
拟南芥MDH2基因GFP载体构建及转基因植株的筛选鉴定
2019-06-11 05:31王媛媛韩洋洋耿庆鎏朱香豫曹树青樊婷婷
安徽农业科学 2019年2期
王媛媛 韩洋洋 耿庆鎏 朱香豫 曹树青 樊婷婷

摘要[目的]以野生型拟南芥为材料构建MDH2基因GFP载体及GFP转基因植株,研究MDH2基因在植物响应镉胁迫机制中的功能。[方法]通过提取野生型拟南芥的RNA,反转录成为cDNA,并以cDNA为模板,利用PCR扩增MDH2基因,将扩增所获得MDH2基因和pXB94-GFP质粒双酶切后连接,并转化进大肠杆菌感受态细胞中,鉴定正确后再转入农杆菌感受态细胞中,通过菌落PCR鉴定出阳性单菌落。再通过浸花法转入野生型拟南芥中,最后利用抗性筛选和PCR鉴定获得MDH2转基因阳性植株。[结果] 成功克隆MDH2基因,并构建了MDH2GFP重组质粒,抗性筛选获得了MDH2GFP转基因植株。[结论]成功获得MDH2GFP转基因植株,为进一步研究该基因在植物响应镉胁迫机制中的功能奠定了基础。
关键词拟南芥;MDH2;载体;转基因植株
中图分类号Q939.9文献标识码A
文章编号0517-6611(2019)02-0096-03
doi:10.3969/j.issn.0517-6611.2019.02.028
土壤重金属污染已成为世界性的环境问题之一,其中重金属镉污染是对动植物具有高毒害性的污染物之一,过量的镉会对动植物细胞造成不可逆的损伤[1-4]。植物对重金属胁迫的响应机制主要有控制金属流入、促进金属泵出、重金属螯合等途径[5-7]。利用植物转基因技术对植物品种进行优化改良是解决上述问题的一种行之有效的方法 [8]。前期研究发现,MDH2功能缺失突变体对重金属镉胁迫表现出耐受的表型,因此笔者拟通过克隆MDH2基因,构建MDH2过表达植株,以进一步探究MDH2基因在拟南芥响应镉胁迫机制中的作用。
1材料与方法
1.1材料
1.1.1材料。该试验所用植物材料为拟南芥(Arabidopsis thaliana)哥伦比亚(Columbia,col)遗传背景,购于美国拟南芥种质资源中心,后由该实验室(合肥工业大学植物分子生物学实验室)繁殖所得。载体构建所用质粒pXB94-GFP、大肠杆菌DH5α、农杆菌GV3101均购自北京全式金生物技术有限公司。
1.1.2主要试剂。Plasmid miniprep Kit、Easy Taq DNA Polymerase、DNA loading buffer均购自北京全式金生物技术有限公司;T4-DNA Ligase、限制性内切酶Xho I、Eco RI购自NEB公司;PrimeSTAR HS DNA Polymerase购自TaKaRa公司;Agar、 NaCl、异丙醇、氯仿、无水乙醇、葡萄糖、蔗糖购自国药集团;壮观霉素、庆大霉素、卡那霉素生工生物工程(上海)股份有限公司;Goldview、Silwet L-77购自索莱宝公司。
1.2方法
1.2.1拟南芥无菌苗培养。配制1/2 MS固体培养基,从冰箱取出1/2MS培养基放置至室温,称取所需要的组分放入三角瓶中,加入一定量的蒸馏水,用5 M/L NaOH溶液调节培养基pH至5.8,用封口膜封好后高压蒸汽灭菌,待培养基冷却至60 ℃左右,将培养基均匀倒入培养皿中,待培养基凝固后,吹干表面水分即可播种。将消毒清洗后的拟南芥种子于滤纸上晾干,种子晾干后按试验需要用牙签蘸取均匀点在培养皿上,待培养皿播种完成后,用封口膜将培养皿封好;将播种完成后的培养皿倒置于4 ℃冰箱中春化3 d,取出后放置于专用培养室中恒温(21 ℃左右)光照竖直培养14 d。定期观察拟南芥生长情况。
1.2.2拟南芥总RNA的提取。提前一天将所需研钵清洗干净,放入烘箱烘干后用锡纸包裹起来,置于180 ℃的高温烘箱中,烘烤至少4 h;在试验开始前,要提前将所用试剂如氯仿、无水乙醇、异丙醇等高温烘烤后的研钵放入-20 ℃冰箱中预冷。
用镊子轻轻从培养皿中挑取拟南芥幼苗100 mg左右,用滤纸吸干水分,置于预冷的研钵中,加入适量液氮进行充分研磨。向充分研磨后的材料中加入Trizol裂解液1 mL,立即快递研磨至全部融化;转移到1.5 mL的离心管中,于室温中静置5 min,让样品充分裂解。在超净台中向每个样品中加入200 μL氯仿,上下剧烈摇晃15 s混匀样品,再于室温下静置5 min后使用冷冻离心机4 ℃离心15 min,转速为13 000 r/min。此时可见液体分层,小心吸取500 μL上清液至另一新的1.5 mL EP管中;再向该EP管中加入500 μL预冷后的异丙醇,轻轻颠倒混匀6~8次,于室温下静置10 min;使用冷冻离心机4 ℃离心15 min,转速为13 000 r/min,小心弃除上清,留取沉淀,沉淀即为RNA。缓慢加入事先用DEPC水配制的75%乙醇1 mL清洗沉淀;冷冻离心机8 000 r/min,4 ℃离心5 min,再次弃去上清,于超凈台室温干燥10 min,使乙醇挥发干净;向离心管中加入40 μL的DEPC水溶解,并放入65 ℃的水浴中10 min,使RNA沉淀充分溶解,溶解后吸出少量立即检测其浓度与纯度。
1.2.3拟南芥cDNA的反转录。利用反转录试剂盒,将浓度及纯度符合标准的野生型拟南芥总RNA溶液反转录为cDNA,用作基因克隆的模板。
1.2.4MDH2基因的克隆。利用Oligo 7.0软件设计以下引物进行MDH2基因的克隆。上游引物为FP:5′-CCGCTCGAGATGTTCCGATCAATGAT-3′,下游引物为RP:5′-CCGGAATTCTTGGTTGGCAAATTTGA-3′,以cDNA为模板进行基因克隆。
1.2.5大肠杆菌的转化。从-80 ℃冰箱中取出大肠杆菌DH5α感受态细胞,迅速置于冰上解冻。解冻后,取5 μL质粒获连接产物加入到感受态细胞中,轻柔吹打混匀后,冰浴30 min。之后42 ℃热击60 s,迅速置于冰上2 min。随后加入到750 mL无菌未加抗性的LB液体培养基中,放在37 ℃摇床中低速震荡培养1 h。6 000 r/min离心10 min,留200 μL涂布于LB+壮观霉素的平板上。平板倒置放在37 ℃培养箱中培养过夜。
在过夜培养的培养皿上挑取单克隆菌株接种于LB+壮观霉素的液体培养基中,37 ℃震荡培养至培养基浑浊即可进行菌液PCR鉴定。
1.2.6农杆菌的转化。从-80 ℃冰箱中取出农杆菌GV3101感受态细胞,迅速置于冰上解冻。解冻后,取5 μL质粒加入到感受态细胞中,轻柔吹打混匀后,加入到预冷的0.1㎝电击杯中,调节电脉冲、电压和电阻数值,迅速将电击后感受态细胞加入到750 mL无菌未加抗性的LB液体培养基中,放在28 ℃摇床中低速震荡培养1 h。6 000 r/min离心10 min,弃去多余培养基,留200 μL涂布于LB+壮观霉素的平板上。平板倒置放在28 ℃培养箱中培养2 d。
在培养后的培养皿上挑取单克隆菌株接种于LB+壮观霉素的液体培养基中,28 ℃震荡培养至培养基浑浊即可进行菌液PCR鉴定是否为所需陽性菌株。
1.2.7花序浸染法获取转基因拟南芥。将已确定的含有表达载体的阳性农杆菌接种于含壮观霉素的LB培养液中,震荡培养至OD600=1.2~1.6,低速离心后收集菌株,重悬于配制好的浸染缓冲液中,调整溶液OD600=0.8~1.0,最后加入一定量的SilwettL-77混匀,进行花序浸染。浸染完成后黑暗处理24 h。隔7 d,再次进行浸染。
1.2.8转基因阳性植株鉴定。将获得的拟南芥种子置于含有卡那霉素的MS固体培养基中进行抗性筛选,长出具有根且子叶颜色嫩绿的幼苗,即为转基因阳性植株。
2结果与分析
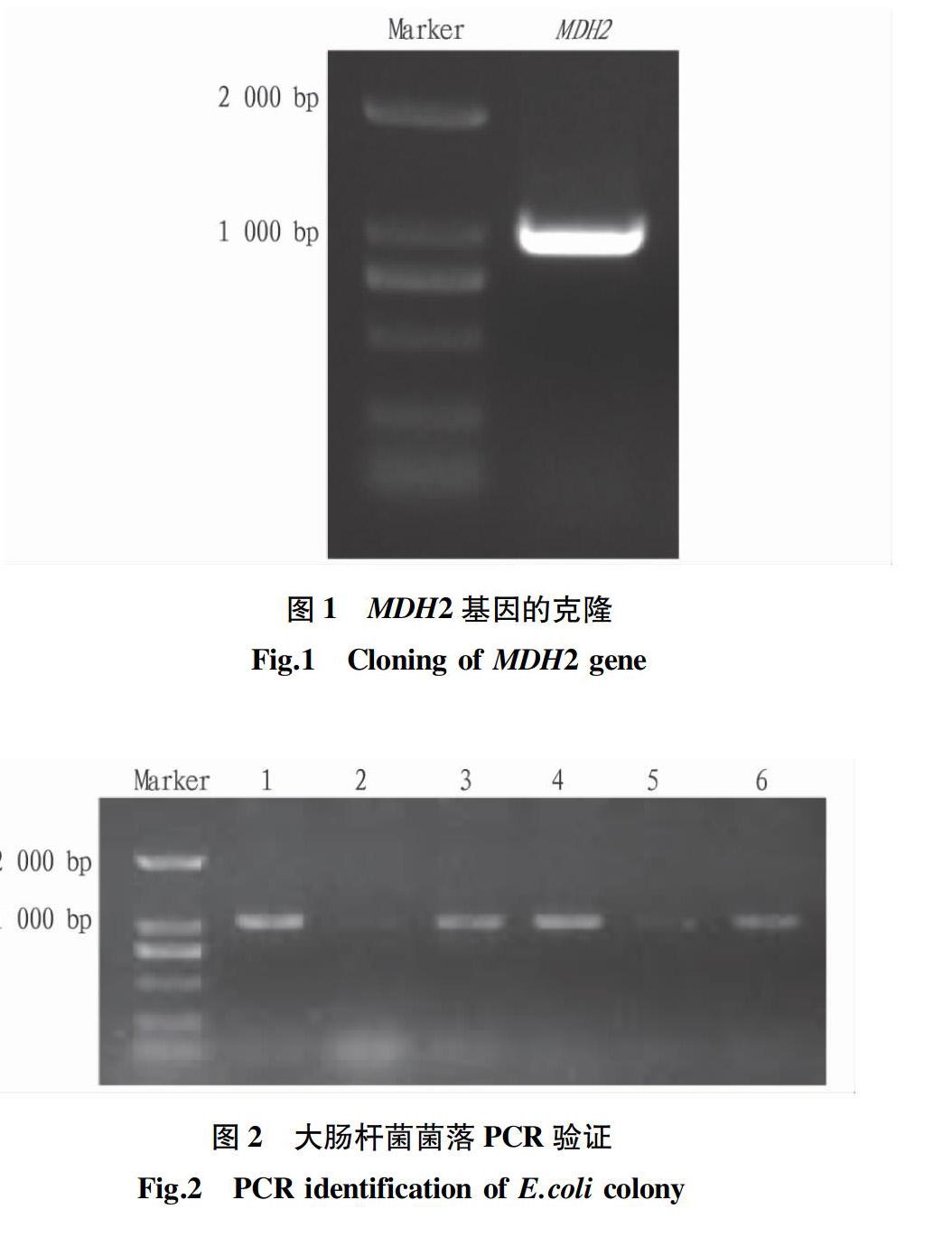
2.1拟南芥MDH2基因的克隆为验证MDH2基因在拟南芥镉耐受机理中的作用,构建MDH2过表达载体。以植物总RNA反转所得的cDNA为模板,通过PCR技术扩增MDH2基因,琼脂糖凝胶电泳检测结果如图1所示,PCR所得片段大小为1 026 bp,与拟南芥网站显示MDH2基因CDS全长结果一致。
2.2过表达载体的连接和转化基因扩增引物设计时,在上下游引物分别添加了限制性内切酶XhoⅠ和EcoRI的酶切位点保护碱基,PCR扩增所得片段可使用限制性内切酶XhoⅠ和EcoRI对通过基因克隆获得的MDH2基因和pXB94-GFP质粒同时进行双酶切。酶切以后进行电泳检测,酶切后的基因片段和载体质粒条带清晰,大小正确。
将酶切后的基因片段与质粒片段用T4连接酶进行连接,连接产物采取热击法导入大肠杆菌DH5α感受态细胞中,培养过夜后挑取单克隆菌落于加有相应抗性的液体培养基中震荡培养至菌液浑浊。
对上一步所得浑浊单克隆菌液进行菌落PCR鉴定,所用引物为MDH2片段扩增引物,结果如图2所示。除了2号与5号菌落,其他菌落PCR条带与MDH2基因大小一致,说明该条带为阳性克隆。选取1号菌液进行测序,结果与MDH2 CDS序列完全吻合,说明成功构建MDH2-GFP载体。
2.3农杆菌转化及花序浸染将测序正确的MDH2-GFP重组质粒通过电击法转化入农杆菌GV3101,接种于含庆大霉素和壮观霉素的培养基上,待其长出单菌落后进行菌落PCR验证,结果如图3所示,所选单克隆菌株除1号菌株外均为阳性单克隆。将转化成功的农杆菌进行振荡培养后,对正处于开花期的野生型拟南芥使用花序浸染法进行转基因。
2.4 MDH2-GFP转基因植株的抗性筛选获取浸染后的拟南芥种子,将获得的种子干燥春化后播种在含有卡那霉素抗性的MS平板上,放置4 ℃ 3 d后置于恒温光照培养箱中培养7~10 d,长出来的小苗(具有根,子叶颜色嫩绿)就是转基因阳性植株,如图4所示。
2.5MDH2-GFP转基因阳性植株的验证将MDH2-GFP转基因阳性植株移栽至土壤中,置于专用培养室中恒温光照培养,20 d后提取转基因植株DNA,进行鉴定。PCR鉴定结果如图5所示,筛选所得植株均为转基因阳性植株,即MDH2-GFP转基因植株。
3讨论
重金属污染对环境及生物的危害严重,尤其是重金属可以通过食物链富集和污染水资源,对人体健康和环境安全造成极大的威胁[9]。近年来,植物修复的出现和迅速发展为土壤重金属污染提供了一条新的治理途径。相对于常规的污染土壤治理方法,植物修复以其廉价、清洁、易操作的优点而成为最有潜力的技术之一[10]。
笔者自拟南芥种子资源中心获得MDH2基因功能缺失型突变体,在前期研究结果显示MDH2基因参与了植物对镉胁迫的响应,因此通过基因工程技术构建MDH2-GFP重组载体,并通过生物学手段将其转入野生型拟南芥中,从而获得转基因植株。这对研究MDH2基因在植物镉耐受调节机制中的作用奠定了基础,是一项非常有意义的研究性课题。
参考文献
[1] KIM D Y,BOVET L,MAESHIMA M,et al.The ABC transporter AtPDR8 is a cadmium extrusion pump conferring heavy metal resistance[J].Plant J,2007,50(2):207-218.
[2] SHIM D,HWANG J U,LEE J,et al.Orthologs of the class A4 heat shock transcription factor HsfA4a confer cadmium tolerance in wheat and rice[J].The plant cell,2009,21(12):4031-4043.
[3] GALLEGO S M,PENA L B,BARCIA R A,et al.Unravelling cadmium toxicity and tolerance in plants:Insight into regulatory mechanisms[J].Environ Exp Bot,2012,83:33-46.
[4] STRAIF K,BENBRAHIMTALLAA L,BAAN R,et al.A review of human carcinogens-Part C:Metals,arsenic,dusts,and fibres[J].Lancet Oncol,2009,10(5):453-454.
[5] KIM D Y,BOVET L,KUSHNIR S,et al.AtATM3 is involved in heavy metal resistance in Arabidopsis[J].Plant Physiol,2006,140(3):922-932.
[6] HALL J L.Cellular mechanisms for heavy metal detoxification and tolerance[J].J Exp Bot,2002,53:1-11.
[7] CLEMENS S.Molecular mechanisms of plant metal tolerance and homeostasis[J].Planta,2001,212(4):475-486.
[8] BELHAJ K,CHAPARROGARCIA A,KAMOUN S,et al.Plant genome editing made easy:Targeted mutagenesis in model and crop plants using the CRISPR/Cas system[J].Plant methods,2013,9(1):1-10.
[9] NAWROT T,PLUSQUIN M,HOGERVORET J,et al.Environmental exposure to cadmium and risk of cancer:A prospective population-based study [J].Lancet Oncol,2006,7(2):119-126.
[10] LU L L,TIAN S K,YANG X E,et a1.Cadmium uptake and xylem loading are active processes in the hyperaccumulator Sedum alfredii[J].Joumal of plant physiology,2009,166(6):579-587.
猜你喜欢
——宣教载体
道路交通管理(2022年12期)2022-12-30
华人时刊(2022年9期)2022-09-06
华人时刊(2020年15期)2020-12-14
国际放射医学核医学杂志(2020年4期)2020-07-27
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年3期)2017-04-10
山东农业工程学院学报(2016年6期)2016-12-01
广州大学学报(自然科学版)(2015年4期)2015-12-23
华东师范大学学报(自然科学版)(2014年3期)2014-03-11
中国火炬(2013年11期)2013-07-25
