
不同氮水平下功能叶片数量和位置对水稻产量性状的影响
2019-06-17 09:30:10田广丽孔亚丽张瑞卿周新国郭世伟
植物营养与肥料学报 2019年5期
田广丽,孔亚丽,张瑞卿,周新国,郭世伟*
(1 南京农业大学资源与环境科学学院,江苏南京 210095;2 中国农业科学院农田灌溉研究所,河南新乡 453002;3 江苏省农垦农业发展股份有限公司新曹分公司,江苏东台 224246)
由于水稻穗粒体积的变化很小,水稻的库容量主要是由穗粒数决定的[1-2]。水稻产量主要取决于单位面积总穗数和穗粒数[3-4]。通常氮肥用量、土壤水分、光照等因素显著影响穗粒数、籽粒的充实度[4-6]。当前生产系统中伴随着新品种的选育,弱势粒灌浆程度差的问题日趋严重[7-8],通过改善籽粒灌浆特性以提升单穗重量将面临更大的挑战[9-10]。相关研究表明,籽粒干物质约有30%来自于花前茎秆、叶鞘储存的光合产物,另外部分来自花后叶片新合成的光合产物[11-12],两者的贡献受环境条件、栽培措施等因素的影响,其中氮肥用量就是影响不同穗位籽粒灌浆特性的主要栽培因素之一[13]。因此采取适当的管理措施对增加水稻籽粒产量至关重要。
本研究以不同氮水平处理下成熟期的水稻为研究对象,系统的研究和分析成熟期水稻单穗重、穗长度、穗梗数、每穗粒数等具体的穗部性状;通过花后修剪叶片的方法即控制花后干物质的合成[14],从一定程度上辨别强、弱势粒物质的主要来源及来源比例随氮水平的变化趋势,旨在为氮肥的合理使用及产量的提高提供更加有力的理论指导。
1 材料与方法
1.1 试验点概况和试验材料
田间试验于2014年在江苏省如皋市农业科学研究所 (120.49°E、32.44°N) 进行,南京农业大学已在此试验点已开展了6年的长期定位试验,供试土壤为由江淮冲积物发育而成的薄层高砂土。试验田地力均匀,种植制度为稻麦轮作。耕层0—20 cm土壤的基本理化性状如下:土壤有机质12.0 g/kg、全氮1.02 g/kg、有效磷24.0 mg/kg、速效钾83.5 mg/kg、pH 7.07。供试水稻品种为适宜在江苏省沿江及苏南地区种植的早熟晚粳稻‘镇稻11号’,分别于每年5月中播种育秧,6月25日前后移栽,10月底收获。
1.2 试验设计
以氮肥用量为主区,设置5个氮肥水平:0、90、180、270和 360 kg/hm2,分别记为 N0、N90、N180、N270和N360。以种植密度为副区,设置2 个密度水平:高密度 (32.47 × 104穴/hm2,HD),行株距为 22 cm × 14 cm;低密度 (25.51 × 104穴/hm2,LD),行株距为28 cm × 14 cm。共10个处理,每个处理3个重复,随机排列,共30个小区,小区面积50 m2。每个小区均单设进、排水口,小区间田埂用防水布覆盖隔离防渗,且每个小区四周均设保护行。
1.3 试验实施方法
田间试验水分管理方式:薄水移栽,寸水活棵;苗期浅水勤灌,在分蘖数达到约300万/公顷时进行烤田;孕穗期保持浅水层,深水抽穗扬花、抽穗1周后干湿交替灌溉,水稻成熟前7~10天断水。氮肥、磷肥和钾肥分别以尿素 (含N 46%)、过磷酸钙 (含P2O514%) 和氯化钾 (含K2O 60%) 的形式施用。其中,氮肥分4次施用,即基肥∶分蘖肥∶促花肥∶保花肥 = 4∶2∶2∶2;磷肥在移栽前以基肥一次性施入;钾肥分2次施用,基肥∶促花肥为2∶1的比例施用。磷肥 (P2O5) 总用量为75 kg/hm2;钾肥 (K2O) 总用量为90 kg/hm2。在移栽的前1天施基肥,插秧后7~10天施分蘖肥,促花肥在倒四叶时施入,保花肥在倒二叶时施入。病虫草害管理根据当地水稻病虫害防治常规方法进行。
1.4 测定项目与方法
水稻于10月26—30日成熟,每小区随机调查20穴植株的穗数,选取3穴具代表性的植株用于测定穗长、穗梗数、每梗穗粒数、总穗粒数、千粒重、结实率。
采用叶片修剪法和产量差法测定不同叶片对产量的贡献。于水稻开花齐穗期 (9月4日),在每个小区选择单茎株高、穗长、穗抽出长度等长势基本一致的单茎穗200个,挂上标签,分别从叶基部整体修剪掉倒1叶 (-T1)、倒2叶 (-T2)、倒3叶 (-T3)、倒1+2叶 (-T1 + 2)、倒1+3叶 (-T1 + 3)、倒2 + 3叶(-T2 + 3)、倒1 + 2 + 3叶 (-T1 + 2 + 3) 和不修剪(-T0,对照) 各20株,组成8个叶位修剪处理。于10月28日 (收获期) 将每小区各叶位修剪处理穗子分别取下,放置烘箱,测定穗重,分别记录千粒重。
1.5 数据处理与分析
采用SigmaPlot 12.5、SPSS 16.0、Microsoft Office Excel 2010、R 2.15.1等软件进行统计分析并作图,所有数据运用LSD法进行方差分析。
2 结果与分析
2.1 氮水平对水稻穗部性状的影响
单穗重随氮水平的增加呈增加趋势,但是5个氮水平间均无显着差异 (图1A);N0时穗长17.2 cm与其它氮水平下穗长19.8 cm有显著差异之外,其它四个氮水平下的穗长均无显着差异 (图1B),同时,随氮水平的增加,穗梗数的变化趋势与穗长的变化具有相似现象 (图1C),在N0与其它四个氮水平下的平均穗梗数分别为11.8、12.2个/穗。通过对低 (N90)、中 (N180)、高 (N270) 三个氮水平下不同穗梗穗粒数的统计结果 (图2) 表明,从穗顶端至穗基部随梗位的增加,梗粒数均呈先增加后下降的趋势。并且在试验中的两个移栽密度下上述穗部性状随氮水平的变化趋势均表现一致,且相同氮水平下同一性状在两个密度间无显著差异,因而表明氮水平对穗部性状影响的稳定性。
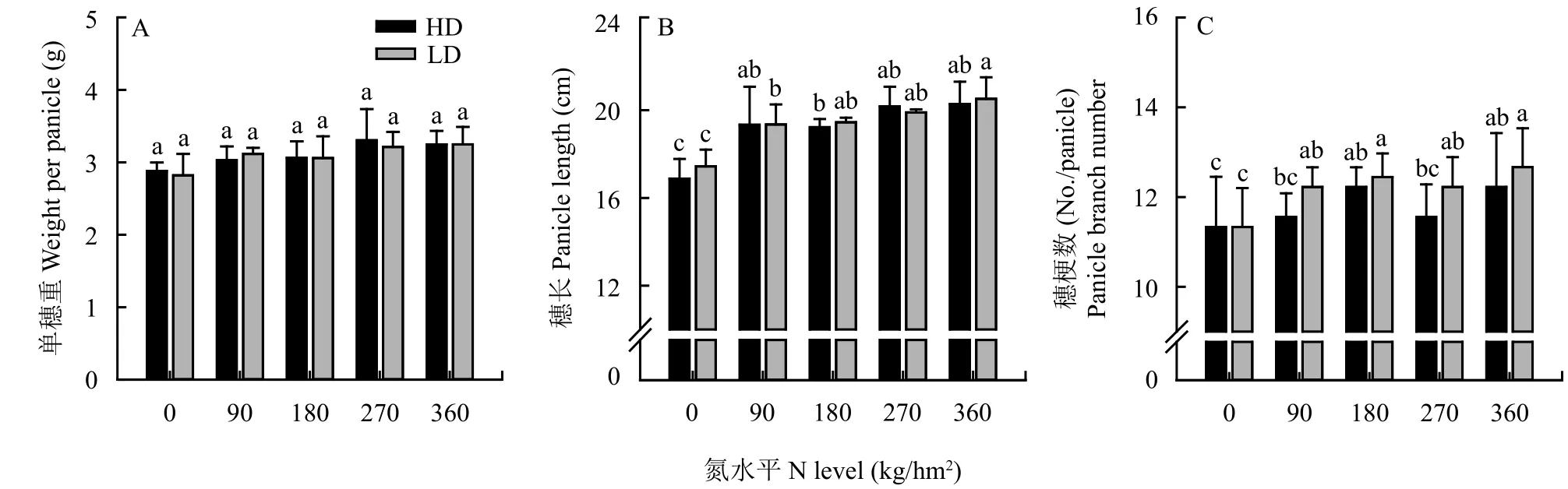
图 1 氮水平对水稻成熟期单穗重、穗长、穗梗数的影响Fig. 1 The effect of N level on the weight per panicle, panicle length, and panicle branche number at maturity stage of rice

图 2 氮水平对水稻不同梗位粒数的影响Fig. 2 The effect of N level on rice kernel number at different branches
2.2 不同氮水平下水稻穗粒产量构成参数
氮水平对水稻穗粒产量构成参数的影响表现为水稻单位面积的穗数对氮水平的响应最大,在不同氮水平间的平均变异系数为90.7%,然后是穗粒数和千粒重,它们在不同氮水平间的变异系数分别为22.5%、20.0%。此外,氮水平响应最小的是结实率,其在不同氮水平间的变异系数为1.3%。多重比较分析结果表明,氮水平对所有产量构成参数影响显著,且氮水平对所有的产量构成参数的影响在两种栽培密度下表现基本一致(表1)。
2.3 不同叶位叶片对单穗重的影响
图3A表明, 单独剪去倒1、倒2、倒3叶中任意一片叶时对单穗重没有显著影响;剪去倒1、倒2、倒3叶中任意两片叶时单穗重显著下降,当剪去倒1、2叶时单穗重比正常单穗重下降5.5%、当剪去倒1、3叶时单穗重下降10.3%、当剪去倒2、3叶时单穗重下降8.3%;剪去倒1、2、3三片叶时,单穗重下降24.6%。不同修剪叶片处理对单穗重的影响在两个栽培密度下呈现一致趋势。
将不同氮水平、栽培密度、不同叶位叶片对单穗重的影响进行统一分析 (图3B) 表明,当剪去一片叶时,N90、N180、N270氮水平下单穗重下降比例依次为0.4%、1.6%、5.8%;当剪去两片叶时下降比例依次为4.7%、8.1%、14.5%;当上三片叶都剪去时下降比例依次为19.0%、26.5%、28.2%。
2.4 不同叶位叶片对结实率和千粒重的影响
图4A、4B、4C均表明,各剪叶片处理对穗粒参数的影响在两种栽培密度下呈现一致的趋势。因此以下分析均以两栽培密度下的均值进行。由图4A可知,各氮水平下单独剪去倒1、倒2、倒3叶中1片叶时,结实率下降6.9%,剪去倒1、倒2、倒3当中任意两片叶时,下降18.4%,将倒1、倒2、倒3叶三片叶全剪去时,下降27.3%。图4D表明,各种叶片修剪方法对结实率的影响均随氮水平的增加而增加,在N90、N180、N270氮水平下,单独减去1片叶,结实率分别下降5.3%、6.3%、9.2%;剪去任意两片叶,分别下降14.7%、16.9%、23.7%;三片叶全剪去时,结实率分别下降26.0%、26.1%、29.8%。
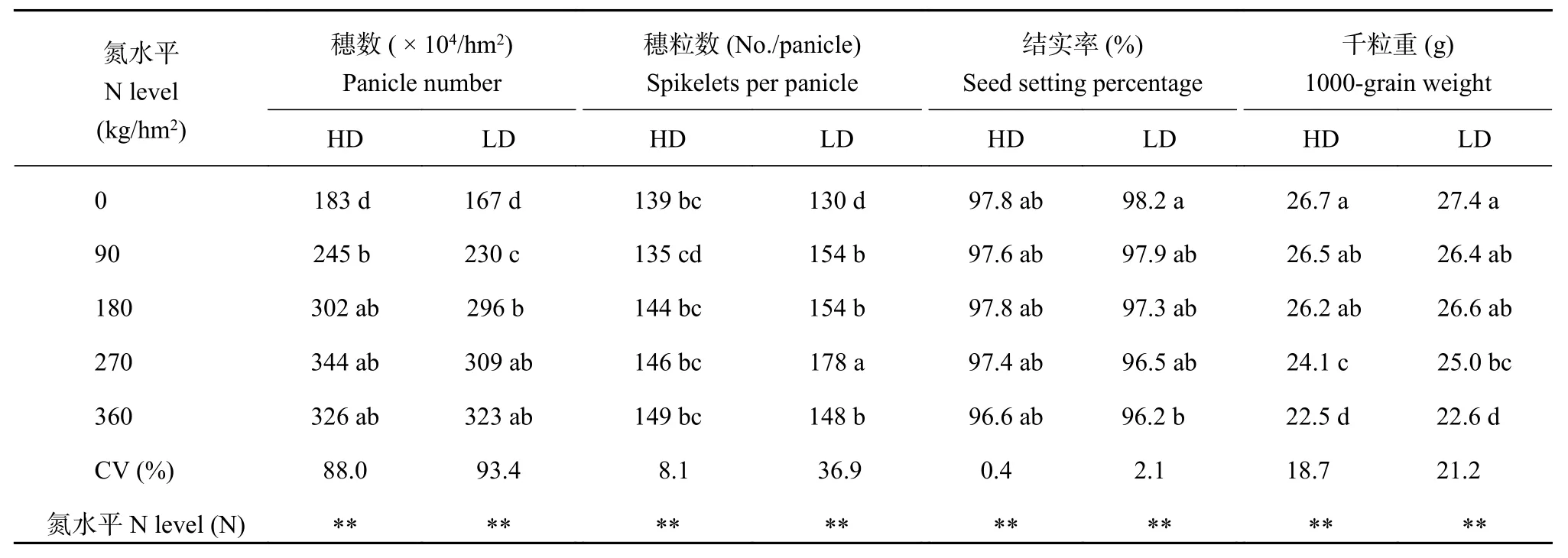
表 1 不同氮水平下水稻穗粒产量构成参数Table 1 The components of rice grain yield under different N levels
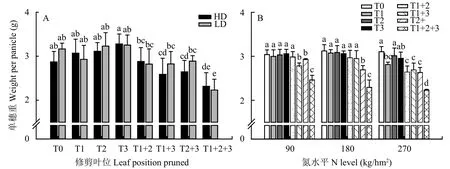
图 3 不同叶位叶片对水稻成熟期单穗重的影响Fig. 3 Effect of different position leaves on weight per panicle of rice at maturity stage
由于水稻强、弱势粒灌浆的异步性,因此其物质来源可能不同,可能来源于抽穗前营养器官储存的非结构性碳水化合物或者抽穗后有效绿色叶片新合成的光合产物,因而叶片的修剪会对强、弱势粒的影响程度存在差异。图4B表明,各氮水平下单独剪去1片叶时,强势粒千粒重下降比例平均2.2%,剪去任意两片叶时下降比例为7.3%,三片叶全剪去时下降比例为12.5%;图4C表明,各氮水平下单独剪去1片叶时弱势粒千粒重下降比例的平均值为7.8%,剪去任意两片叶时千粒重下降比例的平均值为47.5%,三片叶全剪去时千粒重下降比例的平均值为73.5%。同时,图4E、F表明,各种叶片修剪方法对强、弱势粒千粒重的影响均随氮水平的增加而增加,且各种叶片修剪处理下强、弱势粒千粒重的下降比例差异显著,在N90、N180、N270氮水平下具体表现为:单独剪去1片叶时,强势粒千粒重分别下降0.7%、4.5%、1.4%,弱势粒千粒重分别下降5.3%、9.4%、10.2%;剪去任意两片叶时,强势粒千粒重分别下降3.0%、11.2%、7.7%,弱势粒千粒重分别下降49.5%、53.7%、31.7%;三片叶全剪去时,强势粒千粒重分别下降5.4%、15.8%、16.6%,弱势粒千粒重分别下降73.6%、78.2%、64.5%。
2.5 穗粒参数对单穗重变化的解释
在所有解释单穗重变化的广义线性模型 (表2)中,结实率 (SP) 的模型性能优于其他穗粒参数模型,可单独解释大于78%的单穗重变异率;随后的模型性能依次为SP和强势粒千粒重 (S) 组合模型 > SP和弱势粒千粒重 (I) 组合模型 > SP、S、I三者组合模型 >I、S组合模型。而I在逐步分析中已经被默认不是最好的解释变量而未单独出现在分析结果中。
3 讨论
水稻作为全世界人们主要的粮食作物之一[15-16],其产量备受关注。稻穗作为最大的库器官,是水稻育种和提高产量的首要考虑对象,在大多数谷类作物中,库容一般由每株穗数、每穗粒数、灌浆效率三方面决定[17]。对于某特定水稻品种,其每株穗数即个体的分蘖能力由氮水平、种植方式、栽培密度等外界因素决定[18-20]。而每穗粒数与稻穗的次生枝梗、次生枝梗上幼穗的分化有密切的关系[21];此外,当穗粒数一定时,穗粒的重量、结实率均是影响穗部产量的主要因素。
3.1 氮水平对水稻穗部性状和产量构成参数的影响
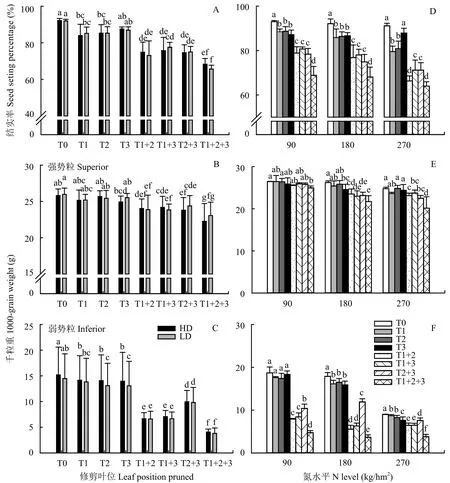
图 4 不同叶位叶片对水稻结实率和强、弱势粒千粒重的影响Fig. 4 The effect of different position leaves on seed setting percentage, and 1000-grain weight of superior and inferior spikelets of rice
本研究表明,随氮水平增加,两栽培密度下的稻穗长度均呈先增加然后趋于平稳甚至降低的趋势。因为穗分化的同时,茎秆处于伸长阶段,即穗与茎秆间存在同伸的关系,因此在茎秆节间的生长随氮水平增加而增加的同时,稻穗长度也在增加[22];穗梗数随氮水平的变化呈类似趋势,一方面,因为氮水平影响着控制稻穗长的数量性状基因的表达[23],氮肥的施用增加了稻穗一次支梗的分化数,因而在此基础上增加了后期的二次支梗数,但稻穗颖花的成活率与单茎干物质的积累成正相关关系[22]。相关研究表明,水稻单茎干物质形成颖花的能力随氮水平的增加而下降[24],同时,过多氮肥的施用在增加水稻颖花分化数的同时也导致了一次枝梗颖花退化量的增加,而氮浓度过低会引起二次枝梗颖花的退化,这是由于幼穗发育的同时还伴随着节间伸长及新生叶片的生长;且植株氮浓度过高或过低时,植株碳氮代谢紊乱,茎、叶过度生长造成群体密闭、光合源减弱[23,25-27]。然而有效穗粒数是颖花分化数与退化数之差,因此,随氮水平的增加穗粒数的增加有一定限度,呈现先增加后下降的趋势。
同时,本研究结果表明,各施氮水平下成熟期单穗重无显著差异。原因在于,随氮水平的增加,水稻粒叶比下降,源库比失调,单位叶面积所承受的颖花数、实粒数、粒重减少[28],最终造成水稻产量构成因素之间的矛盾逐渐增加。表现为随氮水平增加,结实率和千粒重下降程度对单穗重的贡献大于穗粒数增加程度对单穗重的贡献。
3.2 不同叶位叶片对穗粒参数的影响
花后叶片的剪除结果表明,随剪除叶片数的增加,单穗重及穗粒结构参数降幅增加,且降幅随氮水平的增加而增大。因为在穗分化期穗、茎同步迅速生长,穗与茎的相对库强度受源的限制。随氮水平增加,源的限制减小,穗茎比增加,即在穗分化后期较多光合产物输送到了穗部,而分配到茎秆的干物质比例减少。这导致花前储存于营养器官的物质相对减少,明显降低穗粒的贡献率[22],进而使得稻穗在灌浆期更大程度的依赖花后新合成的物质,因而造成花前营养器官储存的干物质和花后新合成的干物质对穗粒充实过程的贡献程度因氮水平不同而不同。因此,随氮水平的提高,伴随叶片剪除数量的增加,稻穗成熟率、粒重均降低[27,29-30]。
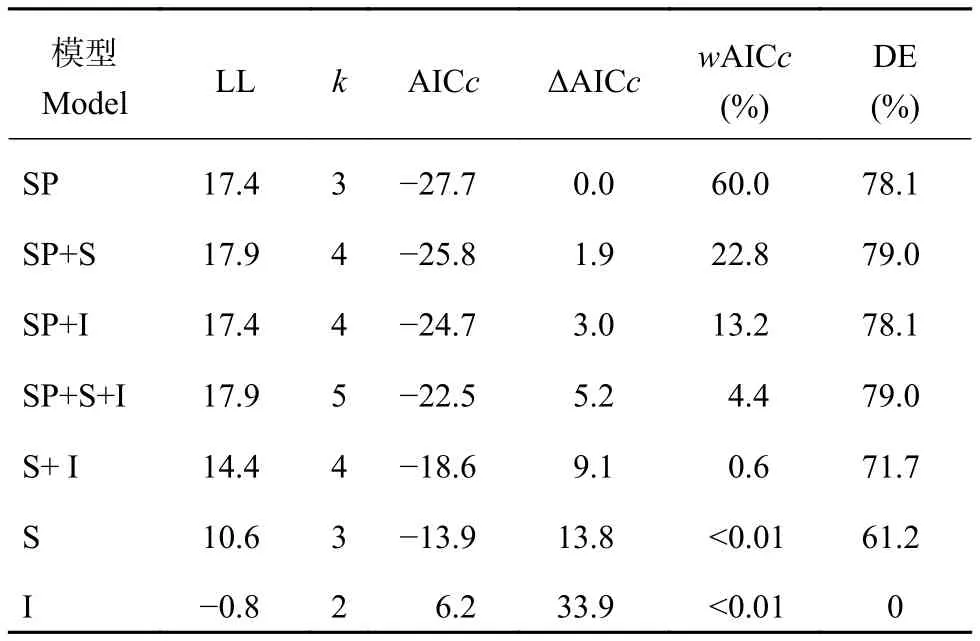
表 2 穗粒参数对单穗重变化的解释Table 2 The interpretation of kernels parameters to the change of weight per panicle
单独修剪倒1、倒2、倒3叶时,单穗重无明显变化,与郭九信[14]的研究结果相似。从一定程度上说明上三片功能叶对穗粒的贡献基本一致,同时也说明上三叶中,任一片叶的缺失不会影响灌浆期穗粒干物质的供给,表明花后剪去部分功能叶后,其余叶片通风透光条件得到改善,可利用的光辐射增加,弥补因叶面积减少而造成的损失,降低了功能叶缺失而致的负面效应。这一现象与王勤等[31]的研究结果相矛盾,比较表明主要是因为叶片修剪时间的不同,王勤等的研究是在孕穗期旗叶刚完全长出时便进行修剪,使得上三片功能叶对穗粒的贡献依次为倒1 > 倒2 > 倒3;此外,试验所用品种、肥料的施用量与方式的不同,均可导致功能叶对穗粒的贡献程度差异。
当修剪任意两片功能叶时,结实率和弱势粒千粒重均显著下降,造成单穗重明显下降,且任意两片叶对单穗重的贡献相似,说明当两片功能叶被剪除时,有效叶面积的缺失不能被第三片叶光能利用效率的增加所弥补。当上三叶全被剪除时,结实率和千粒重的下降幅度显著大于剪除两片叶时的下降幅度,且大于单独剪除一片功能叶和任意两片功能叶的下降总和。说明上三片功能叶对穗粒的贡献之间具有一定的相互依赖关系,而不是完全独立的。当上三片功能叶全部剪除时,源库比严重失调,严重影响穗粒的灌浆过程,瘪粒增多且粒重下降,最终造成单穗重显著下降[32]。
此外,由于试验品种属于强、弱势粒异步灌浆型,其弱势粒中启动籽粒灌浆的内源激素IAA、Z+ZR及ATP含量显著低于强势粒[3],因此各处理下弱势粒的灌浆均在其强势粒灌浆基本结束时才开始,弱势粒进入活跃灌浆期时已经是灌浆后期,叶片光合机能开始下降,光合产物对弱势粒的供应逐渐减少,因此弱势粒参数受花后有效绿叶面积的影响大于强势粒。
4 结论
1) 与不施氮相比,随氮水平增加稻穗长度与穗梗数均呈增加趋势,但两者在氮水平间的差异不显著;梗粒数随梗位的增加呈先增加后降低趋势,基本不受氮水平的影响;结实率和千粒重逐步下降。因而,单穗重在各氮水平间无显著差异。
2) 随花后叶片剪除数量的增加,粒重的下降比例增加,其中弱势粒粒重的下降比例大于强势粒,且下降比例随氮水平的增加而增加,即随氮水平增加花后新合成光合产物对籽粒的贡献率增加,且弱势粒物质更多来源于花后新合成的光合产物。
3) 当剪除叶片数量一定时,剪除叶片的位置对单穗重的影响程度无明显差异,并且花后上三片功能叶依次通过影响SP、SP和S、SP和I、I和S来影响稻穗的重量。
猜你喜欢
西北农业学报(2022年9期)2022-10-08 11:07:18
西南农业学报(2022年1期)2022-03-15 12:51:16
热带作物学报(2021年9期)2021-11-08 11:55:35
核农学报(2021年3期)2021-02-22 08:47:58
疯狂英语·初中天地(2020年8期)2020-09-11 07:43:38
中国化肥信息(2020年7期)2020-03-19 01:54:06
北京农学院学报(2020年1期)2020-03-09 07:18:28
中国化肥信息(2019年7期)2019-08-26 09:46:54
茶叶科学(2018年5期)2018-10-17 01:56:42
中国稻米(2018年1期)2018-03-20 06:30:01
