
闽江口互花米草淤积作用对其自身和短叶茳芏残体分解及硫养分释放的影响
2019-07-05 09:01孙志高
生态学报 2019年10期
王 杰,孙志高,3,*,何 涛,高 会,王 华,李 晓
1 福建师范大学地理研究所, 福州 350007 2 福建师范大学湿润亚热带生态地理过程教育部重点实验室, 福州 350007 3 福建师范大学福建省亚热带资源与环境重点实验室, 福州 350007
湿地残体分解是营养物质循环和能量流动的关键环节,它连接着生物有机体的合成与分解,是维持湿地生态功能的主要过程之一[1- 2]。残体分解速率的高低在很大程度上影响着残体的现存量以及氮、磷、硫等养分和其他物质向沉积物中的归还,并影响着湿地的物质循环状况[3]。河口湿地残体分解受到诸多因素(水分、潮汐、盐度、土壤动物、促淤强度等)的影响[4- 7]。在诸多因素中,促淤是影响残体分解的一个极为重要的因素[8- 10],而植物拦沙淤积对其自身及其他植被残体分解过程的影响可能更为重要,因为河口潮滩植物可通过絮凝作用、消浪缓流作用、改变水体紊动和黏附悬沙运动等复杂过程来促进河流与潮汐以及风暴潮裹挟的泥沙在潮滩快速淤积下来[11- 16]。在这一系列复杂过程中,植物自身以及其他残体可被掩埋到不同深度的沉积物中,从而可能显著影响到河口潮滩湿地自身的物质循环和能量流动。然而,目前关于河口湿地植物促淤作用对其自身及其他植被残体分解影响的研究还鲜有报道。
目前,国外已在河口湿地植物残体分解方面开展了较多研究,且研究内容已涉及残体分解特征、影响因素(水分、盐度、温度、养分和淤积强度等)[4- 7]和元素释放(碳、氮、磷及痕量元素)规律[17- 21]等方面。与之相比,国内在该方面也开展了许多工作,且研究区域多集中于黄河口湿地[19-21]、长江口湿地[22]、闽江口湿地[23- 26]以及南方主要河口的红树林沼泽[7]等。整体而言,当前国内关于河口湿地植物残体分解的研究大多集中在分解规律、养分释放(碳、氮和磷)及一般影响因素(如温度、水分和盐度)的探讨上,而关于河口湿地淤积对残体分解的研究仅集中于黄河口湿地和长江口湿地[8,22],但关于植被拦沙淤积作用对其自身以及其他植被残体分解和养分(特别是硫)释放的影响研究还不多见。
闽江河口位于北亚热带与南亚热带的过渡区,是中国东南部最典型的亚热带河口。鳝鱼滩湿地是闽江河口分布面积最大的一块湿地,其主要植被类型包括芦苇(Phragmitesaustralis)、短叶茳芏(Cyperusmalaccensis)和扁穗莎草(Cyperuscompressus)。2002年以来,互花米草开始入侵鳝鱼滩,至2010年其分布面积已达306.9 hm2,年均增加9.82%。近年来,互花米草开始入侵鳝鱼滩湿地中西部,且主要沿潮沟两侧逐渐扩大其覆盖范围[27]。在沿潮沟向陆一侧扩展过程中,其与本地种短叶茳芏形成了较为明显的交错带。互花米草作为一种耐盐、耐淹的禾本科米草属多年生草本植物,植株粗壮高大(1 m以上)且密度高,加之根系发达,故其存在可显著增加潮滩的表面粗率,阻滞潮流和波浪,具有明显的促淤作用[11- 16]。当前,关于闽江河口湿地植物残体分解的相关研究主要集中在本地植被(芦苇和短叶茳芏)和入侵植被(互花米草)残体分解速率、养分释放(碳、氮和磷)及一般影响因素(如水分、盐度和植物类型)的探讨上[23- 26],而关于互花米草的促淤作用对其自身以及其他植被(如短叶茳芏、扁穗莎草)残体分解和硫释放的影响研究还鲜有报道。鉴于此,本研究以鳝鱼滩互花米草入侵短叶茳芏过程中形成的交错带湿地为研究样地,基于原位淤积模拟试验,探讨了不同淤积强度对互花米草和短叶茳芏残体分解及硫养分释放的影响。研究结果有助于明晰互花米草入侵过程中的养分循环特征,并可为该区湿地的生态保育提供重要依据。
1 研究区概况与研究方法
1.1 研究区概况
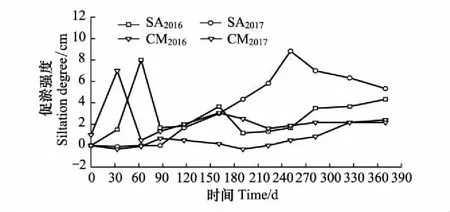
图1 2016—2017年互花米草与短叶茳芏的促淤动态 Fig.1 Dynamics of siltation by Spartina alterniflora and Cyperus malaccensis during 2016—2017SA2016: 2016年交错带淤积强度变化, Change of siltation degree in ectone during 2016; SA2017: 2017年交错带淤积强度变化, Change of siltation degree in ectone during 2017; CM2016: 2016年短叶茳芏群落淤积强度变化, Change of siltation degree in Cyperus malaccensis during 2016; CM2017: 2017年互花米草群落淤积强度变化, Change of siltation degree in ectone during 2017
闽江河口湿地地处中亚热带和南亚热带海洋性季风气候过渡区,气候暖热潮湿,雨热同期,年平均气温19℃,降水量可达1382 mm。闽江河口潮汐属正规半日潮,潮汐作用强烈,湿地土壤类型属滨海盐土,天然植被主要有芦苇(Phragmitesaustralis)、短叶茳芏、扁穗莎草(Cyperuscompressus)和互花米草等。自2002年互花米草入侵鳝鱼滩以来,入侵方向大致可分为海向和陆向。海向主要是向滩涂入侵,而陆向主要是向本地种如短叶茳芏和扁穗莎草分布区入侵。近年来,在鳝鱼滩西北部已形成了大片互花米草与短叶茳芏的交错带群落。目前,国内外已运用不同方法或技术(高程重复测量、210Pb、137Cs测年和P-A拟合方法、遥感技术等)对不同区域(路易斯安娜河口[11]、江苏新洋港[12]、江苏中部海岸潮滩[13- 14])的互花米草促淤速率进行了大量研究,发现其促淤速率大致为3.5—6.5 cm/a。本研究组亦对闽江河口鳝鱼滩的短叶茳芏和互花米草的促淤速率进行了连续两年的研究(图1),具体做法为:首先,在研究样地设置2列标杆,每列4根,间隔为6 m。标杆长度为3 m,其中2 m被垂直打入土壤,以确保其在外力扰动下保持垂直且不发生偏移;其次,在每根标杆距离地表0 cm和10 cm处做标记。若发生地表侵蚀,则侵蚀强度按地面至0 cm处标记的高度进行测量;若发生地表淤积,则淤积强度按照地面至10 cm处标记的高度进行换算。通过每月测定研究样地的侵蚀/淤积强度,以分析其淤积的动态变化,进而可测算出其年淤积强度。据图可知,互花米草的促淤强度在不同时期差异较大,但其年促淤速率较为接近,约为4.33—5.33 cm/a,与上述相关研究结果接近。与之相比,互花米草入侵前短叶茳芏的促淤强度较低,其年促淤速率约为2.17—2.40 cm/a(图1)。
1.2 样地布设与样品采集
2016年2月,基于野外原位分解袋法,在闽江河口鳝鱼滩西北部互花米草(SA)与短叶茳芏(CM)的典型交错带(互花米草入侵初期,1—2 a)同时采集两种植物的残体。将残体带回实验室用去离子水冲洗烘干后剪成 10 cm 左右小段,装入孔径为 0.2 mm,规格为 20 cm×25 cm的分解袋中,每袋15 g(干重)。野外原位淤积模拟试验参照上述国内外研究数据以及本研究组的实际监测数据取平均值将互花米草当前的淤积强度设定为5 cm/a。2016 年2月19日,在互花米草与短叶茳芏的典型交错带设定无淤积(S0,0 cm/a)、当前淤积(S5,5 cm/a)和未来淤积增强(S10,10 cm/a)3种处理,以模拟互花米草入侵过程中导致的淤积作用(或未来淤积增强)对其自身以及短叶茳芏残体分解的影响。在研究样地内,每个处理随机设定3个分解小区。为防止研究期间分解袋受到其他促淤干扰的影响,采用尼龙网(孔径0.2 mm,高1.5 m)将各分解小区围起。本研究所用互花米草与短叶茳芏残体的基本性质如表1所示。
1.3 样品采集与分析
试验期内(0—375 d),不定期从分解小区取回分解袋,每次3—4个重复,共采集13次(15、30、62、94、123、153、184、217、245、276、306、334 d和375 d)。将取回的分解袋及时带回实验室,去掉泥土并拣出袋内杂物,用去离子水将样品清洗干净后,置于烘干箱中烘干至恒重。称量后,将样品用粉碎机磨碎,过100目筛后装袋待测。采用元素分析仪(Vario EL,Germany)测定样品的全碳(TC)和全氮(TN)含量;利用HNO3-HClO4消解,BaSO4比浊法测定样品的全硫(TS)含量。试验进行时,同步测定地表温度、土壤含水量、pH和电导率(EC)等环境因子。
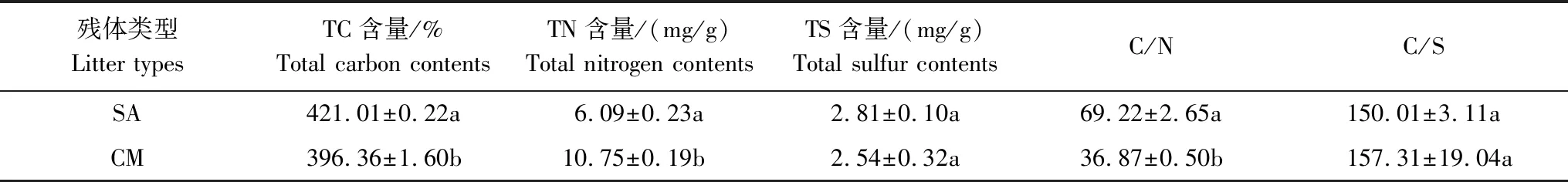
表1 两种植物残体的基本性质
同列不同字母表示数据间在p<0.05水平上差异显著; SA: 互花米草, Spartina alterniflora; CM: 短叶茳芏, Cyperus malaccensis
1.4 指标计算
残体的残留率(R, %)和分解速率(d-1)可用下式计算:
R= (Wt/Wo)×100%
ln (Wt/Wo) = -kt
式中,Wt为分解t时间后残体的残留量(g),Wo为残体的初始质量(g),k为分解速率常数(d-1),t为分解时间(d),R为残体的残留率(%)。
残体分解过程中S的累积或释放可用累积指数(SAI)表示,即
SAI=MiXi/MoXo× 100%,
式中,Mi为残体在t时刻的质量(g),Mo为残体的初始质量(g),Xi为t时刻残体的S含量(mg/g),Xo为残体中的S初始含量(mg/g)。若SAI <100%,表明残体在分解过程中发生S净释放,反之则发生S净累积。
1.5 数据处理与分析
运用Origin 8.0软件对数据进行计算和作图,采用SPSS 20.0软件对相同植物残体在3种促淤强度间以及相同促淤强度下不同植物间的残留率、TS、SAI以及环境因子(地温、含水量、pH、EC)等数据进行单因素方差分析,对残体的残留率、TS含量、SAI、C/N、C/S和环境因子数据进行Pearson相关分析和逐步线性回归分析。
2 结果与分析
2.1 淤积对残体残留率的影响
不同淤积强度下互花米草与短叶茳芏残体的残留率在研究时段内均呈波动下降趋势,且其值均随着淤积强度的增加而增加(图2)。互花米草残体的残留率在经历375 d分解后分别为14.33%(S0)、32.57%(S5)和31.67%(S10),且S0处理与S5(或S10)处理之间存在显著差异(P<0.05)。 与之相比,短叶茳芏残体在S0、S5和S10处理下的残留率分别为6.03%、12.73%和16.13%,但三者之间的差异均不显著(P>0.05)。比较而言,相同淤积强度下短叶茳芏的残留率均小于互花米草,且二者在S5或S10处理下的残留率之间还存在极显著差异(P<0.01)。上述结果表明,互花米草入侵导致的淤积作用对两种植物残体的分解均产生重要影响,且这种影响在短叶茳芏残体分解过程中表现的更为显著。
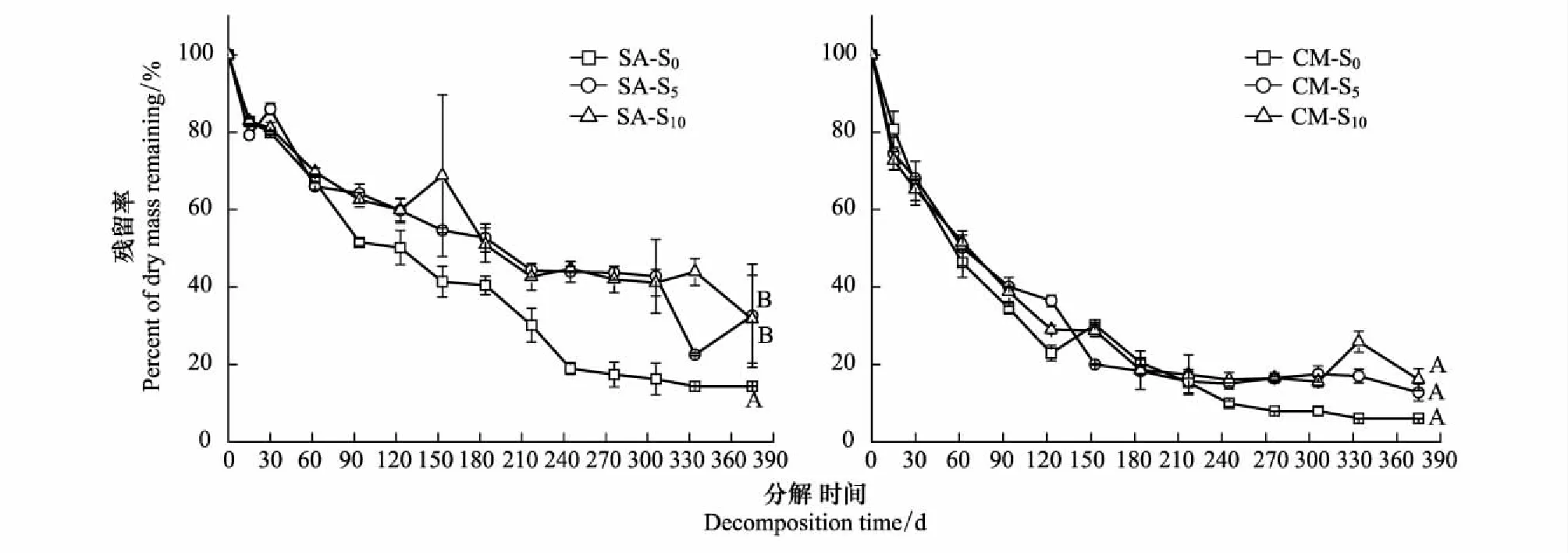
图2 不同淤积强度下残体残留率的变化Fig.2 Variations of dry mass remaining of litters in different siltation treatmentsSA-S0/ CM-S0: 无淤积处理下的互花米草/短叶茳芏; SAS5/ CM-S5: 当前淤积处理下的互花米草; SA-S10/ CM-S10: 未来淤积增强下的互花米草;不同的大写字母表示同种植物不同淤积强度下存在显著差异(P<0.05)
2.2 淤积对残体分解速率的影响
不同淤积强度下互花米草和短叶茳芏的分解速率均存在明显差异(表2)。其中,S0处理下二者残体的分解速率最高(SAS0=0.0055 d-1和CMS0=0.0074 d-1),S5处理次之(SAS5=0.0028 d-1和CMS5=0.0048 d-1),S10处理最低(SAS10=0.0024 d-1和CMS10=0.0041 d-1)。相应的,二者 95%分解所需时间(t0.95)分别为SA=1.36 a与CM=1.01 a(S0)、SA=2.70 a与CM=1.38 a(S5)以及SA=3.18 a与CM=1.54 a(S10)。与S0相比,S5与S10处理下二者的分解速率分别降低SA=49.09%、CM=35.14%(S5)和SA=56.36%、CM=44.59%(S10),t0.95分别增加SA=98.53%、CM=36.63%(S5)和SA=133.82%、CM=52.48%(S10)。比较而言,相同淤积强度下,短叶茳芏(SA)残体的分解速率分别为互花米草(CM)残体的1.35倍(S0)、1.71倍(S5)和1.70倍(S10),t0.95分别缩短0.35 a、1.32 a和1.64 a。上述结果表明,随着互花米草入侵导致的淤积强度的增加,两种植物残体的分解速率均明显降低,但其对短叶茳芏残体分解速率的影响最为明显。
表2 不同淤积强度下残体残留率自然对数(y)与分解天数(x)线性拟合方程及其相应参数
Table 2 Linear equations and parameters between the natural logarithm(y)of mass remaining and decomposition days(x)in different siltation treatments

残体类型Litter types淤积强度Siltation treatments线性回归方程Linear equationsk/d -1R2Pt0.95/aSAS0y= -0.0055x-0.08190.00550.97< 0.011.36S5y= -0.0028x-0.17110.00280.85< 0.012.70S10y= -0.0024x-0.17570.00240.90< 0.013.18CMS0y= -0.0074x-0.27620.00740.96< 0.011.01S5y= -0.0048x-0.48210.00480.84< 0.011.38S10y= -0.0041x-0.53290.00410.75< 0.011.54
S0:无淤积强度处理, No siltation treatment; S5:当前淤积强度处理, Current siltation treatment; S10:未来淤积增加处理, Strong siltation treatment
2.3 淤积对残体碳硫含量变化的影响
2.3.1碳含量变化
尽管不同淤积强度下互花米草和短叶茳芏残体中的TC含量变化模式差异较大(图3),但分解期间S5与S10处理下二者的TC含量均明显高于S0处理(P<0.01)。S0处理下两种残体的TC含量变化趋势在0—270 d基本一致,之后则呈相反规律变化。虽然S5和S10处理下相同植物残体的TC含量变化趋势较为一致(P>0.05),但相同淤积强度下不同植物残体TC含量之间的变化特征差异较大(P<0.05)。375 d后,S5处理下互花米草残体的TC含量相对于初始值增加了3.47%,S0和S10处理下残体的TC含量相对于初始值分别降低了9.17%和5.99%。与之不同,不同淤积强度下短叶茳芏残体的TC含量相对于初始值均降低,降幅分别为23.34%(S0)、5.98%(S5)和6.54%(S10)。上述结果表明,随着互花米草入侵导致的淤积强度的增加,两种植物残体在分解过程中的TC含量整体均呈增加趋势,但其对于分解末期短叶茳芏残体中TC含量变化的影响较为明显。
2.3.2硫含量变化
不同淤积强度下互花米草残体中的TS含量均呈波动增加趋势,其值至试验结束相对于初始含量分别增加了31.46%(S0)、32.57%(S5)、13.60%(S10)。与之不同,S0处理下短叶茳芏残体中的TS含量整体呈较小波动变化(CV=20.35%),而S5和S10处理下残体的TS含量变化趋势较为一致且均呈先增加后降低变化。375 d后,短叶茳芏残体的TS含量相对于其初始含量分别增加了28.22%(S0)、165.51%(S5)、124.00%(S10)。比较而言,S0处理下两种植物残体的TS含量在0—120 d均明显高于S5和S10处理(P<0.05);120—375 d,3种淤积强度下互花米草残体的TS含量变化差异不大(P>0.05),而S5和S10处理下短叶茳芏残体的TS含量均高于S0处理(P<0.01)。此外,相同淤积强度下两种植物残体的TS含量在S0处理下并未达到显著差异水平(P>0.05),而在S5和S10处理下则达到极显著差异水平(P<0.01)。上述结果表明,随着互花米草入侵导致的淤积强度的增加,两种植物残体在分解过程中的TS含量整体均呈增加趋势,但其对于短叶茳芏残体中TS含量变化的影响较为明显(图3)。

图3 不同淤积强度下残体TC和TS含量变化Fig.3 Variations of TC and TS contents in litters in different siltation treatments
2.4 淤积对残体硫释放的影响
不同淤积强度下互花米草和短叶茳芏残体的SAI在研究时段内均小于100%,说明二者在分解过程中均发生了硫养分净释放(图4)。分解期间,3种淤积强度下互花米草的SAI分别降低了25.09%—88.51%、34.14%—67.91%和32.42%—71.56%,而短叶茳芏的SAI分别降低了24.12%—92.25%、31.18%—67.71%和28.12%—65.75%,说明相同淤积强度下短叶茳芏残体释放的硫养分整体要高于互花米草。比较而言,S0处理下互花米草和短叶茳芏残体的硫释放量在0—120 d均明显低于S5和S10处理(P<0.05),而在120—375 d,其硫释放量均明显高于S5和S10处理(P<0.05)。上述结果表明,随着互花米草入侵导致的淤积强度增加,两种残体的硫释放量整体均呈降低趋势,但短叶茳芏残体的硫释放量要高于互花米草。
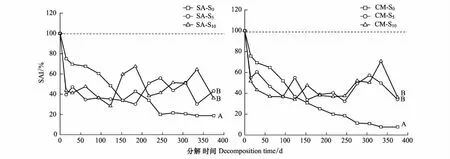
图4 不同淤积强度下残体硫累积指数(SAI)的变化Fig.4 Variations of sulfur accumulation index (SAI) of litters in different siltation treatments
3 讨论
3.1 淤积强度对残体分解速率的影响
本研究表明,不同淤积强度下互花米草与短叶茳芏残体的失重率均随淤积强度的增加而降低,这可能与不同淤积强度下分解样地的环境因子密切相关。相关分析表明,三种淤积强度下,互花米草与短叶茳芏残体的残留率与EC均呈极显著负相关(P<0.01),与pH呈正相关(二者在S10处理下呈显著正相关,P<0.05),与含水量及地温大多呈负相关(P>0.05)(表3)。逐步线性回归分析进一步表明,EC(x)均是不同淤积强度下残体残留率变化的主要影响因素(表4)。本研究中,随着淤积强度的增加,盐度(以EC来表征)越高(表5),两种残体的残留率越高(图2)。尽管前人关于盐度对残体分解的影响尚无定论[6- 7,28- 31],但该结果与多数研究结果相近[7,28- 30],即盐度(以EC表征)增加可抑制残体分解,而这可能与盐度对参与分解微生物的胁迫机制有关[29]。一般而言,盐度会影响微生物的生理类群及酶活性,因为盐分的毒害作用抑制了酶活性和微生物活性,进而导致微生物对残体的降解能力减弱[29]。胡宏友等关于九龙江口秋茄(Kandeliacandel)分解对于盐水梯度响应的研究也得到类似结果,即盐度的升高显著降低了秋茄残体的呼吸速率和分解速率[7]。Roache等的研究也表明,盐分的增加可明显抑制微生物的活性,进而抑制残体的分解[28]。一般而言,随着土壤水分的增加,水体中的氧气浓度下降,而这会对微生物活性产生抑制,进而降低残体的分解速率[9]。另外,在微生物活动的最适宜温度范围内(约为25—35℃)[32],温度升高可通过提高微生物活性而加速残体分解,而温度的降低则会减缓残体的分解[33- 34]。本研究中,尽管两种残体的残留率与含水量(或地温)之间的相关性均不显著,但其在整体上与含水量或地温呈负相关(表3),说明随着淤积强度的增加,含水量和地温对于残体分解均起到了一定的抑制作用,而这点可为表4和图2中的相关结果所证实,即随着深度增加,含水量随之增加,地温随之降低,而残体的残留率增加。尽管残体的残留率与pH呈大多呈正相关(S10处理下呈显著正相关),但其对于不同淤积强度下残体分解的影响可能并不大,原因与不同淤积强度下沉积物的pH变幅较小(介于6.3—6.5之间)且不存在显著差异有关(P>0.05)(表3)。
不同淤积强度下两种残体分解速率的变化不仅与上述分解样地的主要环境因子有关,而且还可能与残体本身的初始基质质量以及淤积作用导致的养分条件改变对残体基质质量的影响有关。由于C/N可反映微生物对养分的需求状况,故常常被认为是反映和预测分解速率的重要指标,C/N越高,残体分解越缓慢,反之越快[2]。本研究中,尽管不同促淤强度下相同残体的初始C/N相同(表1),但在分解期间,互花米草残体的C/N在S0与S5以及S0与S10之间存在极显著差异(P<0.01),且S5与S10处理下残体的C/N均大于S0(图5),说明随着淤积强度的增加互花米草残体的C/N呈增加趋势,而C/N的增加可对残体分解产生一定的抑制作用;与之不同,短叶茳芏残体的C/N在S0、S5和S10处理下较为接近(图5)且不存在显著差异(P>0.05)(表5),即随着淤积强度的增加,残体中的相近C/N是导致3种淤积强度下残体分解不存在显著差异的重要原因。本研究还表明,相同淤积强度下互花米草残体的分解速率均小于短叶茳芏残体(表2),这与互花米草属木质素和纤维素较高的草本植物[35- 36],其残体的C/N均大于短叶茳芏且二者之间存在极显著差异有关(P<0.01)(图5和表5)。另外,两种残体分解速率的变化还可能与淤积作用导致的养分条件改变而对二者基质质量的影响有关。已有研究表明,湿地沉积强度增加可通过许多直接或间接机制来抑制残体分解的进行,如改变残体周围水化学特征、对残体碎屑的物理压实作用、减少残体与周围环境的气体和养分交换、抑制微生物或大型湿地动物活动等[10]。本研究表明,不同淤积强度下相同残体的C/N之间存在一定差异(其中互花米草的S0处理与S5、S10处理均呈显著差异P<0.05(表5)),而该差异与淤积作用导致分解环境中的养分条件及残体的C/N改变有关[21]。由于S0淤积强度下的残体贴近地表且更易从周围水环境和悬浮颗粒中获得养分,由此使得该处理下两种残体的C/N相对于S5和S10淤积强度明显降低(图5),进而导致S0处理下残体的分解较快,而S5和S10淤积强度下较慢。另外,随着淤积强度的增加,沉积物的通气状况变差,好氧微生物的活性降低,进而导致两种残体的分解速率相对于S0处理明显降低。
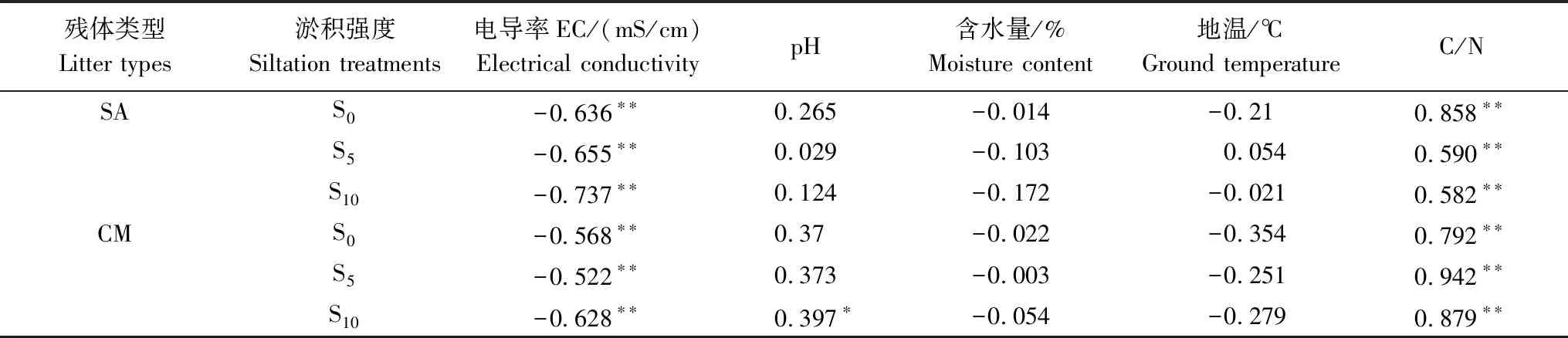
表3 不同淤积强度下残体的残留率与其他因子之间的相关性
*为P<0.05; **为P<0.01
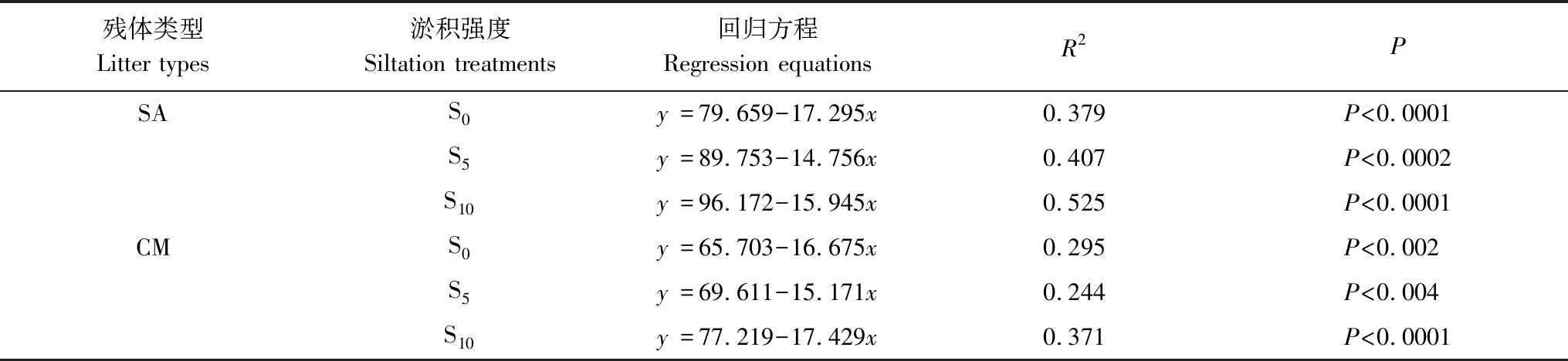
表4 不同淤积强度下残体残留率(y)与主要影响因子(EC, x)回归方程
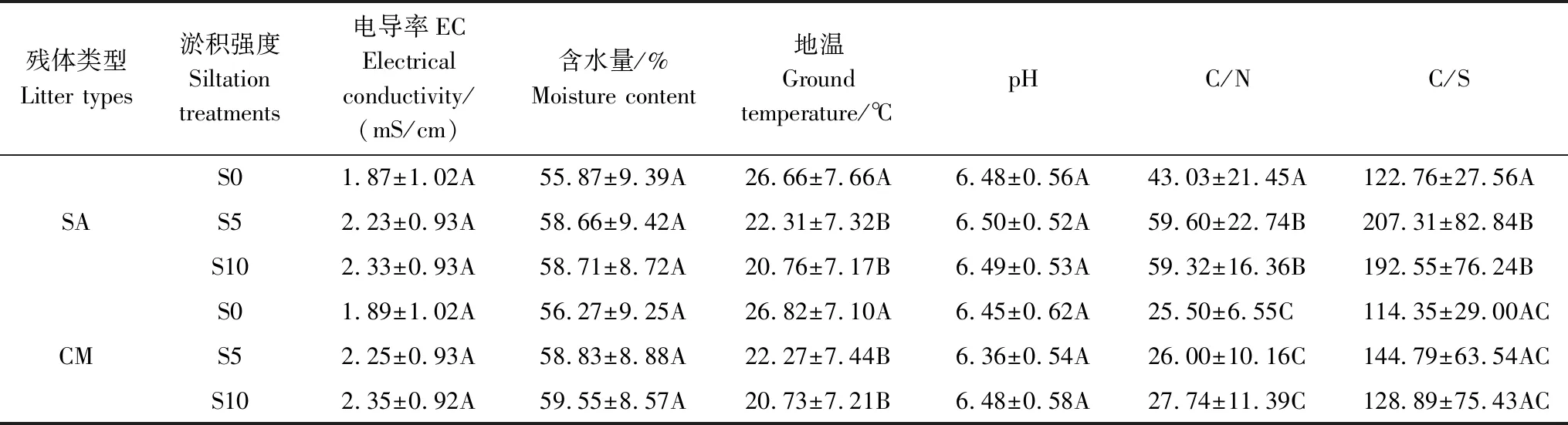
表5 不同淤积强度下的环境因子对比
同列不同字母表示数据间在P< 0.05或P< 0.01水平上差异显著或极显著
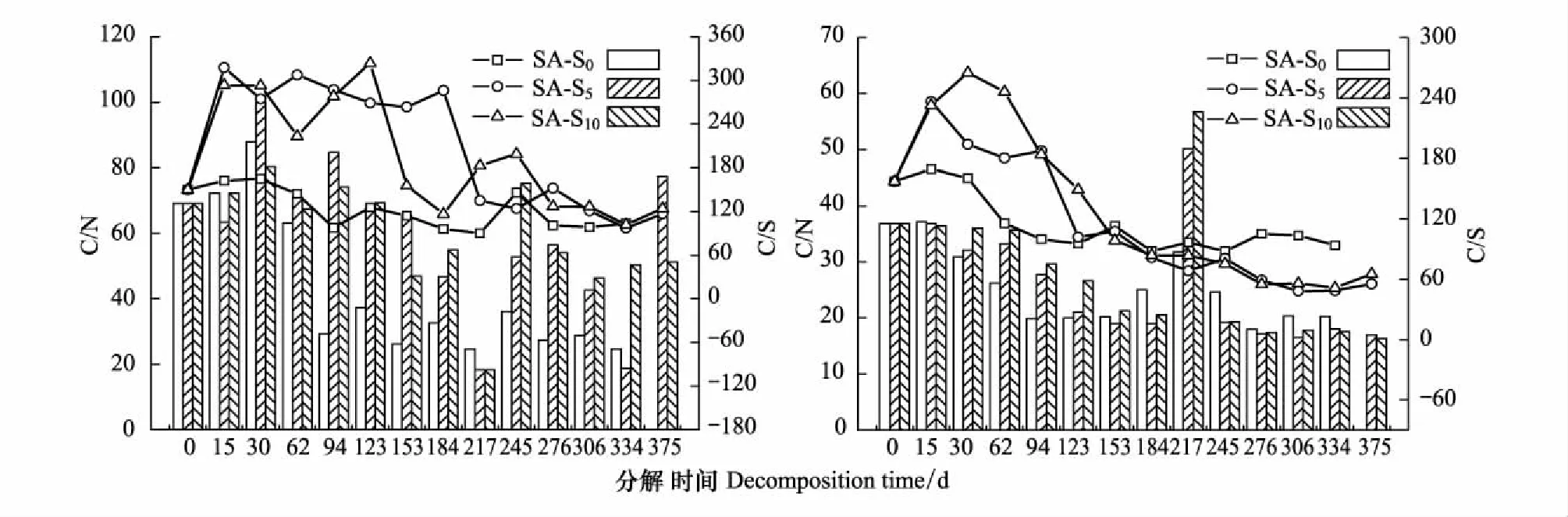
图5 不同淤积强度下残体C/N比和C/S比的变化Fig.5 Variations of C/N and C/S ratios in litters in different siltation treatments
3.2 淤积强度对残体硫含量及其释放的影响
本研究表明,随着互花米草入侵导致的淤积强度增加,两种残体在分解过程中的TS含量整体均呈增加趋势,原因与不同淤积强度下残体质量损失以及淤积作用导致的分解环境养分变化使得C/S改变有关。前述可知,尽管不同淤积强度下残体中的TS含量变化并不完全一致,但其均与相应的C/S呈相反变化(图3和图5)。相关分析表明,不同淤积强度下残体中的TS含量与残留率、C/S之间均呈极显著负相关(P<0.01)(表6)。逐步线性回归分析进一步表明,C/S和残留率对不同淤积强度下残体分解过程中TS含量的变化具有重要影响(表7)。相关研究也得到类似结果,如Sun等和李新华等在分别研究黄河口潮滩碱蓬(Suaedasalsa)和三江平原小叶章(Deyeuxiaangustifolia)残体分解时均表明,残体的硫含量变化与C/S均呈极显著负相关[21,37]。另外,不同淤积强度下互花米草和短叶茳芏残体的TS含量与C/N也均存在极显著负相关关系(表6),而残留率又与C/N呈极显著正相关(P<0.01)(表3)。因此,C/N主要通过影响残留率而间接影响残体的TS含量。本研究还表明,不同淤积强度下两种残体的TS含量在分解期间均出现阶段性升高,而这主要与微生物对分解环境中(如潮水、潮滩沉积物)的硫养分固持作用有关。李新华等在研究三江平原小叶章(Deyeuxiaangustifolia)残体分解时亦表明,残体中硫含量的明显增加主要源于微生物对沼泽水中硫的固持作用[38]。Sun等对黄河口潮滩湿地碱蓬残体分解的研究还表明,残体分解过程中硫含量的阶段性升高主要与微生物对海水和沉积物中硫的固持作用有关[21]。本研究还表明,淤积强度对短叶茳芏残体中TS含量变化的影响较互花米草更为明显,而这可能与相同淤积强度下不同残体的TS含量在很大程度上可能更取决于残留率有关,即残留率越低,TS含量越高。前述研究可知,相同淤积强度下短叶茳芏残体的分解较互花米草更快(图2和表2),由此使得其残体中的TS含量随淤积强度增加的变化较互花米草更为明显。
本研究还表明,互花米草和短叶茳芏残体在分解过程中均表现为不同程度的硫释放,且随着互花米草入侵导致的淤积强度的增加,二者的硫释放量均呈降低趋势(图4)。整体而言,不同淤积强度下两种残体的硫释放特征与其相应的残留率变化趋势较为一致(图4和图2)。相关分析进一步表明,不同淤积强度下两种残体的残留率与相应的SAI均呈显著或极显著正相关(P<0.05或P<0.01),说明不同淤积强度下残体分解过程中的干物质质量损失对其硫养分释放具有重要影响。另外,C/S和EC也是影响残体SAI变化的重要因素(表6)。前述分析可知,不同淤积强度下两种残体中的C/S和残留率均与TS含量密切相关(表6),而EC又与残留率密切相关(表3),故EC对相应残体的SAI亦可产生间接影响,且其与SAI在S0处理下还存在极显著负相关关系(P<0.01)(表6)。本研究亦表明,相同淤积强度下短叶茳芏残体的硫释放量整体要高于互花米草,这可能与短叶茳芏残体较低的初始C/N比(表1)以及分解过程中较低的C/N比有关(表5)。较低的C/N比可导致相同淤积强度下短叶茳芏残体的分解速率高于互花米草(表2),加之两种残体的硫养分释放量在很大程度上均取决于残留率(表6),由此导致短叶茳芏残体的硫释放量高于互花米草。

表6 不同淤积强度下残体TS含量和SAI与不同因子间的相关性
TS: 全硫含量, Total sulfur contents; SAI: 硫累积系数, sulfur accumulation index;*为P<0.05;**为P<0.01
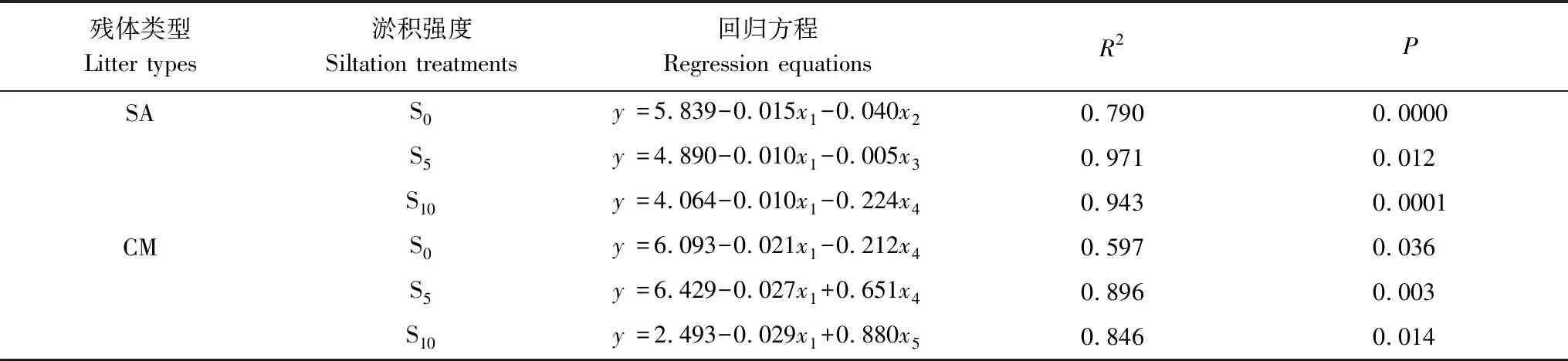
表7 不同淤积强度下残体硫含量(y)与主要因子(x)的回归方程
x1:C/S;x2:地温(Ground temperature);x3:残留率(Mass remaining);x4:EC;x5:pH
综上所述,尽管互花米草入侵过程中导致的淤积强度增加使得其自身和短叶茳芏残体分解速率及硫释放强度均降低,但就相同淤积强度而言,短叶茳芏残体的分解速率和硫释放量均大于互花米草。与短叶茳芏相比,相同生境下互花米草对硫的吸收累积能力更强[39],且其可通过快速扩张来增加湿地土壤的硫库储量,从而影响到湿地的硫养分供给能力[39-40]。根据本研究结果可得,互花米草入侵过程中导致的土壤硫库储量增加在很大程度上可能取决于短叶茳芏残体分解的贡献。
4 结论
(1)随着互花米草入侵导致的淤积强度的增加,互花米草和短叶茳芏残体的分解速率均明显降低,且这种影响在短叶茳芏残体分解过程中表现的更为显著;相同淤积强度下互花米草的分解速率低于短叶茳芏。
(2)随着互花米草入侵导致的淤积强度的增加,互花米草和短叶茳芏残体在分解过程中的TC和TS含量整体均呈增加趋势,但其对短叶茳芏残体中二者变化的影响较为明显。
(3)互花米草和短叶茳芏残体在分解过程中均表现为不同程度的硫释放,且随着互花米草入侵导致的淤积强度增加,二者的硫释放量均呈降低趋势,但相同淤积强度下短叶茳芏残体的硫释放量要高于互花米草。
(4)不同淤积强度下两种残体分解速率及硫养分释放强度的差异不仅与分解环境中的EC密切相关,而且与残留率、初始基质质量(C/N和C/S)以及淤积导致养分条件改变而对分解过程中残体基质质量的影响有关。
(5)互花米草入侵过程中导致的淤积强度增加使得其自身和短叶茳芏残体的分解速率及硫释放强度均降低;但就相同淤积强度而言,短叶茳芏残体的分解速率和硫释放量均大于互花米草。
猜你喜欢
基层中医药(2022年7期)2022-11-17
农业灾害研究(2022年7期)2022-09-08
安徽农业科学(2022年14期)2022-08-04
保健医苑(2022年5期)2022-06-10
海洋通报(2021年5期)2021-12-21
土壤学报(2021年4期)2021-09-10
能源与环境(2020年4期)2020-09-02
中国临床医学影像杂志(2019年6期)2019-08-27
生态学报(2019年8期)2019-05-31
——以咸阳市屯庄水库为例
水利科学与寒区工程(2019年2期)2019-04-28
