
梨小食心虫过冷却点及结冰点测定
2019-06-27 02:07孔维娜王怡郭永福
植物保护 2019年3期
孔维娜 王怡 郭永福

摘要 梨小食心虫Grapholita molesta(Busck)是世界性的果树害虫,以老熟幼虫越冬。由于果树新品种的选育以及气候变暖等原因,梨小食心虫寄主植物的适生区扩大。为了明确梨小食心虫的耐寒能力,即是否可以随着寄主植物种植范围的扩大而向寒冷地区发生迁移,本试验对不同虫态、不同龄期幼虫、不同性别的蛹与成虫以及来源于不同环境下的幼虫的过冷却点和结冰点进行测定,进而为判断梨小食心虫是否可以顺利越冬提供理论依据。结果表明:(1)梨小食心虫不同发育阶段的过冷却点及结冰点差异显著,结冰点和过冷却点表现为幼虫>蛹>成虫;(2)不同龄期幼虫的过冷却点及结冰点差异显著。5龄幼虫的过冷却点(-12.47±0.27)℃与结冰点(-8.21±0.41)℃最低,4龄幼虫的过冷却点(-9.23±0.38)℃与结冰点(-5.06±0.37)℃最高;(3)雌雄成虫与蛹的过冷却点及结冰点差异不显著;(4)不同环境下幼虫的过冷却点及结冰点差异显著,外采幼虫的过冷却点(-24.42±0.69)℃和结冰点(-10.21±0.71)℃最低,室内幼虫经冷驯化后过冷却点降低。因此,在越冬过程中梨小食心虫老熟幼虫的抗寒能力较强,提前经历低温环境可提高其抗寒能力。
关键词 梨小食心虫; 过冷却点; 结冰点; 耐寒性
中图分类号: S 433.4
文献标识码: ADOI: 10.16688/j.zwbh.2018273
Abstract Grapholita molesta (Busck) is a worldwide pest of fruit trees, and overwinters as mature larvae. The suitable growing area of host plants has been enlarged with the new variety breeding of fruit trees and climate warming. In order to determine the cold resistance of G.molesta, or is it possible to migrate to the cold areas following the enlargement of suitable growing area of host plants. The supercooling points and freezing points of different development stages, larval instars, sex of pupae and adults, and conditions were measured, so as to judge whether they could overwinter successfully or not. The results showed that there was no significant difference in the supercooling points and freezing points for the two sexes of pupae and adults, and the supercooling points and freezing points differed among different developmental stages and larval instars, and larval conditions. The ranking of the supercooling points and freezing points for different developmental stages was as followed: larva>pupa>adult. The supercooling point (-12.47±0.27)℃ and freezing point (-8.21±0.41)℃ of 5th instar larvae were lowest, whereas the supercooling point (-9.23±0.38)℃ and freezing point (-5.06±0.37)℃ of 4th instar larvae were highest. When compared with indoor larvae without cold acclimation, outdoor larvae had a lower supercooling point (-24.42±0.69)℃ and freezing point (-10.21±0.71)℃, but the supercooling point of indoor larvae could be reduced by cold acclimation. Therefore, our results indicated that the cold resistance of mature larvae was strong, and it could be improved by cold acclimation.
Key words Grapholita molesta; supercooling point; freezing point; cold tolerance
梨小食心蟲Grapholita molesta(Busck),又名东方蛀果蛾,桃折梢虫,属鳞翅目Lepidoptera、卷蛾科Tortricidae,简称梨小,是世界性的蛀果类害虫[12]。在我国,除西藏未见报道外,其余地区均有分布[3]。主要为害的寄主包括桃、梨、苹果、山楂、李、杏、枇杷、木瓜等[4],其中,梨在秦岭淮河以北地区分布较多[5],苹果、杏在温带地区分布较多,李、枇杷、木瓜等主要在我国南部分布。近年来对果树适生区进行调查发现,由于果树新品种的开发,以及气候变暖,梨小食心虫寄主植物的分布范围在不断扩大。例如,对西藏果树资源考察发现,雅鲁藏布江的中游河谷地带为温暖半湿润区,是苹果、桃、梨的适宜种植区[6];抗寒杂交品种‘寒红梨已在东三省广泛引种试栽[7]。这不仅会使得已知分布区内梨小食心虫为害加重,而且还会随着寄主植物种植范围的扩大而发生迁移,从而给果农造成严重的经济损失[8]。
昆虫不同发育阶段均受温度的影响,为了判断梨小食心虫是否可以随着寄主植物种植面积的扩大而向冬季更为寒冷的地区迁移并成功定殖,耐寒性应进行详细评估。
昆虫耐寒性是指昆虫长期或短期暴露于低温下的存活能力,昆虫通过生理生化调节体内自身抗寒物质的变化,从而在恶劣的气候当中从生理上做好抗寒准备[9]。过冷却点常常作为抗寒能力的一个重要指标来衡量昆虫抗寒能力的强弱[1011]。
为了探索梨小食心虫各发育阶段的抗寒能力以及是否可以随着寄主植物的向北扩散而成功定殖,本试验通过对比梨小食心虫不同虫态(幼虫、蛹、成虫)、不同性别的蛹和成虫、不同龄期幼虫(2至5龄)及不同环境下的幼虫(冷藏处理幼虫、室内幼虫、外采幼虫)的过冷却点和结冰点,探讨梨小食心虫不同发育阶段的耐寒能力,为梨小食心虫的预测预报提供一定的理论基础。
1 材料与方法
1.1 供试虫源
本试验外采幼虫以及室内饲养虫源均采自山西省太谷县桃园。
室内饲养虫源为在实验室继代饲养超过50代的梨小食心虫种群,每年通过野外采集对室内种群进行复壮。成虫均统一饲养于高30 cm的玻璃养虫罐,养虫罐底部放一张稍大于下方口径的圆形硫酸纸,同时侧壁下方在罐内围一圈硫酸纸。以直径22 cm的塑料盆为底座,养虫罐上方开口用皮筋绑一块纱布,从上部悬挂一个棉球(直径为2.5 cm),每日补充5%的蜂蜜水,保证棉球不滴水为宜。每罐内约饲养200头成虫。罐内放有小的青苹果,3~4个为宜。将萼洼部分朝上放置,果柄剪掉[12]。幼虫饲养于装有人工饲料的指形管中(指形管内径1.8 cm,高度为8 cm),待幼虫化蛹后,将蛹转移至化蛹盒中,并将羽化后的成虫接入养虫罐。梨小食心虫的整个生活史均在可调控的智能人工气候箱(MGC450HP)内完成,气候箱内温度为(26±0.8)℃,相对湿度为70%~80%,光周期为L∥D=15 h∥9 h。
冷藏幼虫:将室内饲养的5龄幼虫放置于5℃低温环境下,1 d后取出用于试验。
外采幼虫于2016年11月采集获得,直接用于试验。
1.2 测定方法
试验选取发育情况相近(各测量虫态均为上一虫态或龄期蜕皮后发育的中期)的健康幼虫进行过冷却点测定。测定前用滤纸将虫体表面的水分吸干,利用热电偶原理对过冷却点和结冰点进行测定,测定时将虫体套入与其大小一致的塑料软管中,将与温度检测仪连接的感温探头插入塑料软管中。其中,对幼虫和成虫进行测定时,将感温探头固定于腹部,对蛹进行测定时,将感温探头固定于背板。将虫体置入-25℃的低温培养箱中测定过冷却点(Scp)和结冰点(Fp)。重复测定30头以上。为避免虫体降温速率过快,在虫体与感温头固定后,用泡沫塑料盒包裹外部,使虫体以约 1℃/min 的速率降温。每个处理结束后用冰水混合物对记录仪进行校正。过冷却点仪为UR. 100, 4125(Yologma Elect. Co, Seoul, Korea)。
1.3 数据处理方法
对不同处理间数据进行平方根(Sqrt)转换,将原始数据与转换后数据均进行正态性检验(ShapiroWilk test),若符合正态性分布则进行单因素参数检验(Oneway ANOVA)分析显著性,并进行Turkey法多重比较;若不符合正态性则进行单因素非参数检验(KIndependent Samples)分析显著性,并用DunnBonferroni法分析两两处理间差异。通过两独立样本t检验,比较分析性别间差异。数据处理软件为IBM SPSS v.19.0。
2 结果与分析
2.1 不同虫态的梨小食心虫过冷却点及结冰点
梨小食心虫3种不同虫态的过冷却点差异显著(F=319.94, df=2, 239, P<0.05,表1),成虫过冷却点最低,为(-20.58±0.5)℃;蛹次之,为(-17.95±0.29)℃;幼虫最高,为(-10.6±0.2)℃。不同虫态的结冰点也存在显著性差异(F=119.51, df=2, 249, P<0.05,表1),成虫的结冰点最低,为(-13.33±3.94)℃;其次为蛹(-12.31±0.42)℃,幼虫最高,为(-6.75±0.22)℃,成虫与蛹的结冰点无显著差异。
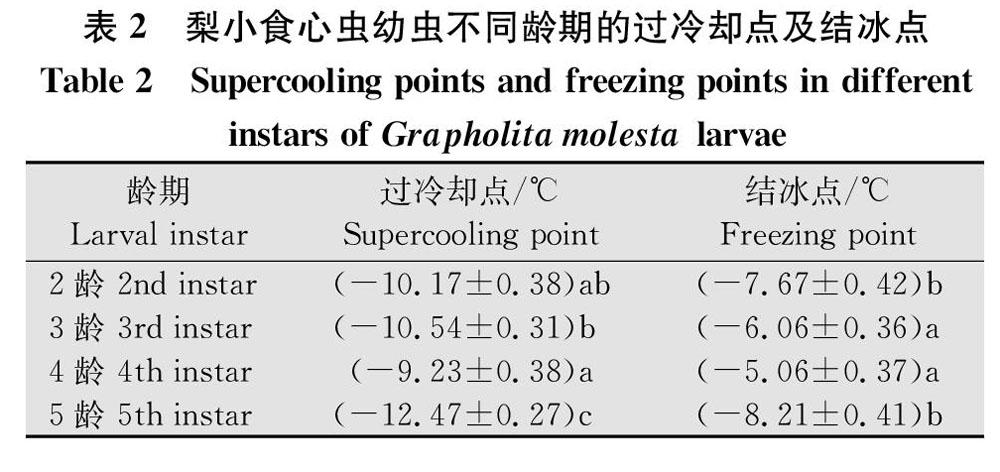
2.2 梨小食心虫不同龄期幼虫的过冷却点及结冰点
幼虫不同龄期之间的过冷却点差异显著(F=16.23, df=3, 119, P<0.05,表2),其中,5龄幼虫的过冷却点最低,为(-12.47±0.27)℃;4龄幼虫的过冷却点最高,为(-9.23±0.38)℃。幼虫不同龄期的结冰点也存在显著差异(F=13.65, df=3, 119, P<0.05,表2),5龄的结冰点最低(-8.21±0.41)℃,4龄的结冰点最高(-5.06±0.37)℃。
2.3 梨小食心虫不同性别蛹与成虫的过冷却点及结冰点
雌、雄蛹的過冷却点差异不显著(t=-1.94, df=58, P=0.054,表3),雌蛹过冷却点略低于雄蛹。结冰点也不存在显著性差异(t=1.77, df=41.75,P=0.084,表3),雌蛹结冰点略高于雄蛹。雌雄蛾的过冷却点和结冰点均没有显著差异(过冷却点:t=-1.42,df=58,P=0.16;结冰点:t=0.24,df=67,P=0.81,表3),雌蛾过冷却点略低于雄蛾,结冰点略高于雄蛾。
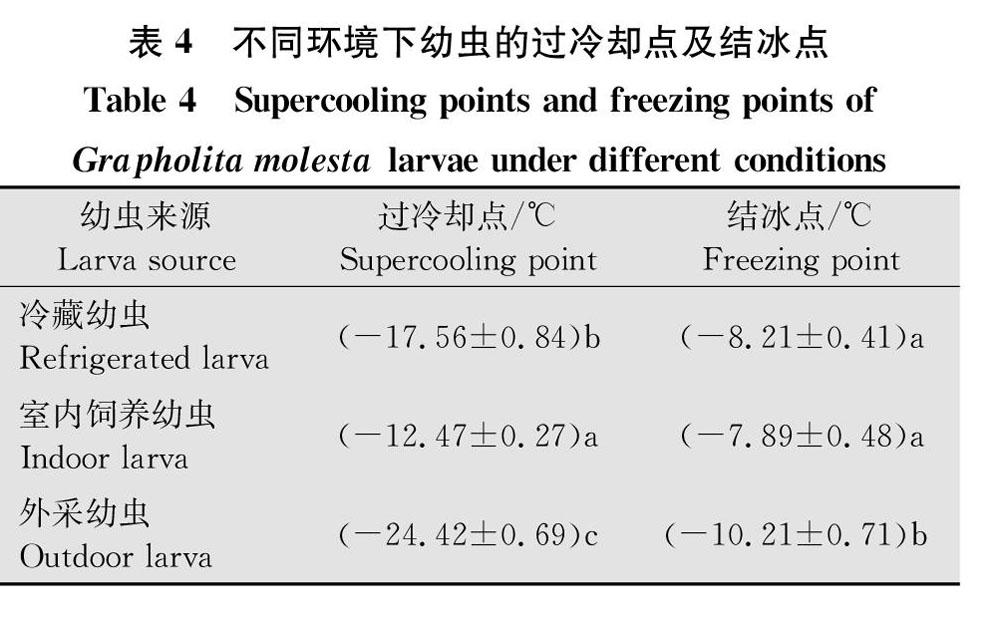
2.4梨小食心虫不同环境下幼虫的过冷却点及结冰点
对于不同环境下(冷藏处理、室内饲养、外采)的幼虫过冷却点及结冰点测定分析表明,不同环境下幼虫的过冷却点存在显著差异(F=86.16,df=2,89,P<0.05,表4),外采幼虫过冷却点最低,为(-24.42±0.69)℃;冷藏处理幼虫次之,为(-17.56±0.84)℃;室内饲养幼虫最高,为(-12.47±0.27)℃。幼虫的结冰点差异显著(F=5.28,df=2,94,P<0.05,表4),外采幼虫的结冰点最低(-10.21±0.71)℃,冷藏处理的次之(-8.21±0.41)℃,室内饲养幼虫的过冷却点最高,为(-7.89±0.48)℃,冷藏幼虫和室内饲养幼虫间结冰点差异不显著。
3 结论与讨论
本试验不同虫态的过冷却点以及结冰点的对比试验,选取的是非越冬代的梨小食心虫。此时,相较于蛹和成虫,幼虫的过冷却点最高。若在梨小食心虫非越冬代的发生期遇到超低温恶劣环境,由于幼虫抗寒性差,会使幼虫在自然条件下大量死亡,使得下一代成虫数量减少。然而,外采的越冬代老熟幼虫的过冷却点明显高于非越冬代的成虫和蛹。昆虫耐寒性除受到个体不同发育阶段的影响外,还受到季节性变化,地理环境等的影响[9]。冬季种群和高纬度及高海拔地区的种群耐寒性强于夏季种群和低纬度及低海拔地区种群[9]。
对比不同龄期幼虫(2~5龄)的过冷却点,在4龄出现过冷却点上升的趋势,到了5龄老熟幼虫期显著低于其他龄期。在田间调查中发现,梨小食心虫主要以老熟幼虫在树干翘皮和距离树干0~1.0 m范围的表层土壤中越冬,其最适深度范围为0~3.0 cm[13]。5龄老熟幼虫较低的过冷却点可帮助梨小食心虫顺利越冬。昆虫通过自身生理生化反应调节体内抗寒相关物质,如:体内脂肪、抗冻蛋白、水分、碳水化合物等的含量,从而为越冬做好准备。5龄幼虫为了适应越冬环境,将体内的糖类、蛋白质等物质转化为脂肪聚集,以此增加抗寒性。在相关研究报道中,山东泰安地区的梨小食心虫4龄之前幼虫的蛋白质含量随着虫龄的增加而增加,但不同虫龄间无显著差异。不同虫龄在经0℃低温诱导后,4龄幼虫的蛋白质及糖类含量显著低于其他龄期[14],即4龄幼虫体内的糖类和蛋白质转化为抗寒物质的速率更快。在本试验中4龄幼虫的过冷却点和结冰点均最高,推测4龄幼虫这一现象出现的原因是由于大量积累糖类和蛋白质导致[15]。同时,4龄幼虫较快地将大量积累的糖类和蛋白质转化为脂肪等抗寒物质,使得5龄老熟幼虫出现较低的过冷却点和结冰点。这一现象值得下一步继续研究。
对比不同性别的蛹及成虫的过冷却点及结冰点,雌雄间虽不存在显著性差异,但是无论蛹还是成虫,雌性的过冷却点均略低于雄性,表明雌性抵御低温的能力略强于雄性。梨小食心虫在越冬结束后,雌雄虫开始交配繁殖后代,而雌虫除交配外还需寻找寄主植物进行产卵。因此,雌虫体内的脂肪含量或者抗冻蛋白的含量高于雄虫,即雄性生理生化的调节能力及强度不及雌虫[16]。在这种情况下,秋冬如遇极端低温恶劣环境,雄虫死亡数量更高,从而导致第二年越冬成虫种群性比偏雌。由于梨小食心虫雌虫只可交配一次,雄虫可多次交配,因此,在一定程度上减轻了低温对种群的影响,在种群水平上提高了梨小食心虫抵御低温的能力。
当幼虫在经过较低温胁迫之后,可以增加其抵御低温损伤的程度,即低温冷驯化[17],在本试验中,经过冷驯化的幼虫可以降低其过冷却点,从而提高耐寒能力,但结冰点与室内饲养幼虫无显著差异性,较短时间的冷驯化并未影响幼虫的结冰点。相较于过冷却点,结冰点更难被改变。然而,经低温冷驯化后幼虫的过冷却点仍高于外采幼虫,除冷驯化的时间较短、冷驯化的温度较高之外,昆虫的抗寒性还受许多外界因子的影响,例如,在自然环境下,取食的丰富度,气候环境的季节性变化等都能提高幼虫的抗寒性。在自然环境中,若越冬代幼虫遇一定时间超低温环境,则会进一步降低幼虫的过冷却点,使得抗寒能力增强。
是否可以顺利越冬是梨小食心虫适生区的判断依据之一。近年来,随着果树抗寒品种的培育,以及全球气候变暖[18],导致部分梨小食心虫非适生区变为适生区。明确梨小食心虫过冷却点及其受环境影响后的变化规律,有助于更为精确地划定适生区,预测种群的扩散和迁移,从而制定相应的防治策略。
参考文献
[1] ROTHSCHILD G H L, VICLER R A. Biology, ecology and control of the oriental fruit moth [M]∥VAN DER GEEST L P S, EVENHUIS H H. Tortricid Pests: Their Biology, Natural Enemies and Control. Amsterdam: Elsevier, 1991: 389412.
[2] 范仁俊,刘中芳,陆俊姣,等.我国梨小食心虫综合防治研究进展[J].应用昆虫学报,2013,50(6):15091513.
[3] 陆鹏飞,黄玲巧,王琛柱.梨小食心虫化学通信中的信息物质[J].昆虫学报,2010,53(12):13901403.
[4] MYERS C T, HULL L A, KRAWCZYK G.Seasonal and cultivarassociated variation in oviposition preference of oriental fruit moth (Lepidoptera: Tortricidae) adults and feeding behavior of neonate larvae in apples [J]. Journal of Economic Entomology, 2006, 99(2): 349358.
[5] 胡国谦,陈凯,周维灼,等.我国优质梨分布区域及其适宜生态指标的研究[J].生态农业研究,1993,1(2):7075.
[6] 段盛烺,宗学普,刘效义,等.西藏果树资源考察初报[J].园艺学报,1983,10(4):217224.
[7] 丁丽华.抗寒优质红皮梨新品种寒红梨选育报告[C]∥中国园艺学会第七届青年学术讨论会论文集,2006.
[8] 陈静,杨祎红,马天文,等.梨小食心虫在蟠桃园的危害和发生动态初探[J].安徽农业科学,2009,37(30):1474514751.
[9] 景晓红,康乐.昆虫耐寒性研究[J].生态学报,2002,22(12):22022207.
[10]黄聪,李再园,王福莲,等.柑桔大实蝇老熟幼虫和蛹过冷却点和结冰点的测定[J].环境昆虫学报,2014,36(1):1721.
[11]侯柏华,张润杰.桔小实蝇不同發育阶段过冷却点的测定[J].昆虫学报,2007,50(6):638643.
[12]BAKER T C, MEYER W, ROELOFS W L. Sex pheromone dosage and blend specificity of response by oriental fruit moth males [J].Entomologia Experimentalis et Applicata,1981,30(3): 269279.
[13]赵爱平,孙聪,展恩玲,等.梨小食心虫越冬场所调查及性诱剂诱捕距离初探[J].中国植保导刊,2016,36(12):2428.
[14]凌飞.三种果树食心虫发生为害和梨小食心虫耐寒性研究[D].泰安:山东农业大学,2011.
[15]张瑞,马纪.昆虫过冷却点的影响因素概述[J].天津农业科学,2013,19(11):7684.
[16]郭海波,许永玉,鞠珍,等.中华通草蛉成虫抗寒能力季节性变化[J].生态学报,2006,26(10):32383244.
[17]王宪辉,齐宪磊,康乐.昆虫的快速冷驯化现象及其生态适应意义[J].自然科学进展,2003,13(11):11281133.
[18]杜军,马鹏飞,潘多.1981-2014年西藏各时次气温的变化趋势分析[J].地理学报,2016,71(3):422432.
(责任编辑:田 喆)
