
沙地樟子松天然林南缘分布区林木竞争、空间格局及其更新特征
2019-07-05 10:59潘磊磊KWONSEMYUNG刘艳书杨晓晖朱媛君时忠杰
生态学报 2019年10期
潘磊磊,KWON SEMYUNG ,刘艳书,张 晓,杨晓晖,山 丹,朱媛君,时忠杰,*
1 中国林业科学研究院荒漠化研究所,北京 100091 2 北京林业大学水土保持学院,北京 100083
林木的生长与更新受到许多生物和非生物因素的影响,其中,竞争和空间格局是影响林木生长、林分结构及其动态的重要因素,在种群更新与演替中发挥着重要的作用,是生态学和林学研究的重要问题之一[1- 2]。竞争普遍存在于森林生态系统中,邻近的林木个体之间通过竞争获取更多的光照、水分和养分等资源,从而影响邻近个体对资源的利用[3-5]。而林木空间格局作为种群分布的外在表现形式,是其分布格局及其属性在空间上的排列方式[6],表达了林木间树种、大小、分布等空间关系,决定了树木之间的竞争优势及其空间生态位,是种群结构、动态和生态学过程、种内种间关系及外界环境相互作用的结果[7-8]。
近年来,许多学者对于林木的竞争[9-10]、空间格局[11-12]和天然更新[13]等进行了较为深入的研究。在林木竞争方面主要集中于种内竞争[14-15]、种间竞争[16-17]、种间差异[14]及其资源利用特性[18]、外界干扰的影响[19]等,而林木空间格局研究主要分析种群生物学特性[20]、种内和种间关系[21]以及植物与环境之间的相互作用和空间分布状态[22-23]等。而林分的更新可能受林木的竞争与空间格局等因素的影响,但对于林木竞争及空间格局与群落更新的关系研究却甚少,制约了对群落更新机制的认识。
我国的樟子松天然林主要分布在大兴安岭山地北部和呼伦贝尔森林草原过渡带的沙地上,其地理分布的最南缘位于呼伦贝尔沙地的南端。由于樟子松具有耐寒、耐旱、根系发达及不苛求土壤水分等特征,也被广泛应用到我国北方沙地造林中,对于防风固沙、农田防护等方面发挥着重要的作用。国内对于沙地樟子松的空间格局、竞争与更新等方面已有较多研究,特别是火干扰对竞争、格局与更新的影响方面[19,24- 25],如喻泓等[19]发现樟子松的竞争压力主要来源于种内,幼树多呈聚集分布特征,大树多呈随机分布特征,而火干扰是樟子松林分天然更新的主要驱动因子之一[24]。但对于天然沙地樟子松林纯林林木竞争、空间格局与更新的关系研究并未深入。为此,本研究针对天然沙地樟子松地理分布最南缘区的纯林,旨在充分认识林木之间的竞争关系、空间格局及其天然更新特征,厘清林木竞争与空间格局对天然更新的影响机制,为樟子松天然林的经营与保护及人工林的营造与管理等提供科学的指导。
1 研究区概况
研究区位于呼伦贝尔沙地东南端、新巴尔虎左旗罕达盖苏木中国蒙古边界附近(47°38′—47°39′N,119°09′—119°12′E)。呼伦贝尔沙地位于我国东北西部,东西长约270 km,南北宽约170 km,面积约1万km2,本区属中温带半干旱大陆性气候,夏季温暖,冬季严寒、日照充足,昼夜温差大,年平均气温0.2 ℃,年降水量280—400 mm左右,多集中在夏秋季,年蒸发量1400—1900 mm,无霜期90—100 d,积雪期长达140 d左右[26]。地貌类型以固定沙丘和半固定沙丘为主,土壤以风沙土为主[27]。植被以森林和草原为主,该区是沙地樟子松(Pinussylvestrisvar.mongolica)种群在我国地理分布的最南缘。本区的樟子松林多以纯林为主,伴生种有白桦(Betulaplatyphylla)、榛子(Corylusheterophylla)等,以及线叶菊(Filifoliumibiricum)、大针茅(Stipagrandis)、羊草(Leymuschinensis)、日阴菅(Carexpediformis)、地榆(Sanguisorbaofficinalis)等杂类草[28]。

表1 样地基本情况
2 研究方法
2.1 数据获取
2017年7—9月,在研究区选取人为干扰较小的沙地樟子松天然林,建立2个密度(DBH>5 cm林木株数/hm2)相似的50 m×50 m的样地,并每木检尺,对所有树高2 m以上的树木统一编号,测定其胸径、树高、枝下高、冠幅(按东、南、西、北4个方位)等林分指标,而树高小于2 m的树木则测定其基径,并利用全站仪(NTS 340,南方测绘仪器有限公司)测定所有林木空间位置坐标。样地基本信息如表1,调查样地内的林木空间坐标信息见图1。
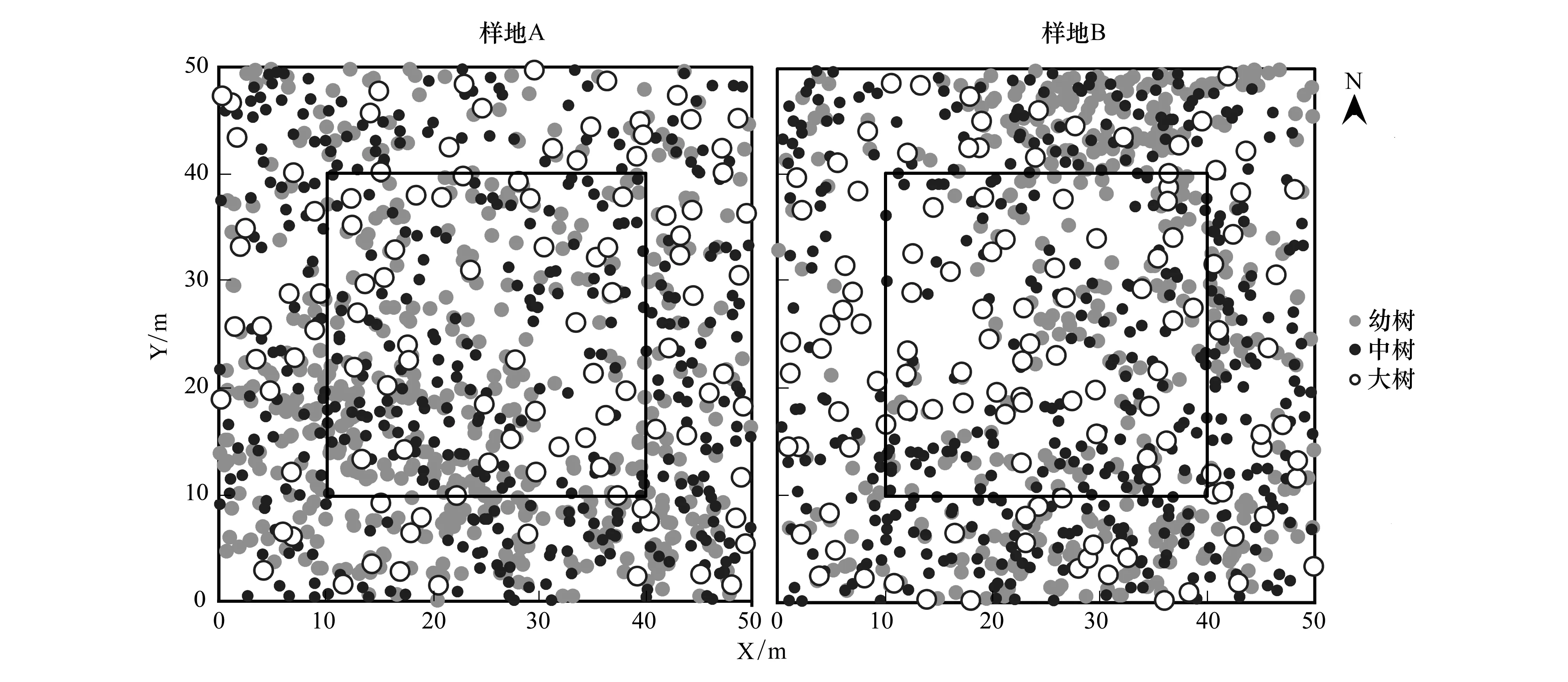
图1 样地林木分布Fig.1 Maps of trees distribution in sampling sites点代表树木,点的大小代表树木胸径,实心灰色、实心黑色、空心圆分别代表幼树、中树和大树;样地内圈黑色实方框指代为消除边缘效应,最终用于林分结构、竞争和更新分析的样方大小
2.2 林分结构与对象木的选择
将两个样地内对象木的胸径从0 cm开始统计,以3 cm为间距,划分为13个径阶:径阶I,DBH<3 cm;径阶II,3 cm≤DBH<6 cm;径阶III,6 cm≤DBH<9 cm;径阶IV,9 cm≤DBH<12 cm;径阶V,12 cm≤DBH<15 cm;径阶VI,15 cm≤DBH<18 cm;径阶VII,18 cm≤DBH<21 cm;径阶VIII,21 cm≤DBH<24 cm;径阶IX,24 cm≤DBH<27 cm;径阶X,27 cm≤DBH<30 cm;径阶XI,30 cm≤DBH<33 cm;径阶XII,33 cm≤DBH<36 cm;径阶XIII,36 cm≤DBH<39 cm,统计各个径阶的植株数,绘制径阶分布图。将对象木的树高从0 cm开始,按2 m为一个高阶,划分为8个高阶:高阶I,H<2 m;高阶II,2 m≤H<4 m;高阶III,4 m≤H<6 m;高阶IV,6 m≤H<8 m;高阶V,8 m≤H<10 m;高阶VI,10 m≤H<12 m;高阶VII,12 m≤H<14 m,统计各个高阶的植株数、绘制树高分布图。为了分析樟子松不同生长阶段的空间分布格局特征,结合实地调查樟子松的生活史特征,将樟子松划分为3个生长阶段:幼树,DBH<9 cm;中树,9 cm≤DBH<18 cm;大树,DBH>18 cm。为了消除边缘效应,将样地划分为25个10 m×10 m的样方(图1),选择中间9个样方内的所有林木作为对象木开展林分结构、竞争和更新分析。
2.3 竞争分析
竞争是个体或群体之间对生存空间与资源利用的关系,目前对个体竞争强度的研究较多,并提出了许多竞争模型,包括与距离相关的和与距离无关的竞争指数[29-30],但以Hegyi[31]的竞争模型模拟效果最好,应用最广泛[32-33]。因此,本研究采用了简便易算的与距离相关的Hegyi的竞争指数,其公式为:
(1)
(2)

一般认为,当植株的树冠或根系发生接触或重叠时,才会发生竞争,所以对象木所受到的竞争压力主要来自于其周围一定距离内的竞争木,而当超出这一距离,其对对象木的竞争逐渐减弱或消失。为此,本文采用固定半径法,选定A样地35株和B样地30株对象木,以2 m为起点,以1 m为样圆半径增量,分别计算2—20 m范围内每个样圆半径对象木的平均竞争指数,做出样圆半径与平均竞争指数的散点图,分析不同样圆半径与平均竞争指数的关系,随着样圆半径的增加,当竞争强度变化不显著时即确定为对象木的竞争范围Lij[34]。
利用樟子松对象木的竞争强度与其胸径进行回归分析,其幂函数回归拟合关系式为:即CI=a×D-b,式中CI为竞争指数,D为对象木的胸径,a,b为模型参数。当P<0.05时,达到显著水平;当P<0.01时,达到极显著水平。
2.4 点格局分析
点格局相关理论是由Ripley[35]首先提出,后经Diggle[36]等人发展,已被广泛应用于种群的空间分布格局以及种内、种间空间关联性分析中。Ripley′sK(r)函数是一个积累分布函数,在应用过程中,常因为积累效应的存在影响到结果的准确性[37-38]。Ripley′ sK(r)函数如下:
式中,A是研究区(样地)的面积,uij为2个点i和j之间的距离;Ir(uij)为指示函数,当uij≤r时,Ir(uij)=1,
当uij>r时,Ir(uij)=0;wij为权重值,用于边缘校正。
成对关联函数(pair correlation function)简称为g(r)函数来源于Ripley′sK函数,但它可以较好地排除K(r)函数引起的累积效应,具有更广的适用性[39-40]。本研究选择g(r)函数分析不同物种的空间分布特征以及空间关联性。g(r)函数与Ripley′K(r)函数之间的关系为:
g(r)=(2πr)-1dK(r)/dr
式中,dK(r)是函数K(r)的微分,dr是半径r的微分。
种间或种内的空间关联可以运用双变量点格局分析,即:
g12(r)=(2πr)-1dK12(r)/dr
该公式计算以关联物种或群体1的每个个体为圆心,半径为r的圆环区域内关联物种或群体2的数量[41- 43]。
在单变量g(r)分析时,g(r)值落在上下包迹线之间为随机分布,在上包迹线以上为显著的聚集分布,在下包迹线以下为显著均匀分布。双变量g12(r)可以用物种种间以及同一物种的不同生长阶段之间的空间关联性,按照植物的生长发育规律,大树会对中树、幼树的空间分布特征产生影响[38]。所以在进行双变量分析时,保持大树的位置不变,中树和幼树随机变化;中树位置保持不变,幼树随机变化。分析时,g12(r)值落在上下包迹线之间,二者之间无相关性;在上包迹线以上为显著的正相关;在下包迹线以下为显著的负相关[44]。
2.5 种群更新分析
为了探讨植株的竞争强度及其空间分布格局与种群之间的关系,将样地内胸径0—9 cm的幼树作为更新苗,选择样地内胸径18 cm以上对象木作为母树,以单株母树为中心,统计2、4、6、8 m样圆半径范围内4个方向(E、S、W、N)的更新苗的存活数及死亡数。A样地选取了37株对象木,B样地选取了44株对象木。
2.6 统计分析
点格局分析通过Programita 2014软件完成,为降低边缘效应对分析的影响,在本研究中空间尺度选择样地边长的一半(0—25 m),步长为1 m,采用Monte-Carlo检验拟合199次,产生置信水平为99%的上下包迹线以检验分析结果的显著性。其他统计分析都在Excel 2013和SPSS 19.0软件完成。
3 结果与分析
3.1 对象木径阶与树高结构
A样地共调查了368株对象木,最小胸径0.8 cm,最大胸径38.7 cm,平均胸径为9.1 cm;B样地共调查了285株对象木,最小胸径为1.2 cm,最大胸径36.8 cm,平均胸径11.0 cm。如图2所示,中小径阶沙地樟子松林木所占比例较高,A样地径阶I—VII(0—21 cm)的株数为355株,占总株数96.47%,B样地的径阶I—VII为267株,占总株数94.35%;A样地、B样地径阶均呈正态分布(Kolmogorov-Smirnov 正态性检验,P值分别为0.696和0.619)(图2),两个样地的樟子松种群均属于增长型种群结构。且两个个样地的高阶分布型非常相似(图2),其平均树高8.0 m,其中高阶VI 10—12 m的林木所占比例最大。
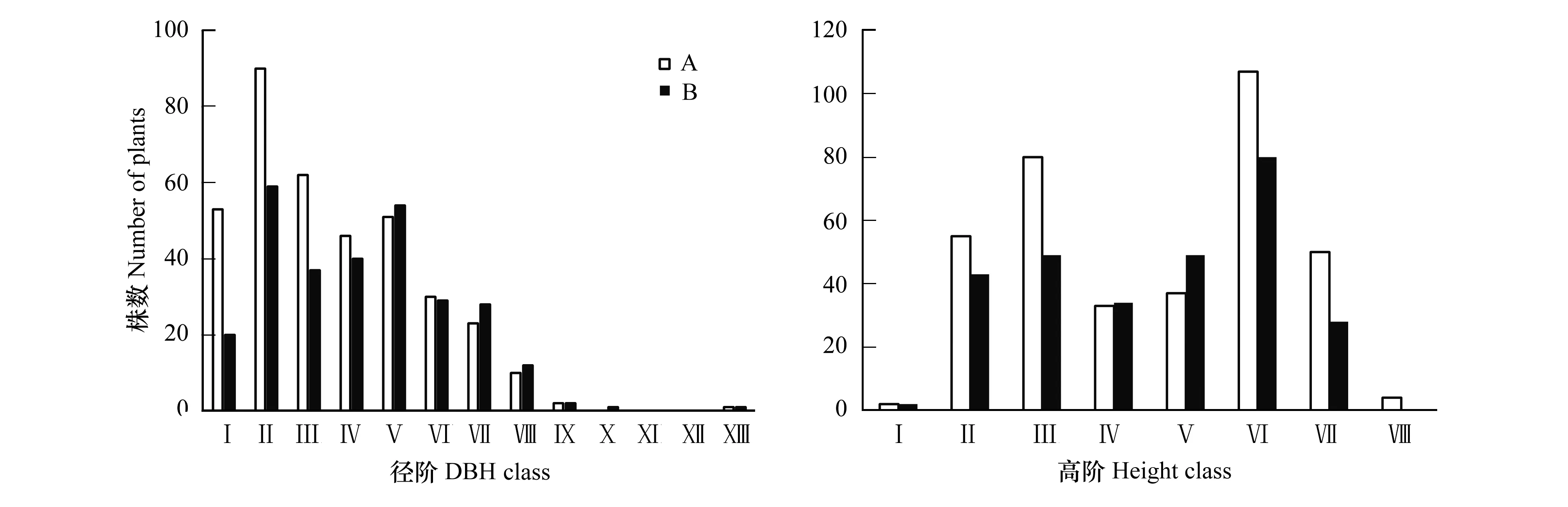
图2 对象木胸径与树高分布特征Fig.2 The distribution features of object trees′ height and DBHI:DBH<3 cm;II:3 cm≤DBH<6 cm;III:6 cm≤DBH<9 cm;IV:9 cm≤DBH<12 cm;V:12 cm≤DBH<15 cm;VI:15 cm≤DBH<18 cm;VII:18 cm≤DBH<21 cm;VIII:21 cm≤DBH<24 cm;IX:24 cm≤DBH<27 cm;X:27 cm≤DBH<30 cm;XI:30 cm≤DBH<33 cm;XII:33 cm≤DBH<36 cm;XIII:36 cm≤DBH<39 cm
3.2 竞争强度
3.2.1竞争范围的选定
如图3所示,随着竞争林木之间距离的增加,林木的平均竞争指数逐渐减小。对竞争指数变化量与林木距离关系研究发现,随着林木之间距离的增加,竞争指数的变化量逐渐减小,在8 m半径处有一个明显的拐点,在8 m半径以内,竞争指数变化量减小非常明显(如样地A 2 m处平均为1.234,4 m处平均为0.644,6 m处平均为0.435;样地B 2 m处平均为1.197,4 m处平均为0.698,6 m处平均为0.471),而在8—14 m半径范围内,半径每增加2 m,竞争指数变化量约减少0.01的幅度,而14 m之外竞争指数变化量基本保持稳定,竞争木对对象木的影响非常微弱。这说明,竞争指数的变化明显与林木之间的距离有关,即竞争木距离对象木越近,对象木受到的影响越大。基于上述分析,本研究认为,对于天然沙地樟子松地理分布最南缘区的中幼林来说,8 m是研究确定其林木竞争的最适宜竞争半径范围,其竞争强度能够较好地反映沙地樟子松种内竞争的真实情况。

图3 样圆半径与竞争指数变化的关系Fig.3 Relationship between radius of the round quadrats and the variation of competition intensity
3.2.2樟子松种内竞争强度
随林木径阶的增大,其竞争指数逐渐减小。由表2可见,A样地中,I径阶的竞争指数最大,为120.31,显著高于其他径阶(P<0.05),IX径阶的竞争指数最低,为5.20,显著低于其他径阶(P<0.05),而IV、V、VI、VII径阶之间无显著差异(P>0.05)。B样地中I径阶的竞争指数最大,为113.65,显著高于其他径阶(P<0.05),IX径阶的竞争指数最低,为5.20,显著低于其他径阶(P<0.05),而IV、V径阶之间以及VII、VIII、IX径阶之间的无显著差异(P>0.05)。以上结果表明径阶小的樟子松受到种内竞争强度较大,径阶较大的受到的种内竞争强度较小。
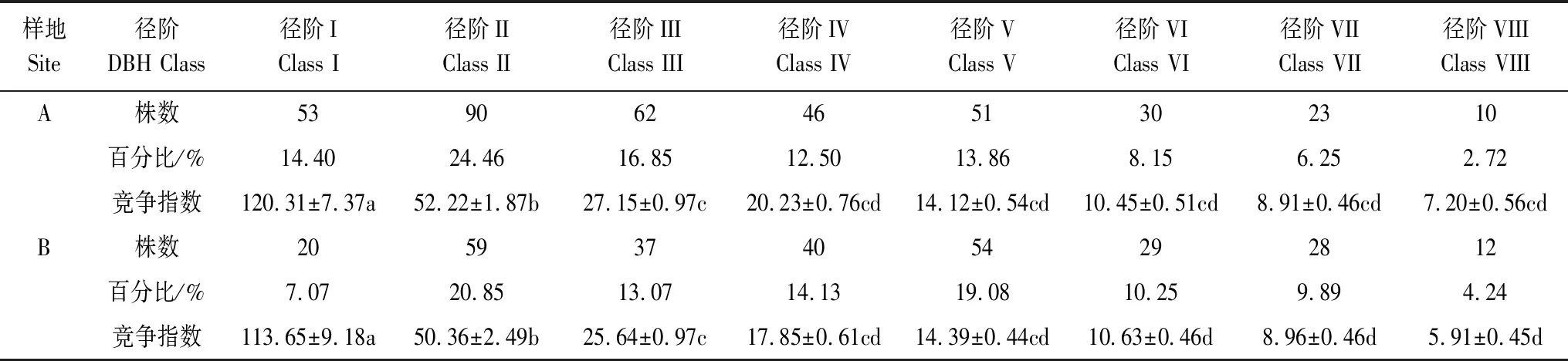
表2 沙地樟子松林分种内竞争强度
不同字母表示不同处理间在5%水平差异显著(P<0.05);径阶IX、X、XI、XII、XIII的株数太少,缺乏统计意义,因此该处未统计分析
3.2.3竞争强度与胸径的关系
A样地的关系为:CI=235.16D-1.10(R2=0.91,P<0.001)(图4);B样地为CI=254.41D-1.15(R2=0.90,P<0.001)(图4);所有林木中对象木胸径与竞争指数的关系为CI=242.24D-1.12(R2=0.91,P<0.001),两个样地均达到极显著水平,表明竞争强度与对象木胸径的关系适合用该模型模拟和预测。由图4可知,随着对象木胸径的增大,个体数逐渐减少,且对象木胸径越大,其所受到的竞争压力越小,而对其周边的个体却可造成极大的竞争压力。当对象木胸径达到20 cm 以上,竞争强度趋于稳定,维持在较低水平。

图4 对象木竞争指数与其胸径的散点图Fig.4 Scatter of competition index of object trees and its DBH空心圆代表樟子松,虚线代表幂函数拟合曲线
3.3 樟子松空间格局分析
3.3.1不同生长阶段林木的空间格局
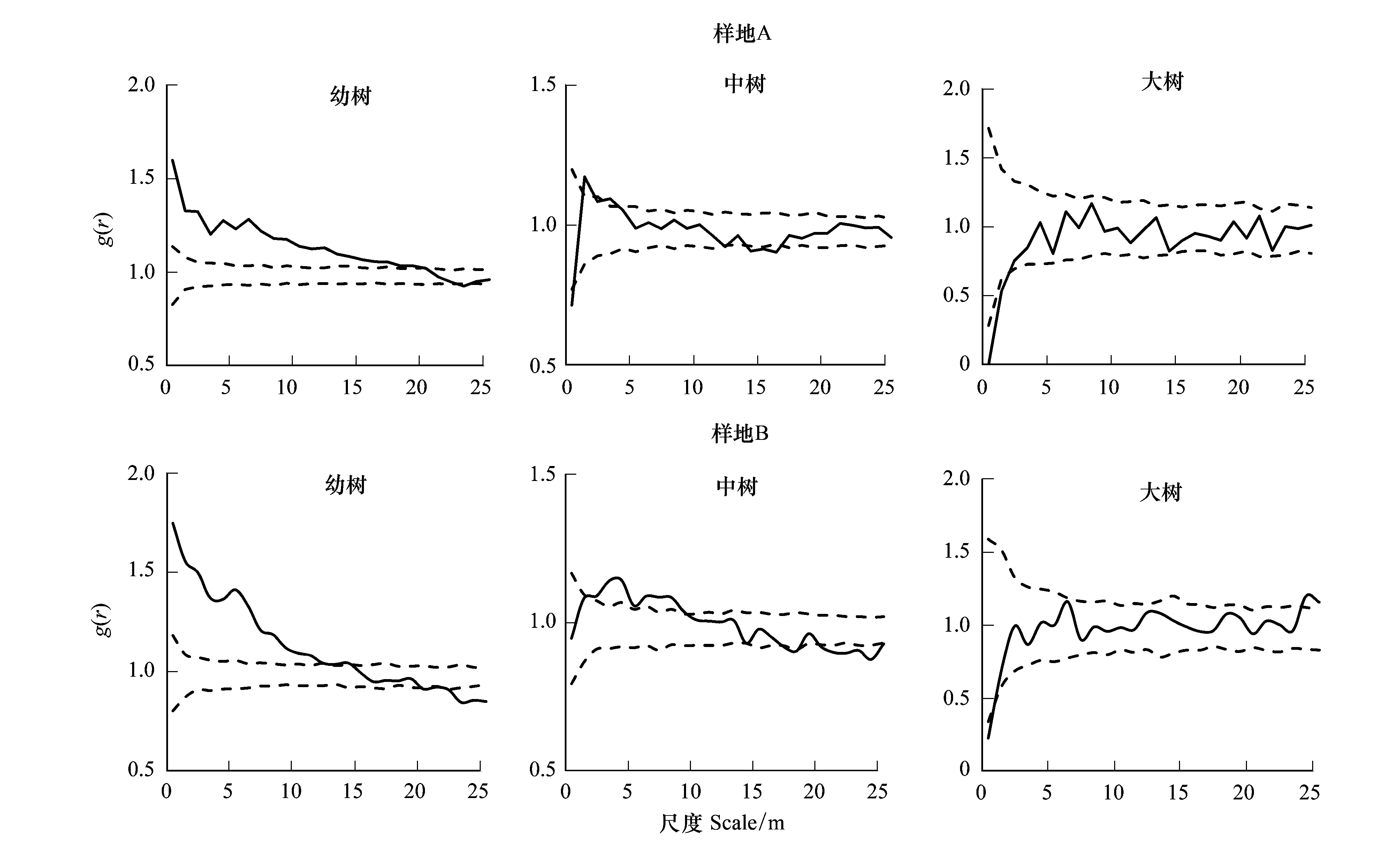
图5 不同生长阶段树木的空间格局Fig.5 Spatial patterns of Pinus sylvestris var. mongolica among different life stages in sampling sites
如图5所示,对两个样地的樟子松种群进行g(r)单变量分析。A样地中,幼树在0—20 m尺度上为显著聚集分布,随着尺度的增加,基本上为随机分布;中树在0—1 m和13.5—16.5 m尺度上为均匀分布,在1—4 m尺度上为聚集分布,在4—14 m和16.5—25 m尺度上为随机分布;大树在0—1.5 m尺度上为显著均匀分布,其他尺度上为随机分布。
B样地中,幼树在0—14.5 m尺度上为显著聚集分布,随着尺度的增加转变为随机分布和均匀分布;中树在2.5—9.5 m尺度上为显著聚集分布,在20—25 m尺度上为均匀分布,其他尺度上为随机分布;大树在0—1.5 m尺度上为显著均匀分布,其他尺度上为随机分布。
3.3.2不同生长阶段林木的空间关联分析
如图6,对樟子松种群的3个生长阶段的空间关联性进行双变量g12(r)分析,结果表明,A样地中幼树与中树在2.5—7.5 m尺度上呈显著正相关,B样地在0—3.5 m尺度上呈显著正相关,其他尺度上两个样地均无相关性;两个样地的幼树和大树呈相互独立关系,无空间相关性;A样地的中树和大树在0—1.5 m尺度上呈显著负相关,而B样地在0—2.5 m尺度上呈显著负相关,其他尺度上无空间相关性。
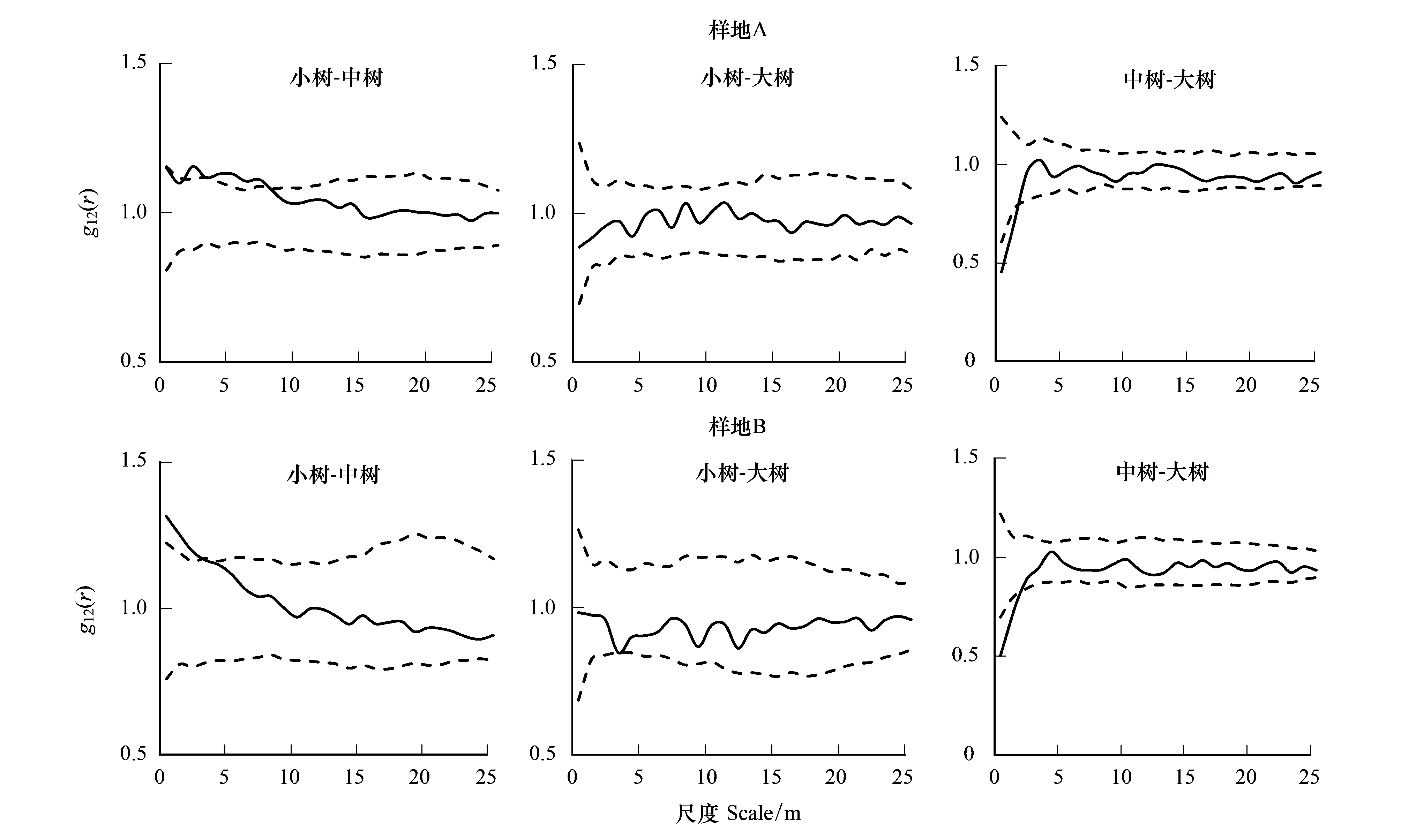
图6 不同生长阶段树木之间的空间关联性Fig.6 Spatial association of Pinus sylvestris var. mongolica among different life stages in sampling sites
3.4 幼树更新
A样地内更新幼树占样地林木总数量的50.51%,B样地的占43.07%;分别统计对象木的东、南、西、北方向上0—2、2—4、4—6 m和6—8 m样圆半径范围内的更新苗数量,结果如图7所示,在对象木外围0—2 m半径范围内,更新幼树的密度最大,在此范围内A样地的平均更新幼树密度为0.09株/m2, B样地的平均更新幼树密度为0.08株/m2;而在A和B样地分别在6—8 m和4—6 m范围内的更新幼树密度最低,A平均更新幼树密度分别为0.03株/m2和0.01株/m2。对不同的方向更新幼树统计发现,对象木冠层南侧的密度最大,平均达到0.43株/m2,冠层西侧的密度次之,平均为0.42株/m2,冠层北侧更新幼树密度最低,为0.39株/m2。然而在A样地6—8 m范围内,E方向与S和W方向差异显著(P<0.05),其他范围内4个方向都无显著从差异(图7)。
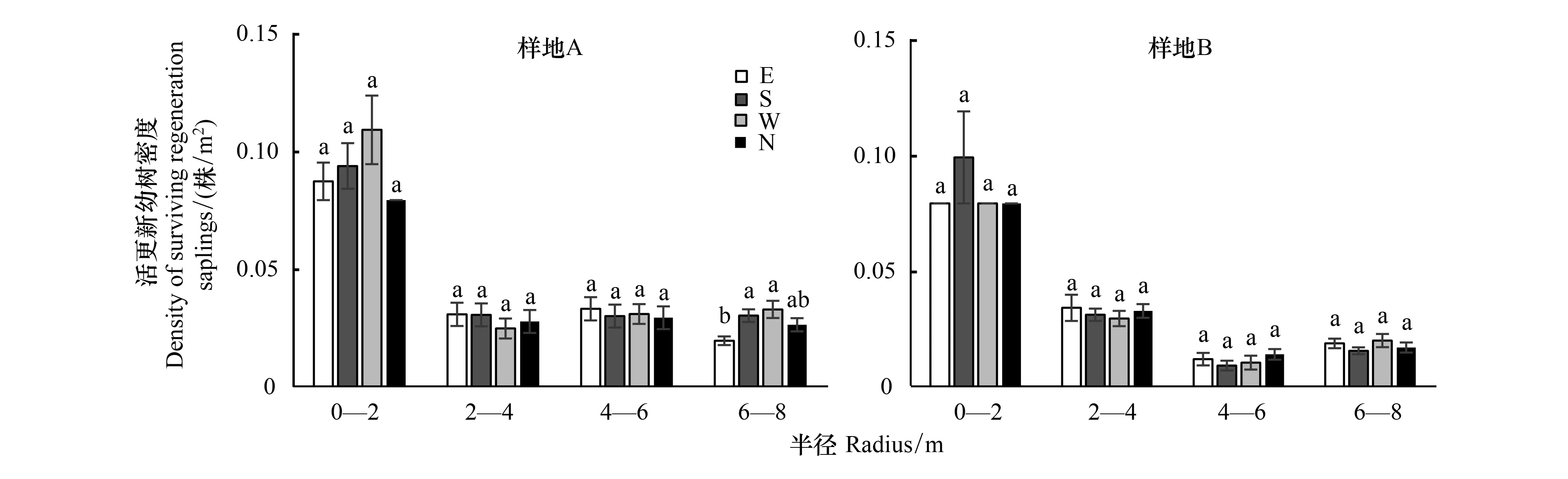
图7 对象木不同方位和半径的更新幼树密度Fig.7 The saplings′ density of different radii in 4 orientations of object trees不同字母表示不同处理间在5%水平差异显著(P<0.05);E:正东方向,East direction;S:正南方向,South direction;W:正西方向,West direction;N:正北方向,North direction
对A和B样地内更新幼树密度与竞争指数进行相关分析,结果如表3,竞争指数与所有更新幼树密度呈极显著正相关(P<0.01),与存活更新幼树密度基本呈正相关。表明随着竞争指数增加,不同样圆半径范围内的更新幼树的密度也随之增加。

表3 更新幼树密度与竞争指数的相关性
**P<0.01,*P<0.05
4 讨论
4.1 种内竞争
在林分内部,林木之间的竞争是经常存在的,由于林分内林木个体的空间生态位大多处于重叠状况,其个体之间必然为其生长而竞争利用有限的资源如光照、土壤水分和养分等,但林木个体之间的竞争是有一定范围的,主要发生在相邻或相近的个体之间[34]。为了确定林木竞争的范围,目前的方法主要有固定半径法[45]、树冠重叠法[46]、角规计数抽样方法[34]和圆锥调查[47]等方法,但应用较多的是简便易算的固定样圆半径法。基于此方法,张学龙等[48]发现,祁连山青海云杉种群的种内竞争主要发生在6 m范围内,黄山松的竞争范围主要集中在7.5 m半径以内[49],而在浑善达克沙地天然沙地榆种群,其竞争主要发生在8 m半径范围以内[50]。
对于天然沙地樟子松地理分布南缘的纯林来说,由于林分密度较大,个体之间的竞争是非常明显的,本研究通过固定样圆法研究发现,当样圆半径介于2—8 m时,其竞争指数持续下降,当超过8 m时,其竞争指数保持在一个较稳定的幅度内,因此,本研究认为8 m是沙地樟子松竞争估算最适宜的范围,这与毛磊等[51]的研究有所差异,他们认为10 m是最适宜的竞争估算样圆半径。这可能与两个研究地点樟子松树木大小不同有关。在红花尔基自然保护区,其天然樟子松较大,林木冠幅半径多介于3—5 m,胸径介于18.5—22.9 cm[51],而本研究中的樟子松树冠半径多介于2—2.5 m,平均胸径9.7—10.5 cm。
林木的竞争受到多种因素的影响,其中包括内在因素(如个体大小、所处生长发育阶段、个体生活力)和外界因素(如所处地形、气候、相邻植株个体大小等)[52]。Hegyi单木竞争指数是测度林木个体间竞争强度较常用的指标[19,32,53-54],它能间接反映出林木个体对光照、水分和营养等资源的现实分配。树木的大小对其竞争强弱有着较大的影响,本研究拟合了胸径与其竞争指数的关系,发现竞争强度与对象木胸径服从幂函数关系, 这与其他一些研究的结果基本一致[19,55]。此外,本研究也发现,当树木胸径低于20 cm时,竞争指数变化显著,而当胸径大于20 cm时,其竞争强度变化很小,基本趋于稳定。
4.2 空间格局
许多研究已经发现自然生态系统内的幼年个体大多呈聚集分布的特征,然而,随着个体逐年长大,生态系统内的个体可能会逐渐转向随机或均匀分布的状态[7]。在本研究的中幼林中,幼树阶段的樟子松种群大约在小于15—20 m的尺度上呈聚集分布状态,在此阶段,由于个体较小,其对资源与空间的需求较小,通过聚集分布有利于共同抵抗不良环境的影响,以抵抗外界干扰来维持种群自身稳定性,从而促进种群的繁衍[56-57];而随着树木胸径的增加,个体对资源和空间的需求越来越大,在有限的资源环境约束下,生活力弱的个体由于激烈的竞争而无法获取水分与营养等资源,种群开始出现自疏现象,在这一作用下,个体之间的相互作用变弱,种群的分布趋向于均匀或随机分布[58]。而在1—4 m尺度上中树呈聚集分布特征,这可能是由于生境异质性效应(土壤养分、洼地等)导致的[59]。
许多生态过程会导致物种的聚集分布特征,这是空间格局尺度依赖性的重要理论基础[60]。许多研究表明,种子传播限制和生境异质性(如土壤养分与水分、地形差异等)是影响物种聚集分布的重要因子[61-63]。本研究中,种子繁殖是樟子松种群的主要繁殖方式[64-65],种子成熟后会在重力和风等因素的共同作用下落到地面,以母树为中心形成聚集分布状态,而随着与母树距离的增加樟子松的种子密度逐渐降低[64-65],或者在风力、地面径流、地面植物的阻截等外力作用下形成聚集状态。因此,种子传播限制是影响樟子松幼树聚集分布格局的重要因子。此外,生境异质性也是樟子松幼树聚集分布的重要影响因素之一,在沙丘的作用下,土壤、地形、光照等资源呈现出空间上的斑块状分布,资源良好的生境为种子的萌发和定植创造了有力条件,从而导致物种的聚集分布[66]。
在中或较大尺度上,幼树、中树和大树之间彼此都表现相互独立的关系,但幼树和中树在较小尺度上呈正相关,这可能是由于中树为其种子的萌发和幼树的生长提供适宜的微环境,形成了一种相互有利的空间关系[67-68],即植物个体间的“庇护与被庇护”效应[69]。由于具备更多的可用水量、更高的土壤养分以及良好的遮阴环境,“庇护个体”以其固定的树冠可以持续地为种子的萌发、幼苗的存活提供良好的环境[70]。幼树与大树在所有尺度上无相关性,这或许是由于大树对幼树造成极大的竞争压力,使得幼树无法获取生存所需资源,最后导致大树周围的幼树存活率降低[71],从而主要分布于竞争压力适宜的中树周围;中树和大树在小尺度上呈负相关,这可能与两者的营养生态位发生重叠有关,中树在获取资源及生存空间时会受到大树的抑制,中树的平均冠幅达1.8 m,而大树的平均冠幅约3.0 m,且两者的冠层在小尺度上发生较大的重叠效应,这支持了本研究的结果,而在较大尺度上,两者之间的竞争效应减弱,从而表现出相互独立的关系。
4.3 种群更新
种群更新是一个极为重要的生态学过程,对群落演替与动态变化具有重要的影响,更新过程涉及到种子扩散、土壤种子库、种子萌发与生长、幼树竞争等方面[72-74]。本研究发现樟子松幼树更新多分布于树冠的南侧和东侧,这与闫德仁等[73-74]的研究结果较为一致,他们在进行土壤种子库研究时发现,树冠南侧土壤种子库的种子数量最多,且随距树干距离(2—6 m)增加而增加,但距树干8 m后降低。另外,本研究也发现更新幼树密度最大的区域多位于树木外围的0—2 m半径范围内,即树冠下,这与闫德仁等[73-74]的研究稍有不同,他们认为樟子松更新苗主要集中在母树树冠的边缘和距离母树10 m范围内,这可能与闫德仁等[73-74]研究的树木冠幅较大有关。
种群更新受许多生物因素如母树大小、密度[73]和环境因素如气候[73-74]、火干扰[19,24- 25]等的影响。本研究分析了母树竞争与幼树更新的关系,发现随着竞争指数增加,不同样圆半径范围内的更新幼树的密度也随之增加。即竞争指数越高,胸径越小,个体的竞争能力就越弱,其受周围个体的竞争压力就越强,而对周围更新幼树产生的竞争压力就会越小,因此更新幼树趋向于分布在竞争指数高的个体周围。
本研究发现更新幼树呈聚集分布的空间格局,在小尺度上,更新幼树与中树呈正相关,而在较大尺度上,与中、大树之间均呈相互独立的关系,这与尤海舟等[71]的结果相似。即两个径阶之间差异越小,其个体之间的正相关就越强,个体之间更趋向于协调[71]。而母树竞争强弱又会影响着幼树的更新与格局,即随着竞争指数的增加,更新幼树的密度会逐渐增加。
在本研究区的沙地樟子松纯林中,林分受外界干扰较少,天然更新良好,种群处于增长状态,但由于林分密度较大,个体受竞争的影响非常大,推测未来竞争可能导致生活力弱的个体死亡率增加,自疏作用增强。在竞争压力下,林分的空间格局可能发生改变,导致形成许多林隙,从而使樟子松的幼树多以集群状态分布在林隙边缘竞争较弱的个体周围,从而加速本区沙地樟子松种群的扩张。
5 结论
本研究采用Hegyi竞争指数与点格局等分析方法,分析了呼伦贝尔沙地东南端沙地樟子松天然林沙地樟子松的径阶和树高结构、种内竞争、不同生长阶段空间格局及空间关联性、以及幼树更新特征,主要结论如下:1) 在地理分布南部边缘区的沙地樟子松林处于中幼龄阶段,属增长型种群;2) 沙地樟子松对象木胸径与竞争指数服从幂函数关系,随着林木胸径的增加,其竞争指数逐渐减小(即竞争强度越强);3) 林木幼树在小尺度上呈聚集分布特征,而随着胸径的增加,在自疏等作用影响下,个体之间相互作用变弱,促使中树与大树在中大尺度呈随机分布;幼树与中树在“庇护与被庇护”效应的作用下导致其在小尺度呈正相关,而中树与大树由于营养生态位的重叠,导致两者在小尺度呈负相关;4) 竞争指数与更新幼树和存活更新幼树的密度均呈显著正相关(P<0.05),即更新幼树趋向于分布在竞争较弱的林木个体周围。未来在沙地樟子松的经营管理中,需要充分考虑林木竞争、空间格局以及种群更新的关系,统筹规划,采取合理的培育措施,以促进沙地樟子松种群的天然更新。
猜你喜欢
林业科学研究(2021年6期)2022-01-05
内蒙古林业调查设计(2021年5期)2022-01-05
林业科技情报(2021年2期)2021-07-13
河北果树(2020年1期)2020-02-09
农民致富之友(2019年8期)2019-05-22
现代园艺(2018年2期)2018-03-15
现代园艺(2018年2期)2018-03-15
北极光(2018年12期)2018-03-07
现代园艺(2017年11期)2017-06-28
土壤与作物(2015年3期)2015-12-08
