
杉木N、P代谢对模拟土壤增温及隔离降雨的响应
2019-07-05 10:59朱锦懋杨智杰熊德成杨玉盛陈光水陈龙龙林婉玉
生态学报 2019年10期
方 璇,王 健,王 彬,朱锦懋,杨智杰,熊德成,杨玉盛,陈光水,陈龙龙,林婉玉
1 福建师范大学生命科学学院,福州 350117 2 福建师范大学福建省植物生理生态重点实验室,福州 350007 3 福建师范大学地理科学学院,福州 350007
最近60年气温上升的幅度尤为显著,平均每10年上升0.23℃。据统计,全球平均地表温度从19世纪末开始,在1951—2012年间上升了0.89℃[1]。与此同时,在温度上升和人类行为的共同作用下,全球降水格局发生了一定的变化,具体表现为:高纬度地区降水增加,中低纬降水减少,且随着温度持续上升、降水格局变化,未来可能会引起一系列的资源、环境和生态效应改变[2- 4]。目前,有关植物控温、水分变化的研究多数集中于中高纬度地区[5-6],而对于低纬度亚热带地区植物研究较少。
植物中的养分如何响应全球气候变化是全球变化生态学的一个重要科学问题。氮(N)、磷(P)是陆地生态系统中植物生长的主要限制性元素,同时也是评估植物营养状况的必要指标,其含量的高低影响植物的生长与群落动态[7-10]。已知土壤增温和降雨变化都会对植物养分产生影响,由于研究区域、增温以及干旱处理的方式、实验材料的不同,目前已有对植物C、N、P浓度的影响并没有取得一致的结果,有升高[11]、降低[12- 13]或无明显变化[14]等。且在未来变暖环境下,温度偏差较小的亚热带树种可能比温带树种更容易受到气温变化的影响[15]。因此,有必要对亚热带树种养分特征进行进一步的研究。同时,当前对于植物养分吸收率的研究已经受到广泛关注[7],植物吸收N和P是植物保持N和P的重要内部策略,尤其是在全球环境变化导致养分利用率发生变化的情况下[16]。Yuan等[17]认为随着年均温和年均降水量下降,N吸收率增大,而P吸收率下降。然而,Vergutz等[18]却认为N、P元素的吸收率随着年均温和年均降水量的下降而增加。因此,N、P吸收率在温度和降雨条件下的变化仍存在极大争议。
杉木(Cunninghamialanceolata)是中国亚热带地区重要的速生造林树种,在中国人工林生产中发挥着重要作用[19-20],在中国亚热带森林生态系统中占有重要地位,占中国南方人工林面积的60%—80%,占全国人工林面积的26.6%[21]。本实验以杉木为研究对象,设置对照(Control)、增温(Warming, +5℃)、隔离降雨(Precipitation exclusion, -50%)和增温×隔离降雨(Warming and precipitation exclusion interaction)4个处理,每个处理5个重复,共20组,后根据“主干法”确定针叶龄级,将样品分为为一年生、二年生、三年生、四年生,共4个年份的叶片。研究增温、隔离降雨对鲜叶、凋落叶C、N、P浓度的影响,N、P吸收率的变化,以及叶片N、P浓度与其吸收率二者的关系。本研究旨在了解植物C、N、P化学计量特征和植物对N、P的吸收如何随着亚热带地区持续变暖和干旱而发生变化,结果有望为今后亚热带木本植物适应全球气候变暖的研究提供有价值的参考。
1 材料与方法
1.1 试验地概况
实验样地位于福建三明森林生态系统和全球变化研究站陈大观测点(26°19′ N,117°36′ E)。该地区属中亚热带季风气候,年平均温度为19.1℃,年平均降雨量1749 mm,降雨多集中在3—8月,年平均蒸发量1585 mm,相对湿度81%。土壤以花岗岩发育的红壤和黄壤为主。
1.2 试验设计
试验设置参照(冯建新等)[22]。试验采用两因素两水平完全随机区组设计,设置增温及隔离降雨共4个处理,即①对照(CK):不增温,无隔离降雨;②增温(W):增温5℃,无隔离降雨;③隔离降雨(Pe):不增温,隔离降雨50%;④增温×隔离降雨(WP):增温5℃,隔离降雨50%,每个处理5个重复,试验小区面积2 m×2 m。土壤增温设置:试验小区四周采用4块PVC板(200 cm×70 cm深)焊接而成。于2013年10月安装加热电缆(所有小区都布设相同电缆)。2013年11月,每个2 m×2 m小区均匀种植4棵1年生2代半短侧枝杉木幼苗,杉木位置均处于两条电缆线之间。杉木种植5个月后,在样地小区高处每隔0.05 m安放0.05 m×5 m的透明U型管,以隔离50%的降雨(试验期间总降水量为1994.2 mm)。样地布设完成后开始通电增温(2014年3月)。土壤本底资料如表1[23]。
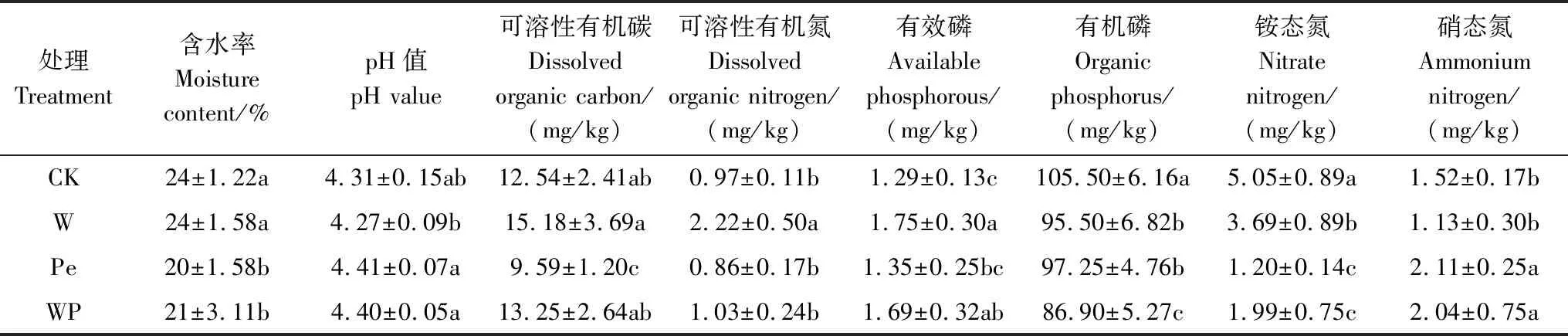
表1 不同处理的土壤理化性质
CK:对照 Control;W:增温 Warming;Pe:隔离降雨 Precipitation exclusion;WP:增温×隔离降雨 Warming and precipitation exclusion interaction,下同不同小写字母表示处理间差异显著(P<0.05)
1.3 样品采集与处理
于2017年8月份采集植物样品,在试验小区中选取植物同一高度,朝南面带有完全展开叶片的树枝,根据“主干法”将采集的样品分为不同叶龄叶片,即为一年生、二年生、三年生、四年生叶片。此外,收集各试验小区的凋落叶,分别装入标记好的信封中,带回实验室。将鲜叶与凋落叶置于60℃的烘箱中烘干致恒重,后粉碎并过0.149 mm的筛,用于测定C、N、P浓度。
叶片全磷采用硫酸-高氯酸消煮法提取,提取液用连续流动分析仪(Skalar San++,荷兰)测定;叶片碳、氮元素浓度采用碳氮元素分析仪(Elemental Analyzer Vario ELI-II,德国)测定。
1.4 数据分析与测定
N、P吸收率的计算公式如下[24-25]:
式中,NuRE表示养分吸收率,Nugreen表示鲜叶养分浓度,Nusenesced表示凋落叶养分浓度(NRE表示N吸收率,PRE表示P吸收率)。
绘图由Origin 9.0完成,采用SPSS 22.0软件对数据进行统计分析,采用考虑交互作用的双因素方差分析(two-way ANOVA)比较处理间差异。
2 结果与分析
2.1 增温及隔离降雨对杉木凋落叶C、N、P浓度及其化学计量比特征影响
如图1所示,增温、隔离降雨对凋落叶N浓度、C/N比值、N/P比值均无显著影响;但在隔离降雨处理凋落叶N浓度达到最高值8.35 mg/g; C/N比值达到最小值;在无隔离降雨的增温处理,凋落叶C浓度极显著降低(P<0.01),比对照组下降4.63%;而无增温的隔离降雨使凋落叶P浓度显著增加23.32%(P<0.05),使C/P比值显著下降18.57%(P<0.05)(图1)。
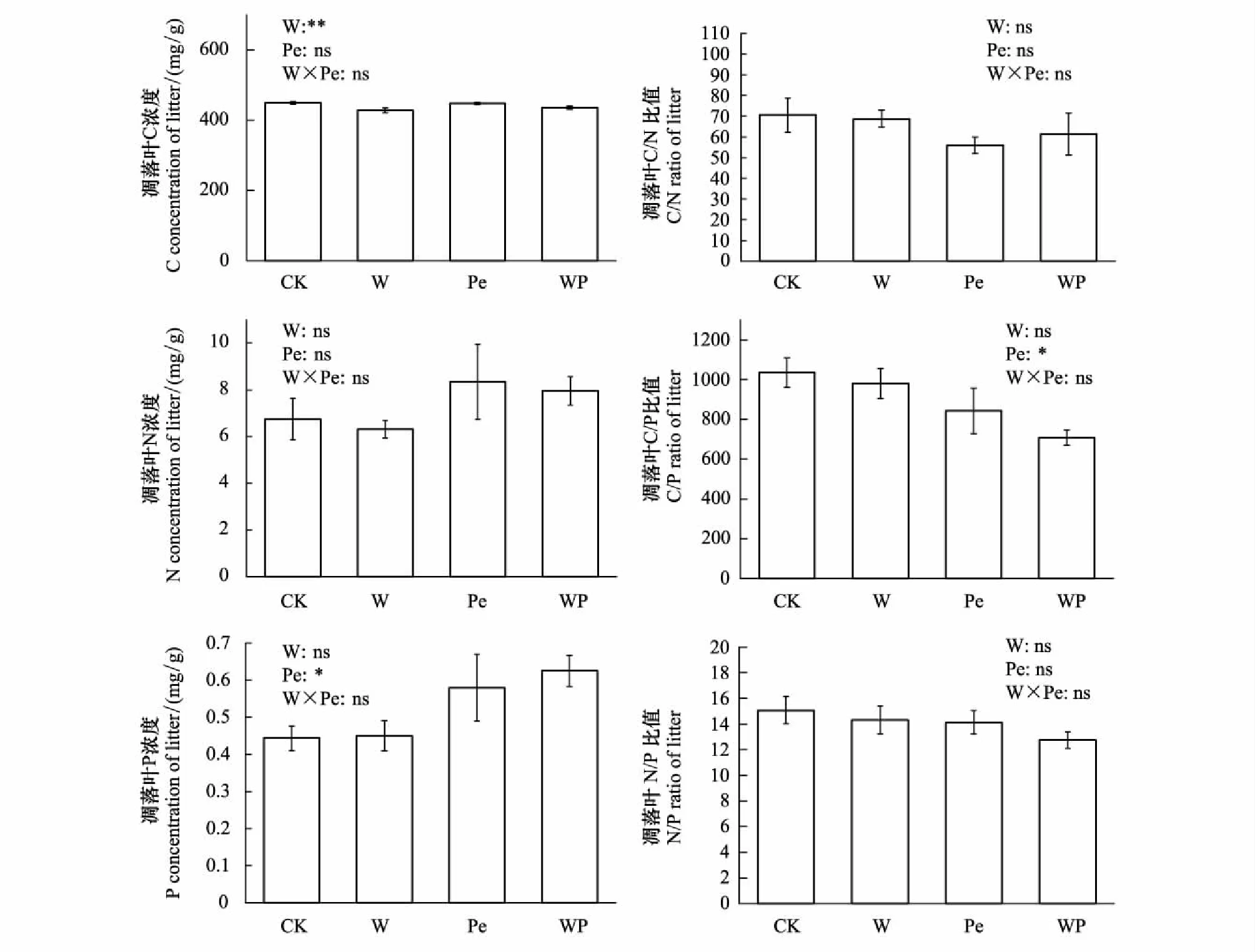
图1 模拟增温及隔离降雨对凋落叶C、N、P化学计量特征的影响Fig.1 Effects of simulated warming and precipitation exclusion on stoichiometric characteristics of litter C, N and P星号表示显著性(*P<0.05,**P<0.01), ns表示不显著
2.2 增温及隔离降雨对杉木不同叶龄鲜叶C、N、P浓度及其化学计量特征影响
增温仅对杉木一年生叶片N/P比值及二年生叶片N浓度有显著影响(图2, 图3,P<0.05)。其中增温对一年生叶片N/P比值具有显著的促进作用(P<0.05)。
隔离降雨对杉木三年生及四年生叶片N浓度有极显著的促进作用(P<0.01),二者较对照组分别增加18.15%和25.33%(图2),同时极显著降低三年生及四年生叶片C/N比值,二者较对照组分别减少17.36%和24.39%(图3,P<0.01)。
增温×隔离降雨交互作用极显著降低一年生鲜叶N/P比值(图3,P<0.01)及二年生鲜叶N浓度(图2,P<0.05)。
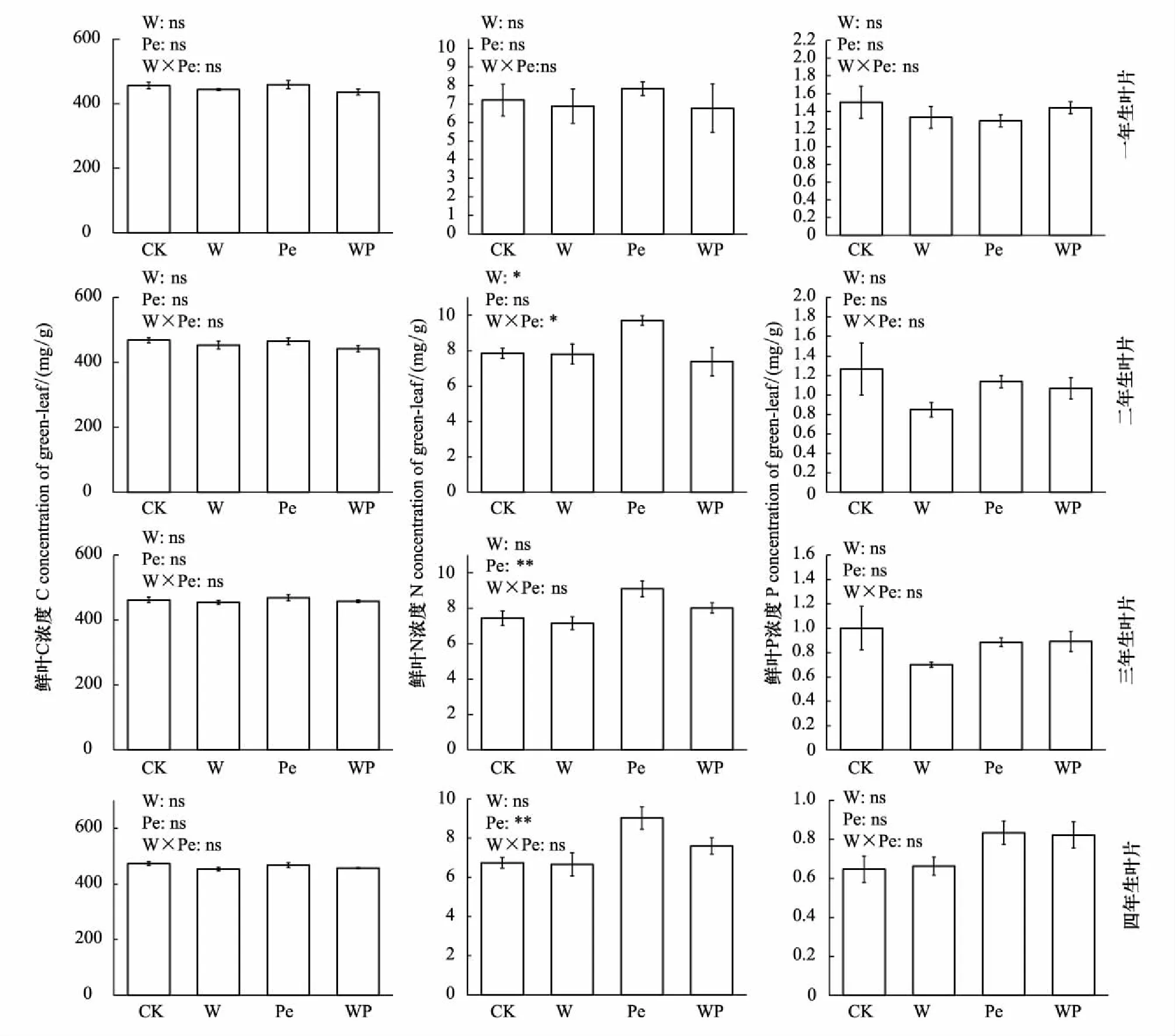
图2 模拟增温及隔离降雨对不同叶龄鲜叶C、N、P浓度的影响Fig.2 Effects of simulated warming and precipitation exclusion on C, N and P concentrations in leaves of different ages
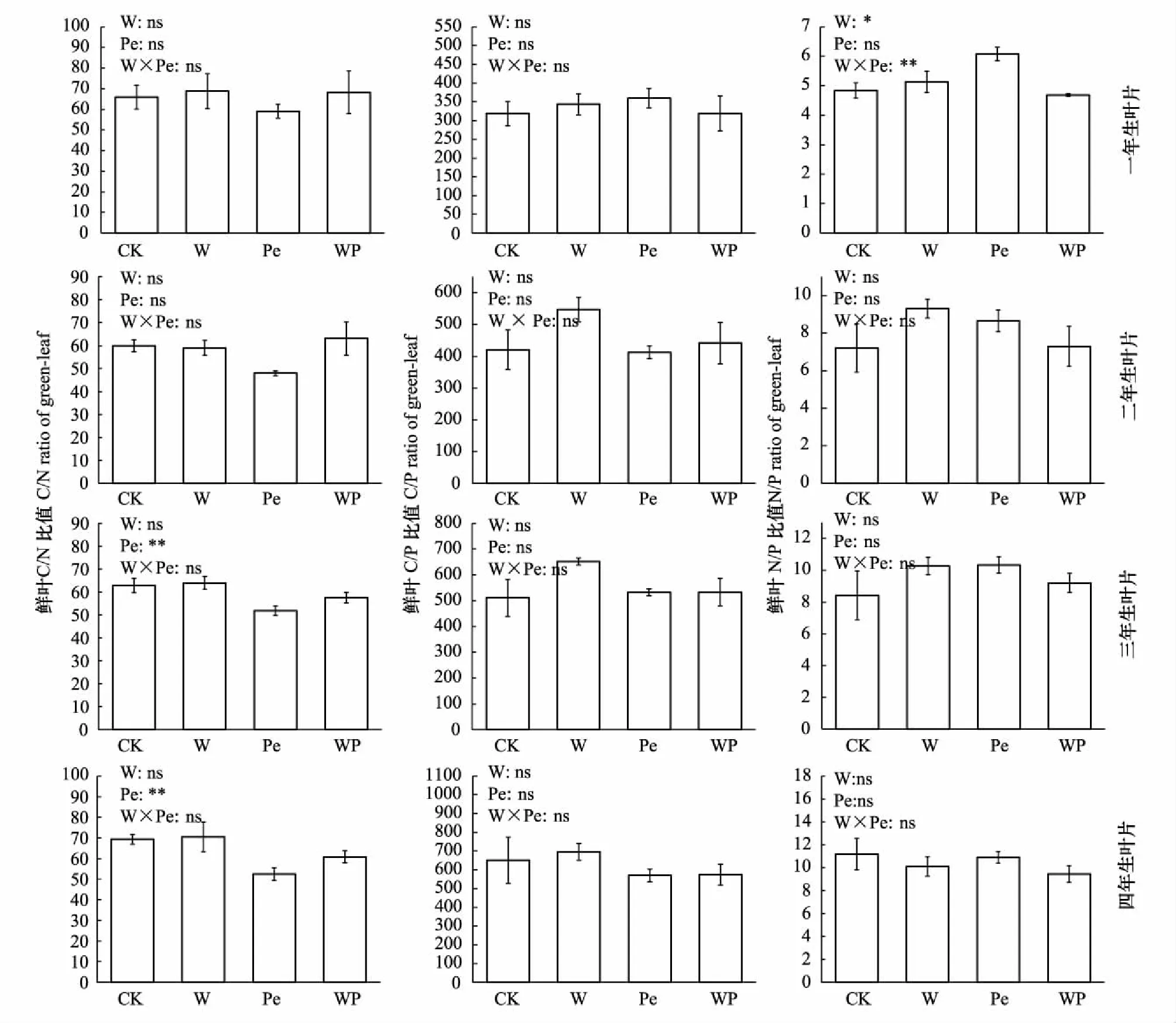
图3 模拟增温及隔离降雨对不同叶龄鲜叶C、N、P化学计量比的影响Fig.3 Effects of simulated warming and precipitation exclusion on the ratios of C, N and P in leaves of different ages
2.3 增温及隔离降雨对杉木不同叶龄鲜叶N、P吸收率影响
由于NRE波动剧烈,增温、隔离降雨、增温×隔离降雨处理杉木叶NRE与对照相比均无显著差异(图4)。其中无增温的隔离降雨使NRE呈上升趋势,而无隔离降雨的增温,增温×隔离降雨处理使NRE呈不同程度下降趋势,且同一处理不同年份的NRE差异不显著。
PRE值远大于NRE值(图4)。除一年生叶片的PRE在增温处理出现显著差异(P<0.05)外,增温、隔离降雨对其他3个年份叶片PRE均未造成显著差异。对同一处理不同年份叶PRE进行研究发现,PRE随叶龄增加呈下降趋势,即:一年生>二年生>三年生>四年生,仅隔离降雨处理下不同年份杉木叶PRE未有显著差异,而增温及增温×隔离降雨处理均使PRE随叶龄显著下降(P<0.05)。
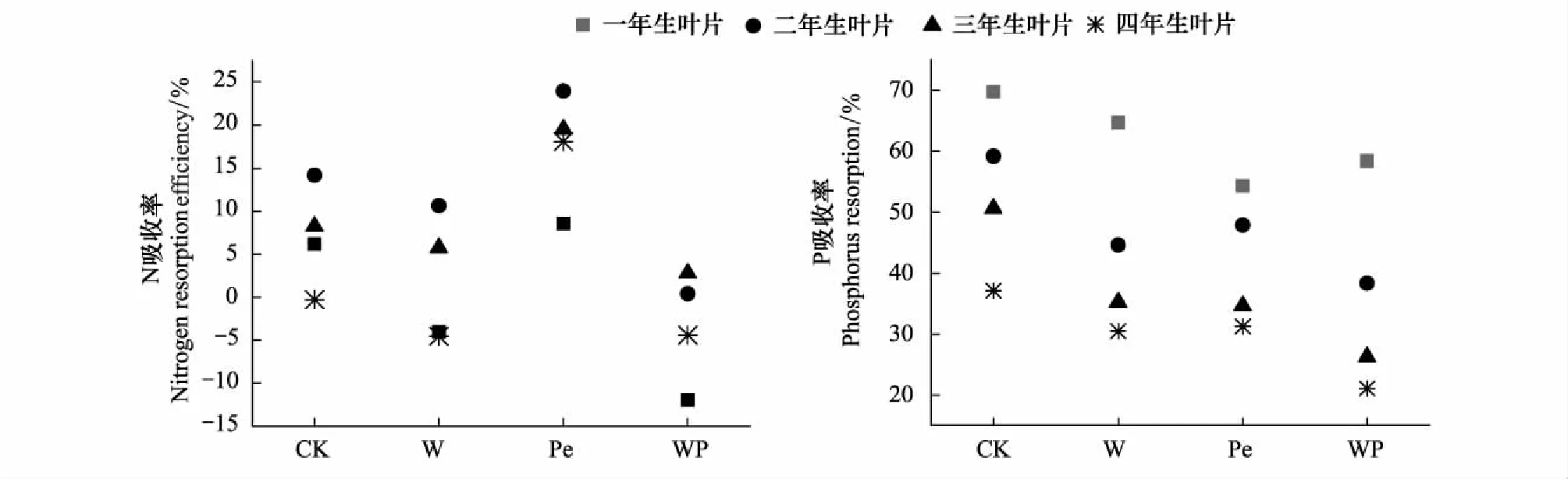
图4 模拟增温及隔离降雨对不同叶龄鲜叶N、P吸收率的影响Fig.4 Effects of simulated warming and precipitation exclusion on N and P resorption efficiency of leaves in different ages
2.4 增温及隔离降雨对杉木鲜叶N、P浓度与N、P吸收率拟合关系的影响
对鲜叶N浓度及NRE进行拟合关系分析(图5)。对照组中鲜叶N浓度与NRE的拟合度为负值,二者拟合关系极低(图5)。而增温、隔离降雨及增温×隔离降雨处理均不同程度增加鲜叶N浓度及NRE的拟合关系。其中增温及增温×隔离降雨处理鲜叶N浓度对NRE的贡献率达到极显著水平(P<0.01)(图5),而隔离降雨无显著增加二者关系(图5)。经过拟合计算,在对照组鲜叶中N浓度大于11.89 mg/g时,NRE为正值;在增温组鲜叶中N浓度大于0.62 mg/g时,NRE为正值;在隔离降雨组鲜叶中N浓度大于10.08 mg/g时,NRE为正值;在增温×隔离降雨组鲜叶中N浓度大于8.02 mg/g时,NRE为正值。
对鲜叶P浓度及PRE进行拟合关系分析,如图6所示。鲜叶P浓度与PRE关系均呈线性关系,除隔离降雨处理P浓度与其PRE显著正相关外(P<0.05,图6),其他处理组下的P浓度与其PRE都呈现极显著的正相关(P<0.01,图6)。与N浓度及NRE拟合关系结果正好相反,不同处理均降低了鲜叶P浓度与PRE拟合关系。

图5 四个处理组下鲜叶N浓度与N吸收率(NRE)的拟合关系Fig.5 Fitting relationship between N concentrations and N resorption efficiency of leaves in four treatments
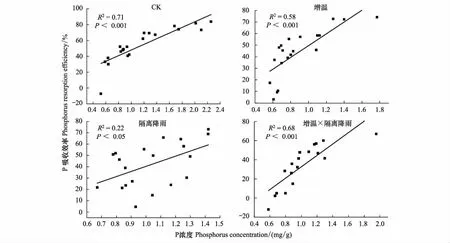
图6 四个处理组下鲜叶P浓度与P吸收率(PRE)的拟合关系Fig.6 Fitting relationship between P concentrations and P resorption efficiency of leaves in four treatments
3 讨论
3.1 增温及隔离降雨对凋落叶C、N、P化学计量特征的影响
凋落叶在陆地生态系统养分循环和土壤肥力的维持中起着重要作用,土壤温度和土壤水分含量可直接和间接地影响凋落叶分解过程,使保留在凋落叶中的养分浓度发生变化[26-27]。本研究结果显示无隔离降雨的增温处理显著降低凋落叶C浓度,无增温的隔离降雨处理使凋落叶P浓度显著增加,C/P比值显著下降。增温导致凋落叶C浓度显著降低的原因可能是土壤增温导致土壤C损失,同时刺激树木木质组织中C增加[28],使凋落叶C分解速率加快,以补充土壤中损失的C。同时,Cai等[29]认为凋落叶中较高的营养浓度会降低其吸收率,并据Killingbeck[30]表述,如果凋落叶中P浓度小于0.05 mg/g,认为植物中具有较高的P吸收力。因此推测,无增温的隔离降雨可能使植物中P吸收率下降,导致更多P保留在凋落叶中,由于未造成C浓度显著差异,从而导致C/P比值下降。
3.2 增温及隔离降雨对鲜叶C、N、P浓度及生态化学计量特征的影响
在陆地生态系统中,植物体内的 N、P 含量及其化学计量比很容易受到温度等非生物因素的影响[31]。Bai等[11]通过meta分析认为植物中鲜叶N浓度与温度呈正相关。但有相反研究表明随温度增加叶片N、P浓度呈下降趋势[13, 32]。而彭阿辉等[33]对青藏高原高寒草甸和高寒沼泽中两种优势物种模拟增温实验,发现增温仅降低两种草的N浓度,而对P浓度没有影响。本研究结果显示无隔离降雨的增温处理对P浓度无显著影响,但使其呈下降趋势,同时仅使二年生叶片N浓度显著下降,一年生叶片N/P比值显著上升。造成此现象的原因可能是一年生叶片是整个植物体生长最旺盛的部位之一,也是养分集中处,因此,外界对其养分影响较小;而三、四年生叶片为成熟叶,自身已有营养储藏则对外界环境变化响应较弱。而造成P浓度呈下降趋势,N浓度显著下降的主要原因是:一方面增温诱导土壤N矿化和硝化速率的增加[28],让更多养分固定在土壤中,使植物吸收的养分减少;另一方面增温还提高了植物的生物化学反应速率,使富含N的酶及富含P的RNA活性上升,导致植物体内 N、P浓度下降来增加其在温度变化下的适应能力和新陈代谢水平[34]。
干旱是不同植物器官和生态系统元素化学计量变化的潜在驱动因素[35-36]。Chen等[19]研究发现干旱能够提高叶片N浓度,本研究也显示无增温的隔离降雨促进三四年生叶片中N浓度极显著上升,C/N比值极显著下降。说明较高的叶片N浓度是植物适应干旱条件的反映[37],叶片N浓度增加有助于植物在较小气孔导度下达到最大光合容积,提升了水分利用效率,降低光合作用的水分散失[38]。植物C/N、C/P比值意味着植物吸收营养用以同化C的能力,在一定程度上反映了植物的养分利用效率[39]。表明本地区杉木老叶在隔离降雨处理同化C的能力小于N的吸收能力,导致C/N比值下降。See等[40]及Jie等[41]对苹果树(Malusdomestica)进行干旱胁迫发现P浓度下降。而本研究中隔离降雨未造成鲜叶P浓度显著差异,造成不同结果的原因可能是研究物种不同以及不同环境下植物对养分吸收的选择性不同[42]。本文结果说明本地区杉木鲜叶N浓度较P浓度对水分更为敏感,且适度干旱可以缓解N限制植物生长的情况。
在增温×隔离降雨处理下,新叶N浓度下降而老叶上升,P浓度无显著变化。两种环境因子的结合可能通过增温导致植物内部水分供应的减少来增强隔离降雨的影响,且新叶较老叶对水分更加敏感,适度的水分亏缺会增加N浓度,过度的水分亏缺可能造成N浓度下降。增温及隔离降雨的交互作用还使一年生叶片N/P比值极显著下降。Dijkstra等[10]研究表明干旱胁迫使植物N、P浓度均下降。增温会加剧土壤的水分亏缺程度,本研究结果可能是增温×隔离降雨减少土壤水分导致微生物活动和氮矿化作用减弱,同时降低养分流动率,引起植物养分的吸收减少和土壤中N或P有效性下降,且在增温和隔离降雨交互作用下导致严重水分亏缺条件下,P较N更为敏感[10]。
3.3 增温及隔离降雨对N、P吸收率的影响
植物的养分内吸收与植物从土壤中摄取养分几乎同样重要,这减少了对土壤可利用性养分和根部吸收养分的依赖[43]。增温、增温×隔离降雨均使不同年份的杉木叶NRE呈下降趋势,而隔离降雨使NRE呈上升趋势。Yuan等[44]对北美草原的增温实验表明增温降低5种优势物种NRE。而Aerts等[45]对瑞士亚北极泥碳地的气候变化研究表明,增温对多年生物种的NRE基本无影响。这种不一致可能是由于增温处理时间、增温幅度、生态系统不同所导致的[46]。已有研究中干旱对植物NRE影响也结果不一。Jordi等[47]对地中海森林植物隔离30%的降雨,结果显示干旱处理降低杨梅(Arbutusunedo)叶片NRE,与本研究结果不同。造成此差异可能是由于干旱程度不同造成的水分亏缺程度不一致,导致不同的土壤淋溶作用程度不同,低估凋落物中至少8%和10%的氮、磷元素的浓度[48],进而影响NRE。而增温×隔离降雨使NRE下降可能是增温及隔离降雨均导致土壤的含水率下降[11],有研究显示在干旱条件下,增温会增强植物的水分胁迫作用,降低植物对N的吸收能力[49]。
PRE是节约P元素和提高植物在恶劣环境中生存的关键过程,在本研究中增温、隔离降雨、增温×隔离降雨均使PRE下降,其中隔离降雨使一年生叶片的PRE显著下降。已有研究显示PRE对温度的响应与NRE相同,但并没有一个清晰的结论[40, 50- 51]。水分亏缺会造成PRE下降[52],如:See等[40]对温室栽培的苹果树(Malusdomestica)进行干旱胁迫处理,叶片P浓度低于对照组,表明干旱胁迫降低了叶片对P元素的吸收率[41]。
本研究中NRE为-17%—17.1%,而PRE为21%—69.7%,说明植物对自身养分的吸收率存在物种差异,其中NRE出现负值的原因可能是叶子在衰老期间富集N素导致[25]。杉木叶PRE较NRE高得多,这是由于植物能够从土壤中吸收更多的N,减少从衰老叶片中转移N(如Huang等的研究结果[53]),而P在土壤中的移动性比N小得多,且不能在生态系统边界上轻松移动[54],因此较多的P留在土壤中被植物和土壤微生物所利用,同时微生物的竞争力大于植物[55],导致植物需要从衰老叶片中获取更多的P来满足植物生长需要。Gallardo等[56]认为在P受限制的土壤中,有较高的PRE,进一步说明本实验植物生长受P限制,而本地区土壤不存在缺N现象,或者杉木从土壤中获取N比获取P更为容易。
3.4 叶片N、P浓度与其吸收率的关系
在物种内以及在物种和环境变化之间,鲜叶中养分浓度是衰老叶片养分浓度的重要决定因素,因此认为养分吸收率受鲜叶营养物质浓度影响。以往大部分研究都是通过分析土壤N、P元素来讨论植物的养分吸收率及其机制[57- 58]。然而,尽管在不良环境中植物通常具有较低的叶片营养浓度,但养分可利用性与吸收率之间的负相关关系并未普遍存在,鲜少有研究考虑植物随着自身N、P浓度变化时,PRE、NRE变化情况。我们的研究结果显示,在对照组中鲜叶N浓度与NRE的拟合度极低,而无论增温、隔离降雨或者二者交互作用下,NRE与N浓度拟合度均极显著上升,猜想造成全球变化增强N浓度和NRE拟合度变化的原因可能是N元素在自然界中较活泼,受到全球变化影响后,对环境改变做出一定的适应性反应。在增温条件下,当鲜叶中的N浓度大于0.62 mg/g时,叶片具有吸收N效应,而在隔离降雨条件下,只有当N浓度达到10.08 mg/g时,植物叶片才能吸收植物内N,进一步说明水分对植物NRE影响较大。在不同处理中鲜叶P浓度与PRE呈显著正相关,即P浓度高时鲜叶PRE高,且与对照相比其他处理拟合度更小,鲜叶的P浓度对叶片的PRE具有指示作用,同时温度、水分等外界因素削弱PRE和鲜叶P浓度拟合度,猜想造成此现象的原因是P元素不能在自然界中轻松移动,因此当外界环境改变时,P的利用效率提高以适应恶劣环境的P需求故使其不随环境变化而改变。与N元素的情况相似,水分对P浓度及PRE拟合度的影响也较大。
猜你喜欢
山西冶金(2021年3期)2021-07-27
农民致富之友(2020年8期)2020-05-11
现代园艺(2017年23期)2018-01-18
现代园艺(2017年23期)2018-01-18
中央民族大学学报(自然科学版)(2017年4期)2017-06-11
饮食科学(2016年3期)2016-07-04
饮食科学(2016年3期)2016-07-04
浙江农业科学(2016年11期)2016-05-04
中国茶叶加工(2015年3期)2015-02-27
中国茶叶加工(2015年3期)2015-02-27
