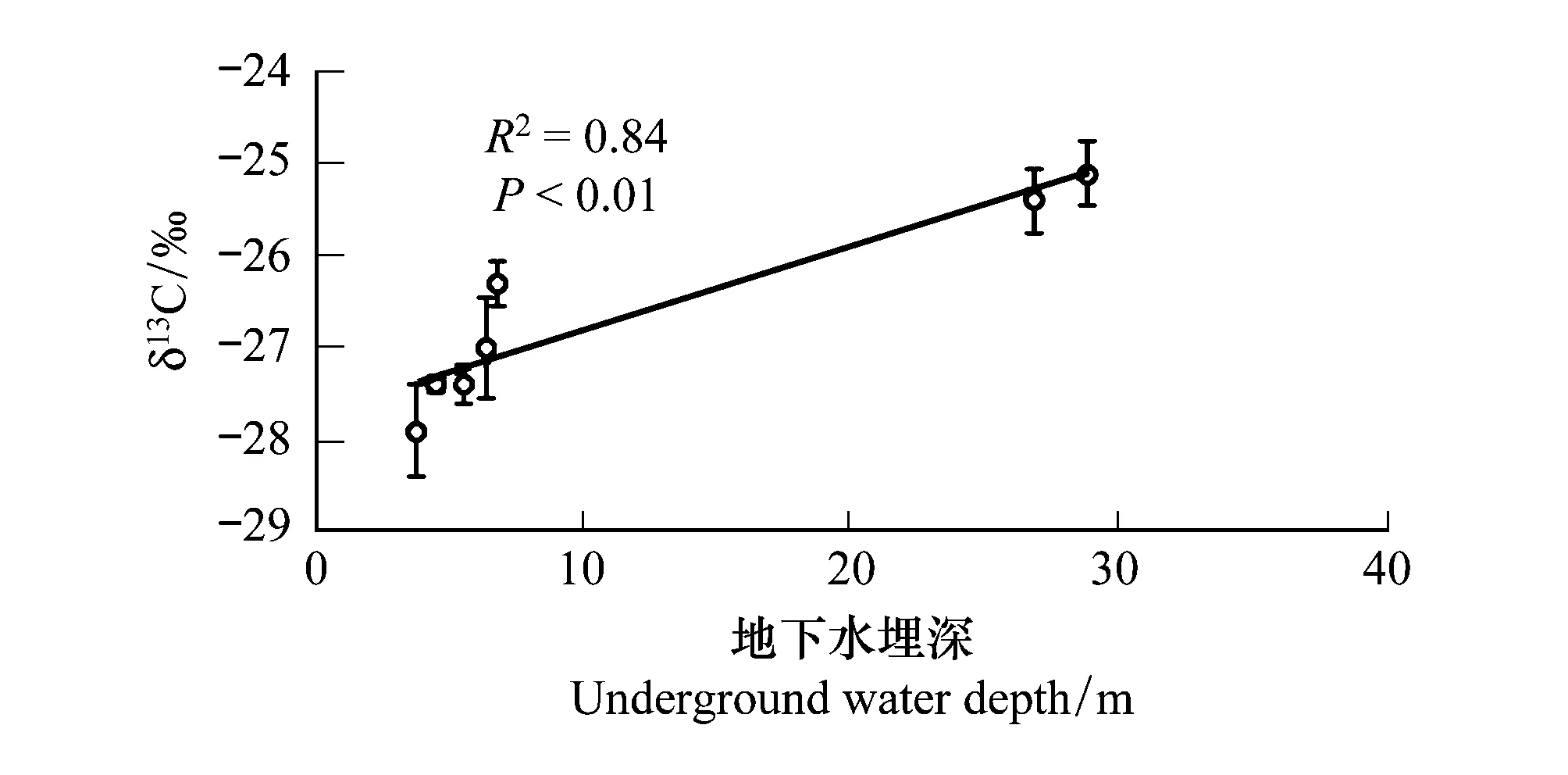
图1 地下水埋深对叶片δ13C值的影响 Fig.1 The effect of underground water depth on the leaf δ13C content
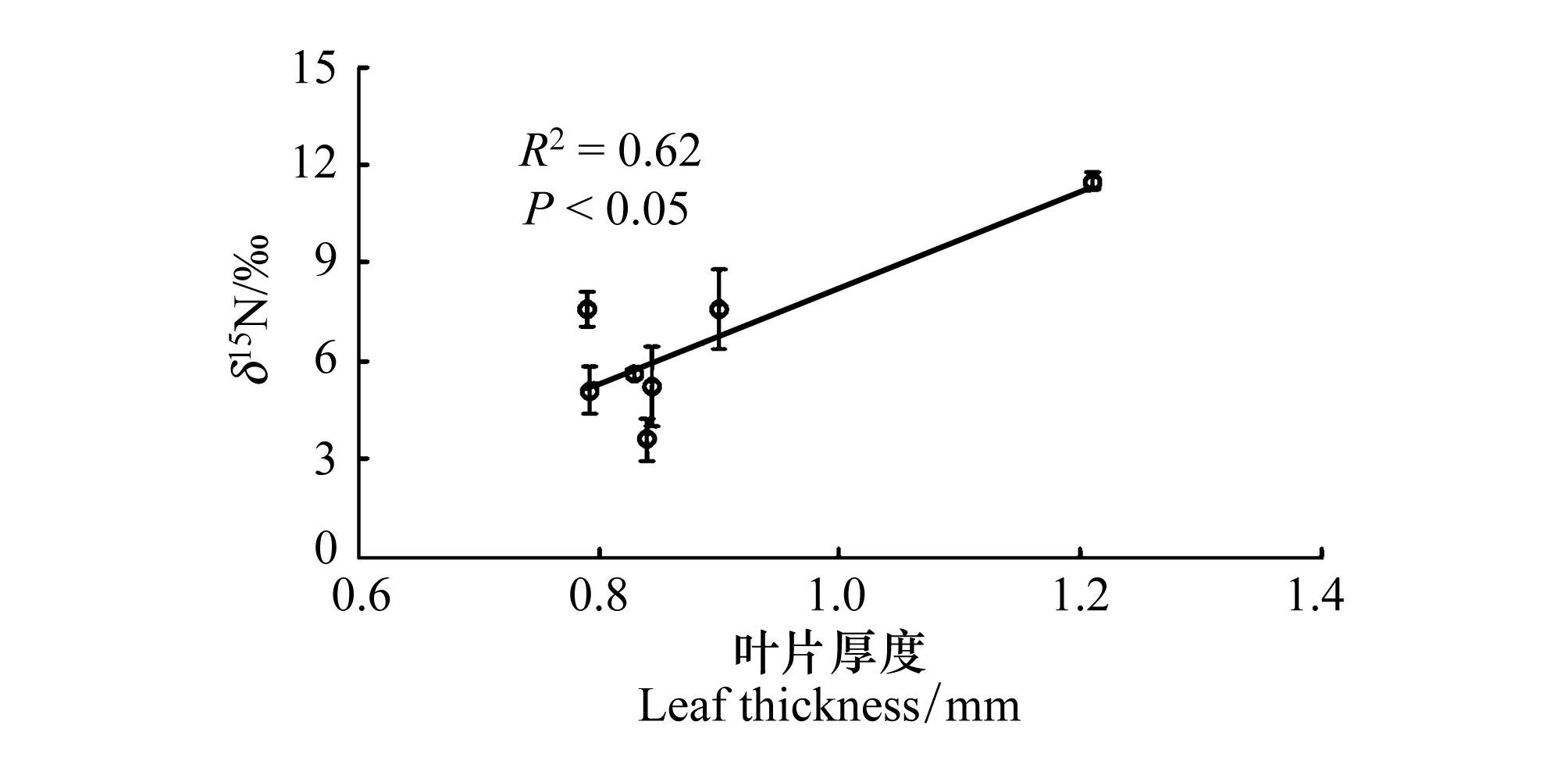
图2 叶片厚度对叶片δ15N值的影响Fig.2 The effect of leaf thickness on the δ15N content
Reich和Oleksyn[31]基于全球452个站点1280种植物的叶性状分析,发现在年平均气温>10℃时,Nmass随温度增加呈现降低趋势, 但当温度<10℃时, Nmass随温度减少而降低或不变。本研究发现,Nmass与年最高气温大于35℃日数(表4)呈显著负相关, 这一现象在该温度范围内与全球格局相一致[31-32]。在本研究中,比叶面积和叶氮含量并没有表现出显著的正相关关系,这可能与选择的样点间的综合环境因子有关。叶氮含量主要受温度影响,降雨没有成为其限制因子(表4)。因此,在降雨不成为叶氮含量变化的决定性因子时,叶氮含量的变化主要是对温度变化的一种适应策略。唐古特白刺对干旱贫瘠环境的适应主要表现为: 通过提高叶面积和叶片厚度来适应干旱环境,同时可通过降低比叶面积和叶氮含量适应温度较高的环境。尽管相关分析表明环境因子(降水、温度、相对湿度和地下水埋深)与SLA、Nmass并无直接的显著相关关系,但叶厚度与温度表现出了一定的相关关系,特别是年最高气温大于35 ℃日数与叶片厚度达到极显著相关,说明唐古特白刺主要采取改变叶厚度的方式来适应干旱、高温环境。
3.2 影响唐古特白刺叶片长期水分利用效率的关键因子
δ13C既由植物本身的生物学特性决定,同时又受到外界环境因素的影响。环境因子(如大气温度、降雨量和土壤含水量)能改变植物组织的碳同位素组成,通过对叶导度或光合速率的改变,或两者同时改变进行调控。较厚的叶片能够减少水分的散失,提高叶片水分利用效率[33]。另外,比叶面积的降低还被认为是植物通过增加叶片密度来提高对水分的保持力以抵抗水分散失的一种途径,比叶面积一般与植物的叶片光合能力和相对生长速率密切相关,而且受到植物体内水分和碳含量的影响,植物叶片比叶面积大小与植物水分利用效率存在显著负相关关系[34],较低的比叶面积一般伴随着较高的水分可利用性,同时还能减少外界对自身带来的损害[35]。干旱区植物生存环境的水分条件是影响植物功能性状变化的重要因素。降水量、空气湿度和土壤含水量三者是相互关联的,其中任何一个都可用来衡量植物可利用的水分。随着降水的减少,空气湿度以及土壤的含水量降低、水分的胁迫加重,从而植物将关闭部分气孔以避免水分的丢失。荒漠区植物根系分布范围和深度决定其水分获取途径、水分生理响应和适应特性,当降雨强度较小时,根系主要从稳定的深层土壤吸收水分,但随着降水强度的增加也可从上层土壤吸收水分[36-37]。在干旱、半干旱区降雨强度一般均较小且波动性强,唐古特白刺可利用的土壤有效水主要来自深层土壤水分,因此结果表明叶片δ13C只受地下水埋深的影响,其他环境因子和叶性状对其影响均未达到显著水平。
韩家懋等[38]研究认为C3植物的δ13C值为-23‰—-32‰,C4植物为-6‰—-19‰,本研究中7个唐古特白刺天然分布区叶片δ13C范围介于-25.1‰—-27.9‰之间,唐古特白刺叶片δ13C值落在C3植物区。任书杰等[39]研究表明中国区域478种C3物种叶片δ13C的变化范围为-22.00‰—-33.50‰,叶片δ13C变化范围也在此范围内。此外,唐古特白刺叶片δ13C平均值为-26.6‰,接近于刘光琇等[40]已经报道的青藏高原北部C3植物叶片δ13C值(-23.8‰—-29.5‰)和周咏春等[41]关于青藏高原草地群落植物叶片δ13C值(-25.88‰—-26.55‰)的研究,由于青藏高原属于高原气候,海拔较高,降水相对不足,植物在面对长期的干旱逆境过程中,产生了提高水分利用效率的生态适应策略,即表现出了较高的叶片δ13C值,而且唐古特白刺叶片δ13C平均值高于中国区域灌木植物叶片δ13C平均值(-27.50‰),说明荒漠植物为了适应干旱恶劣的环境,与高寒地区的植物形成了同样的抗逆生存策略。本研究中7个站点不同地区唐古特白刺叶片δ13C存在显著性差异(P<0.01),表现出了明显的空间变异性。唐古特白刺叶片稳定碳同位素值对不同环境条件的这种响应模式支持了唐古特白刺是一种以提高水分利用率而适应极端荒漠生境的典型超旱生植物。大量研究表明,植物δ13C随水分的增加而降低,植物δ13C的变化不仅包含了降水因素的影响,也包含了影响水分条件的其他环境因子的影响,它们的影响可能会叠加在直接的水分影响之上[42-43],本研究表明其他环境因子及叶片性状因子对叶片δ13C值变化影响不显著,只有地下水埋深与叶片δ13C值相关关系达到显著水平(P<0.01)。
关于叶片性状与δ13C值的关系研究认为影响叶片δ13C值变化的生理机制主要有两种:光合效率[44]和气孔限制[45]。而据Ares等[45]研究发现,在野外和温室环境下,柳叶桉和金合欢叶片的δ13C值与叶片性状均没有显著相关性,说明这两种植物δ13C值的变化主要受气孔限制的影响,且不是由光合效率的改变引起的。从这个角度来讲,唐古特白刺叶片碳同位素δ13C与叶功能性状指标的相关性分析表明(表5),叶片δ13C与叶面积、比叶面积总体呈负相关关系,与叶氮含量总体呈正相关关系,但其相关性均未达到显著性水平(P>0.05),仅地下水埋深与叶片δ13C表现出了极显著的正相关关系(P<0.01),可以推断唐古特白刺叶片δ13C值变化的主要影响因素是气孔限制,并不是光合效率,从而叶片形态指标和叶氮含量变化并不直接对叶片δ13C值产生影响。唐古特白刺在降水量小于100 mm的区域,主要利用地下水源,成为隐域植被,从而降低了对其他环境因子的响应,而当受到环境胁迫时,气孔的调整将可能是影响植物水分利用的关键因子,从而弱化了δ13C值与叶片其他性状的相关性,但其具体原因和影响机制有待进一步研究。唐古特白刺叶片特征对不同环境因子的响应模式在一定程度上反映和指示了唐古特白刺主要通过气孔调节以提高水分利用效率而适应极端气候(高温)的荒漠生境。
3.3 叶片δ15N值变化特征
氮是影响和限制植物生长最重要的营养元素之一。植物叶片稳定性氮同位素组成(δ15N)在很大程度上受到植物生长环境(如:叶片N含量、植物氮源、温度、降水、土壤氮有效性等)的影响,气候变化持续不断地对氮循环产生影响,植物叶片δ15N值可以在一定的时间和空间上揭示与植物生理生态过程相联系的一系列气候环境信息[22]。植物δ15N 与水分可利用性相关[46], 在全球尺度和较小的区域范围内,人们普遍发现植物叶片δ15N 随着降水量的增加呈现递减趋势[46-47]。温度也是影响植物δ15N值的重要气候因子,大量研究表明[47-49],陆地植物δ15N值与其生长温度呈正相关,即随温度升高,植物叶片δ15N值增加。但是,也有另外的研究发现与此不一致的结论[50-51]。许多研究已经在不同空间尺度上证明叶片N浓度与其δ15N正相关[51-52],是因为与叶片N浓度低的植物相比,叶片N浓度高的植物生长的地区土壤N有效性高,而土壤N有效性高的地区土壤N循环更加开放,使得植物叶δ15N增大。可见,现有关于叶片δ15N与叶片N浓度正相关的解释都是基于叶片N浓度可以反映土壤N有效性。然而,有研究却发现在某些地区叶片N并不能够反映土壤N的有效性,在高寒地区,植物为了适应严酷的生存环境(低温、低土壤N有效性),而使更多的N分配到叶片的光合器官内,以及生长速率降低,对植物N浓度的稀释作用降低,均可导致随着土壤N有效性降低,叶片N含量反而增加[53]。本研究指出决定δ15N的关键因子是根系主要吸收利用层土壤养分结构,降水量小于100 mm区域,根系主要利用地下水,该区域生态系统物质循环缓慢,根系接触的土壤层发育不充分导致该区域叶氮小,但唐古特白刺根际可培养固氮细菌类群,使得土壤N有效性较高,因此叶片δ15N高。
猜你喜欢
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:48:06
新疆农业科学(2020年1期)2020-02-14 03:22:46
农学学报(2018年10期)2018-11-01 01:19:30
中成药(2018年7期)2018-08-04 06:04:06
西夏学(2018年1期)2018-04-29 09:09:02
西夏研究(2017年1期)2017-07-10 08:16:55
青苹果·教育研究版(2016年7期)2016-12-12 05:19:52
西南农业学报(2016年5期)2016-05-17 05:42:28
中国麻业科学(2015年5期)2015-12-28 06:22:11
天然产物研究与开发(2014年6期)2014-04-27 14:15:58
