
浅析高中生命科学中物质、能量、信息转变的“桥梁”物质
2019-06-26 08:51王翔
生物学教学 2019年6期
王 翔
(华东政法大学附属中学 上海 200052)
生命体系中,一种物质转变成另一种物质,一种形式的能量转变成另一种形式的能量,一种信号转变为另一种信号,都不能凭空跨越,都需要依赖于某种“桥梁”。高中《生命科学》(沪教版)教材中出现了哪些充当桥梁作用的物质呢?它们是如何起桥梁作用的?机体又是怎样调节这些桥梁物质的?本文就这些问题作简要分析。
1 物质转变的“桥梁”物质
生命活动最基本的特征是新陈代谢,新陈代谢是通过一系列的化学反应来完成的。在化学反应中,底物需要与酶结合后才能变化为产物。我们把酶叫做生物催化剂,没有酶,底物就难以转变为产物。也就是说,酶是底物“通往”产物的“桥梁”。 1897年,Bründler兄弟证明了不含细胞的酵母提取液能使糖转化为酒精,是最早说明物质转变是酶作用的结果的研究报道,为此他们获得了1911年诺贝尔化学奖。
从底物到产物,酶是如何起作用的?首先要知道,在化学反应时,底物分子中只有活化分子才能在分子碰撞中发生化学反应,一般分子需要给以活化能成为活化分子后才能发生化学反应。酶在化学反应时,与底物分子瞬时结合形成酶—底物复合物过渡态,降低了反应所需要的活化能,导致活化分子增加,底物经化学反应形成产物,酶又恢复游离状态,重新参加反应[1]。
由于酶比非生物催化剂更显著地降低活化能,所以酶催化效率更高;一种酶只能与一种或一类底物结合,酶具有高度的专一性;酶是由细胞产生的生物大分子(蛋白质或RNA),凡是能改变生物大分子空间结构的因素,如高温、强碱、强酸、重金属盐等都能导致底物不能与酶结合,使酶失去作用。
酶的分泌与活性是受到机体调节和控制的。例如,唾液腺分泌唾液受到植物性神经调节,乳腺组织在分娩后受激素调节合成乳糖合成酶,胃蛋白酶只有进入胃腔内才能起消化作用[2],等等。
2 能量转变的“桥梁”物质
2.1 叶绿素a 在光合作用光反应过程中,某些叶绿素a分子吸收光能后能释放出高能电子(图1),这些叶绿素a分子是光能“通往”电能的“桥梁”。
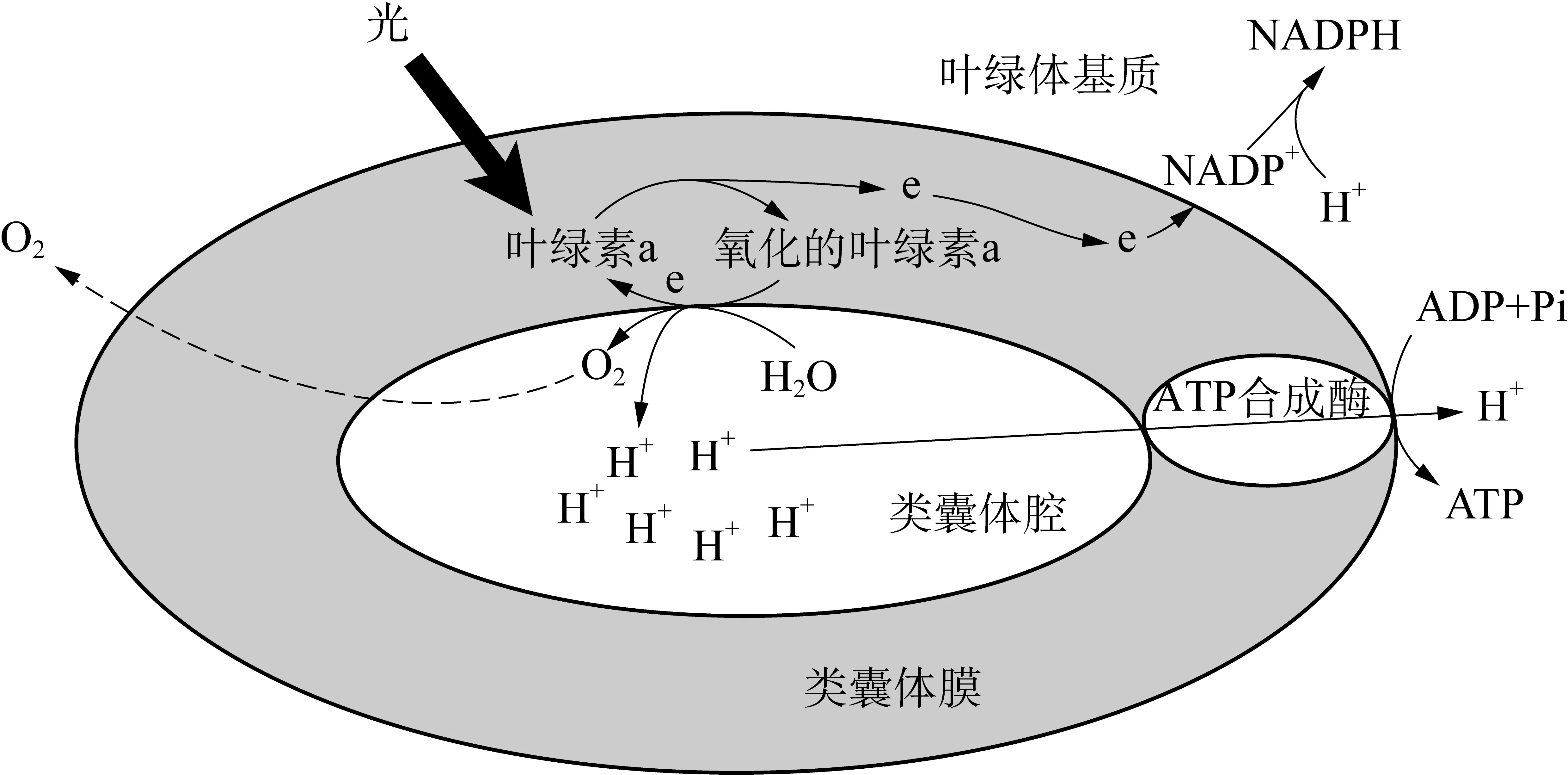
图1 光合作用光反应过程
叶绿素a分子是怎样将光能转为电能的?类囊体膜上的各种色素将吸收的光能传递给某些叶绿素a分子后,叶绿素a(P)激发而成为激发态叶绿素a(P*),释放出电子给原初电子受体(A)。于是叶绿素a分子被氧化(带正电荷,P+),原初电子受体被还原(带负电荷,A-)。氧化的叶绿素a分子从原初电子供体(D)得到电子恢复原来状态(P),原初电子供体被氧化(D+)。这样叶绿素a就不断地把电子从原初电子供体送给原初电子受体[2],完成了光能转换为电能的过程。
叶绿素a将光能转换为电能受控于电子供体和电子受体。光反应中最终电子供体是H2O,最终电子受体是NADP+。如果缺乏H2O或缺乏NADP+,那么叶绿素a无法将光能转换为电能。如果仅有电子供体H2O,而无电子受体NADP+,同样无法将光能转换为电能,这就是没有暗反应(源源不断提供NADP+)就没有光反应的原因。1937年,R. Hill把叶绿体悬浮液在无CO2条件下照光,不能进行光反应产生O2,但在悬浮液中加入电子受体(Fe3+)就能进行光反应产生O2,这就是希尔反应。
2.2 ATP合成酶 类囊体膜内外产生的质子浓度差(电化学势)能在H+通过类囊体膜上ATP合成酶时,转为活跃的化学能(ATP)(图1)。ATP合成酶是电化学势转换为活跃的化学能的“桥梁”。ATP合成酶是怎样起作用的?英国生物化学家米切尔通过研究后提出了“化学渗透学说”: 当类囊体腔内H+达到一定浓度时,意味着类囊体膜内外产生了一定的电化学势,使镶嵌在类囊体膜上的ATP合成酶活化,形成质子通道,H+沿质子通道进入膜外,释放的自由能推动ADP和Pi合成ATP[2]。化学渗透学说已得到充足的实验证据。米切尔因此于1978年获得诺贝尔奖。
ATP合成酶的活化需要类囊体腔内H+达到一定浓度。凡是能影响类囊体腔内H+浓度的因素(如H2O的光解情况、PQ接受类囊体膜外传来的质子多少)都能影响ATP合成酶的活化。
3 信息转变的“桥梁”物质
3.1 神经递质受体 神经递质(化学信号)与突触后膜或效应器细胞膜上的神经递质受体结合能将化学信号转为电信号。神经递质受体是化学信号转换为电信号的“桥梁”。神经递质受体是怎样将化学信号转换为电信号的?神经递质分子与突触后膜或效应器细胞膜上的特殊受体结合后,受体蛋白分子内部发生变构,引起与受体结构耦联一起的某些离子通道蛋白构象发生改变,离子通道由原来的关闭状态进入开放状态,允许某些离子顺浓度梯度通过,导致突触后膜或效应器细胞膜的静息电位发生改变[3],产生动作电位(如Na+内流)或更加静息(如Cl—内流、K+外排)。
神经递质受体的活动,一方面受突触前膜释放神经递质的多少与快慢调节;另一方面,由于与神经递质受体结合的神经递质很快就被突触间隙中的神经递质酶降解而失去活性,所以也受酶的调节。
3.2 tRNA tRNA一端能与mRNA配对,另一端携带一个相应的氨基酸分子,是“核酸语言”转为“蛋白质语言”的“桥梁”(图2)。
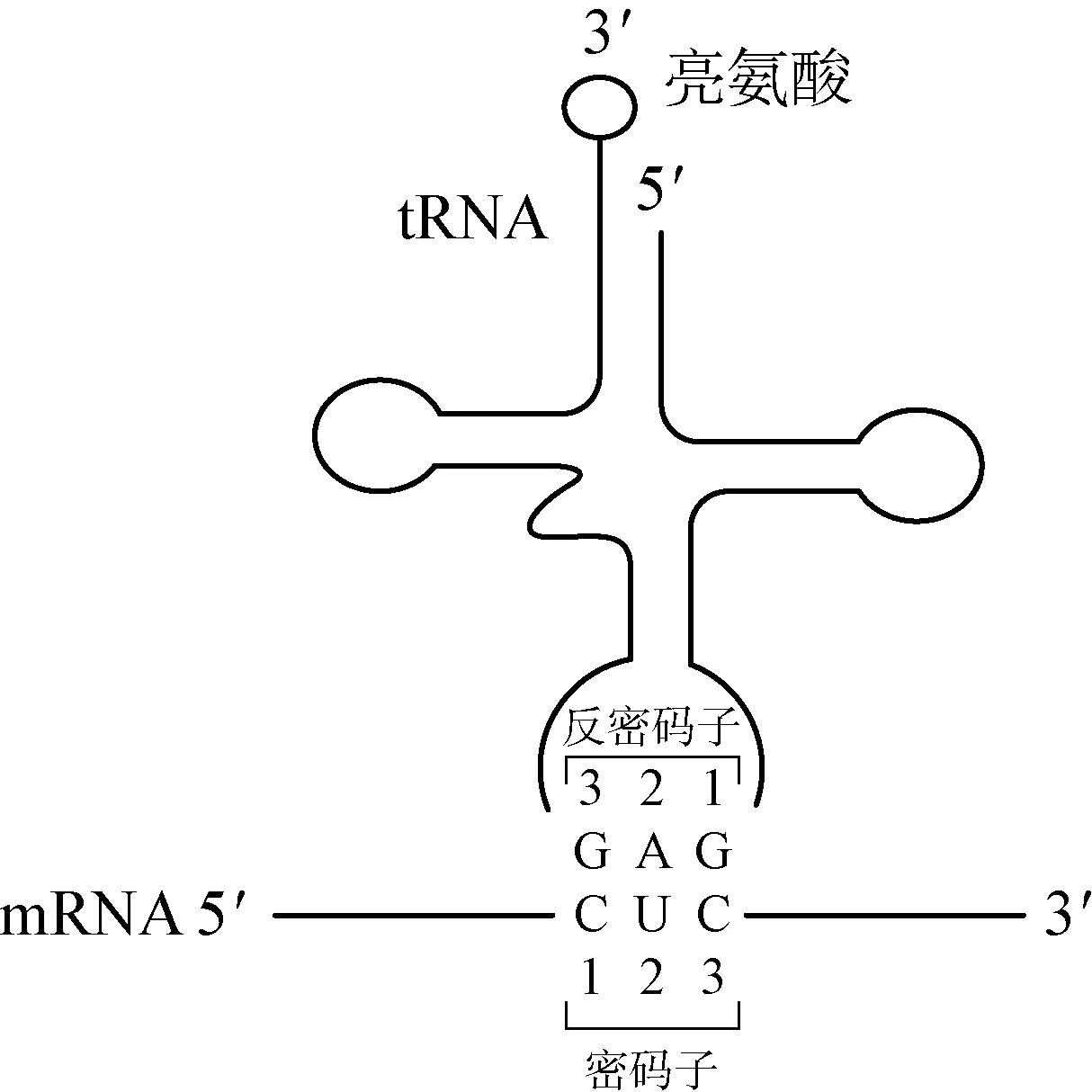
图2 tRNA模式图及氨基酸运送方式
tRNA将“核酸语言”翻译成“蛋白质语言”的原理是: 每种氨基酸都有特异的tRNA,它们之间的特异性是靠氨基酰tRNA合成酶来识别的。氨基酰tRNA合成酶具有绝对专一性,其对氨基酸、tRNA两种底物都能高度特异地识别活化。氨基酰tRNA合成酶对氨基酸和tRNA的正确选择是由两者相嵌的几何形状所决定,只有适合的氨基酸和适合的tRNA进入合成酶的相应位点,才能合成正确的氨基酰tRNA。氨基酸一旦与tRNA形成氨基酰tRNA后,进一步的去向就由tRNA来决定。tRNA分子上有核糖体识别位点和反密码子位点。tRNA进入核糖体,凭自身的反密码子与mRNA分子上的密码子相识别,把所带的氨基酸送到肽链的一定位置上。当核糖体沿mRNA模板向3′端移行A位上出现终止密码时,相关的释放因子(RF)能识别这些密码子并与之结合,水解P位上多肽链与tRNA之间的二酯键,于是新合成的多肽链和tRNA从核糖体上释放,翻译终止[1]。
tRNA的“桥梁”功能开始于氨基酰tRNA合成酶的识别,结束于释放因子(RF)对终止密码识别与结合。可见,tRNA的活动受到氨基酰tRNA合成酶、释放因子(RF)等因素的调节。
4 小结
综合上述生命体系中的5类桥梁物质,不难看出它们有以下几点共性: ①可反复使用。5类桥梁物质没有一类物质在使用后会消失或遭到破坏。②具动态性。5类桥梁物质没有一类物质是静止不变的。例如,酶与底物结合时,底物能诱导酶分子的构象发生变化,使酶分子能与底物很好地结合,从而发生催化作用;叶绿素a能激发成为激发态,激发态又能恢复到原状;ATP合成酶活化后变构形成H+通道;受体蛋白与神经递质结合后,分子内部发生变构;tRNA携带氨基酸从细胞质基质进入核糖体。③受机体调节。另外,5类桥梁物质中“ATP合成酶”与“酶”的桥梁作用有所不同,ATP合成酶既是电化学势转换为活跃的化学能的“桥梁”,又是底物ADP和Pi“通往”产物ATP的“桥梁”。
猜你喜欢
中国图书评论(2022年4期)2022-11-22
世界科学技术-中医药现代化(2022年3期)2022-08-22
材料与冶金学报(2022年2期)2022-08-10
右江医学(2022年3期)2022-04-17
科学与财富(2021年33期)2021-05-10
大众健康(2021年2期)2021-03-09
教育周报·教育论坛(2020年3期)2020-10-21
科学(2020年2期)2020-08-24
作文成功之路·小学版(2020年6期)2020-07-27
作文成功之路·小学版(2020年5期)2020-06-11
