
羰基还原酶基因工程菌产酶和不对称合成(R)-苯乙醇的条件优化
2019-06-11 11:35郝军莉章英杨天兵王婷杨宇王丹
中国测试 2019年4期
郝军莉 章英 杨天兵 王婷 杨宇 王丹

摘要:以羰基还原酶基因工程菌全细胞为催化剂,探讨不对称合成(R)-苯乙醇的各种影响因素,优化其产酶和转化条件。结果表明,优化后的发酵培养基为甘油10g/l、蛋白胨10g/L、酵母粉5g/L(NH4)2SO45g/L、NaCl l0g/L,氨苄青霉素50ug/mL,pH 7.2;诱导剂IPTG的浓度为0.25mmol几,诱导时间为20h,诱导温度为18℃,此条件下测得基因工程菌粗酶活最高为6.65u/mL。建立的最佳转化条件:羰基还原酶基因工程菌发酵20h后的细胞浓度为03g/mL,转化体系初始pH为7.0,温度为37℃,辅助底物异丙醇浓度为10%,底物终浓度为60mmol/L,转化时间为24h。此时底物转化率最高可达98.88%,产物对映体过量值(e.e、值)为99.43%。反应体系扩增至3000mL后,产物e.e.值仍保持在99.0%左右,底物转化率可为92%以上,(R)-苯乙醇的产量也可达到6.67g/L。
关键词:羰基还原酶;基因工程菌;不对称合成;产酶条件;转化条件
中图分类号:Q936文献标志码:A 文章编号:1674-5124(2019)-04-0073-07
0引言
手性仲醇作为合成手性药物、天然产物的重要中间体,可衍生出几十种制药行业急需的手性化合物,如光学活性纯的苯乙醇及其衍生物可进一步合成(L)-氯丙那林、(R)-地诺帕明等重要手性药物。利用微生物或其体内的酶,不对称合成手性仲醇及其衍生物的方法属于生物催化法。近年来,生物催化因反应条件温和、产物立体选择性高以及对环境友好等已成为不对称合成领域的研究热点。
研究证实微生物基因组中有多个还原酶基因,它们可相互干扰,有的甚至对映体选择性完全相反。如Napora等在—株解脂耶氏酵母Yarrowia lipolytica的基因组中发现了5个编码羰基不对称还原的还原酶基因,这5个蛋白就被视为假定还原酶(或潜在还原酶)。在此研究背景下,若直接从产酶微生物中分离、纯化酶蛋白,将无法精确分离到催化性能优良的还原酶。因此,前期采用基因组狩猎策略,对从自然界中筛选到的一株可将模式底物苯乙酮不对称还原成(R)-苯乙醇的Yarrowia lipolytica cmq6-8的基因组进行搜索分析,构建了一个由31个羰基还原酶基因组成的重组羰基还原酶库。从该酶库中筛选到一个催化活性稳定且催化效率相对较高的重组羰基还原酶Ylcmq6-8CR。鉴于该重组羰基还原酶在纯化过程中酶活损失较大,相比之下全细胞催化具有更好的稳定性且基因工程菌体细胞中的确定酶源及诱导调控机制可确保催化过程的高度立体选择性及底物转化率,同时基因工程菌自身具备羰基还原酶所必需的辅酶再生功能,可为全细胞催化不对称合成反应提供所需的辅酶,降低了生产成本。因此,本文以羰基还原酶基因工程菌全细胞为催化剂,探讨不对称合成(R)-苯乙醇的各种影响因素,确定其产酶和不对称合成(R)-苯乙醇的最佳条件,旨为该方法的工业化应用提供理论和实验依据。
1材料与仪器
1.1药品与试剂
苯乙酮及其衍生物、(R/S)-苯乙醇均购自中国医药集团上海化学试剂公司,纯度为99%;(s)-苯乙醇、(R)-苯乙醇等标准品购自Sigma-Aldrich,ChiraSelect,≥99.0%(sum of enantiomers,GC);異丙基-β-D硫代半乳糖苷(IPTG),上海美季生物技术有限公司;氨苄青霉素购自BIOSHARP公司;其他试剂均为市售分析纯或生物试剂。
1.2菌株及各种培养基
羰基还原酶基因工程菌E.coliBL(DE3)/pET32a-vlcmq6-8cr为课题组自建。斜面培养基(LA固体培养基):蛋白胨10.0g/L,酵母浸出粉5.0g/L,NaCl10.0g/L,氨苄青霉素0.05g/L,琼脂粉17.0gm,pH7.2;种子和初始发酵培养基均为LA液体培养基。上述培养基的灭菌条件均为121℃、20min。
1.3仪器设备
超声破碎仪(上海五相仪器仪表有限公司JY一1800L);气相色谱仪(上海海欣色谱仪器有限公司,GC960),HP Chiral 10%3-Cyclodextrin手性色谱柱(30m×0.32mm×0.25um);高速冷冻离心机(常州市万丰仪器有限公司TG-16G);低速离心机(北京医用离心机厂),等。
2方法
2.1羰基还原酶基因工程菌的诱导表达
从羰基还原酶基因工程菌的保藏斜面上刮一环菌苔于种子培养基37℃、170r/min摇床培养12h;再按10%的接种量转人发酵培养基,37℃、170r/mm摇床培养,待发酵液OD600达到0.6~0.8时,添加0.25mmol/LIPTG诱导10h。
2.2粗酶液的酶活测定
收集诱导后菌体细胞,超声破碎后取上清液,制备粗酶液。根据NAD(P)H在340nm波长下有特异性吸收,利用分光光度计检测340nm吸收值的变化确定其酶活性,反应体系如表1所示。酶活力单位定义:测定条件下每分钟消耗1umol的NAD(P)H所需的酶量。酶活力计算公式如下:
2.3产酶条件优化
2.3.1碳源、氮源及其配比
利用单因素方法,在初始发酵培养基的基础上考察碳源如葡萄糖、蔗糖、甘油,以及复合氮源及其剂量对羰基还原酶基因工程菌的生物量和粗酶活性的影响。
2.3.2诱导条件
探讨诱导剂IPTG的浓度(0.25,0.5,0.75,1mmol/L)、诱导温度(37,18℃)、诱导时间(0,6,10,16,20h)等因素对诱导酶表达量的影响。
2.4基因工程菌菌体细胞的培养及其转化
从菌种斜面上刮1环菌苔接种于20mL发酵培养基中,37℃、170r/rain摇床培养24h后收集发酵液,于6000r/min、4℃离心10min获得菌体细胞,再用pH 6.8的磷酸盐缓冲溶液洗1次后进行称重,并称取2g湿菌体转接人20mL pH 6.8的磷酸盐缓冲溶液中,按终浓度为45mmol/L加入底物苯乙酮,30℃、170r/mm,转化24h。
2.5产物GC分析
还原反应结束后,反应液用乙酸乙酯萃取(1:1,v/v),再用无水MgSO4干燥后使用GC-960气相色谱仪进行产物分析。检测条件:载气为氮气,进样器、色谱柱和FID检测器温度分别为220℃、110℃和200℃;分流比为l:100;进样量为0.1mL。分别以底物转化率(Conversion)和产物的对映体过量值(e.e.value)表示反应的转化程度和立体选择性,其表达式如下:
式中:co—一底物的初始浓度;
C苯乙醇——反应终止时的产物浓度;
CR,Cs——(R)型和(s)型产物的浓度。
2.6转化条件优化
以Conversion(%)及产物e.e.值为考察指标,利用羰基还原酶基因工程菌全细胞为催化剂,单因素方法探究影响不对称还原反应的主要因素,包括转化时间(2,4,6,12,16,20,24,28,32,36h),辅助底物种类(正丁醇、甲醇、异丙醇、乙醇、甘油、葡萄糖)及最适辅助底物浓度(2%、4%、6%、8%、10%、12%、14%、16%、18%、20%),反应体系初始pH(4、5、6、6.2、6.6、6.8、7、8、9),转化温度(20,24,28,32,34,37,40,42,45,50℃),底物浓度(35,40,45,50,55,60,65,70,75,80mmol/L),菌体细胞浓度(0.1,0.15,0.2,0.25,0.3,0.35,0.4,0.45,0.5,0.55,0.6g/ml)等,最终建立最佳转化条件。
2.7反应体系放大实验
在最佳转化条件的基础上,逐级放大转化体系:50,100,250,500,l 000,2000,3000mL,以(R)-苯乙醇的产量g/L为评价指标,考察反应体系的稳定性。(R)-苯乙醇的实际产量计算方法如下:
式中:y——产物(R)-苯乙醇的实际产量;
Conversion(%)——底物转化率;
ER——(R)-苯乙醇的对映体过量值;
Es——(s)-苯乙醇的对映体过量值;
Yo——物的理论产量,即底物全部转化成产物的质量。
3结果
3.1发酵培养基优化
3.1.1最佳碳源及配比
和初始发酵培养基相比,添加不同剂量的葡萄糖、蔗糖、甘油作为唯一碳源时,工程菌的菌体生物量和粗酶活性均有所提升,见表2。其中以10g甘油为唯一碳源时,菌体生物量和粗酶活性均最高。因此,发酵培养基中确定添加甘油为唯一碳源,添加剂量为10gm。
3.1.2最佳氮源组合及添加剂量
在碳源考察结果的基础上,进一步考察复合氮源对菌体细胞的生长和粗酶活性的影响。结果如表3所示,发酵培养基中含有复合氮源时,菌体细胞的生物量和粗酶酶活性明显高于只含有机氮源的培养基。因此,发酵培养基确定添加复合氮源,即10gm蛋白胨、5gm酵母浸粉、5g/L(NH4)SO4。
3.2诱导条件优化
3.2.1IPTG最適浓度
如图1所示,添加IPTG后目的蛋白表达量明显增多,并具有一定的浓度依赖性。但考虑到IPTG浓度过大会对菌体细胞产生毒副作用,最终确定IPTG的浓度为0.25mmol/L。
3.2.2最佳诱导时间
如图2所示,重组羰基还原酶表达量随诱导时间的延长而增加,诱导20h后酶的表达量最高,故确定最佳诱导时间为20h。
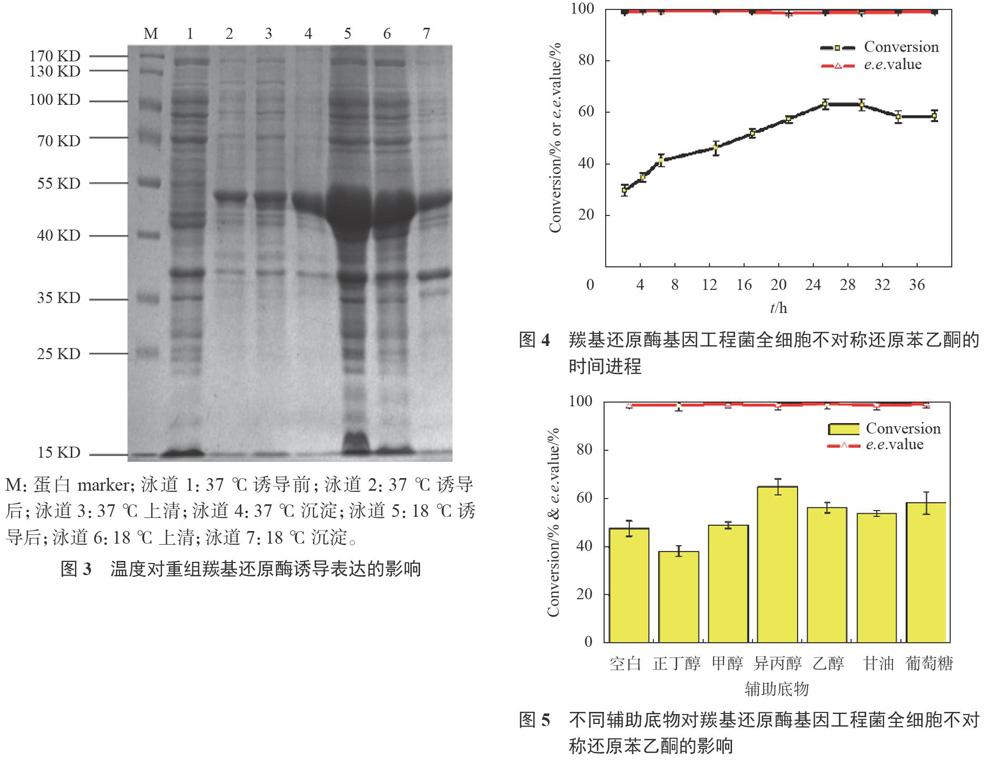
3.2.3最佳诱导温度
如图3所示,经37℃诱导大部分目的蛋白以包涵体形式存在;当诱导温度降为18℃时,不仅目的蛋白的表达量增加,且大部分以可溶形式存在于细胞破碎液中。原因可能是低温可减慢菌株的生长和代谢速度,从而有利于目的蛋白的正确折叠,有效降低包涵体的形成量,使目的蛋白多以可溶性形式存在。最佳产酶条件下,羰基还原酶基因工程菌的粗酶活最高可达6.65U/mL,是优化前最高值
3.44U/mL的1.93倍。
3.3转化条件优化
3.3.1最佳转化时间
如图4所示,随转化时间的延长,底物转化率逐渐增加;24h时,底物转化率趋于平稳。此时,延长反应时间对提高底物转化率无明显作用。分析原因可能是底物苯乙酮和不断累积的产物苯乙醇均为有机化合物,它们过量时均可抑制微生物菌体细胞的生长,从而导致细胞内的羰基还原酶活性降低,此时再延长转化时间,对底物转化率及产物e.e值的影响不大,最终确定最佳转化时间为24h。
3.3.2最佳辅助底物及其最适浓度
如图5所示,反应体系中未加辅助底物时,羰基还原酶基因工程菌仍可不对称还原部分苯乙酮。原因在于基因工程菌在发酵过程中利用培养基提供的甘油为碳源进行生长时,甘油作为能源物质经菌体细胞的分解代谢可产生羰基还原酶所必需的辅酶NAD(P)H,因此菌体细胞内累积了一定量的辅酶,它们与胞内羰基还原酶的共同作用实现了底物苯乙酮的不对称还原。因此,为实现辅酶的再生需向反应体系中额外添加甘油等辅助底物。当以2%异丙醇为辅助底物时,获得底物转化率最大,葡萄糖和乙醇次之,而产物e.e值相差不大,故确定异丙醇为最适辅助底物。
如图6可知,未添加辅助底物时,由于没有后续能源物质补给,细胞中还原型辅酶数量逐渐减少甚至消失,所以底物转化率较低。随着添加的辅助底物异丙醇的浓度增加,底物转化率也随之增加,当浓度达到10%时,底物转化率达到最大值,若再增加异丙醇的浓度,底物转化率不增反降。考虑到异丙醇浓度对产物e.e值影响较小,最终确定辅助底物异丙醇的最适合浓度为10%。
3.3.3反应体系最适初始pH及温度
如图7所示,转化体系初始pH为7.0时底物转化率和产物e.e值最大,故确定反应体系的最适初始pH为7.0。随着温度的增加获得的底物转化率也不断加大,当温度达到37℃时,获得的底物转化率最大,若温度继续增高,底物转化率反而下降,见图8。这是由于过高的温度会使化学本质为蛋白质的酶变性失活,考虑到温度对产物e.e值的影响不大,故确定转化最适温度为37℃。
3.3.4最适底物浓度
为获得更多产物,尝试不断增大转化体系中的底物浓度。如图9所示,随着底物浓度的增加,获得的底物转化率也增大。当底物浓度增加到60mmol/L时获得的底物转化率达到最大值,若再进一步增加底物浓度,底物转化率不增反降,原因在于底物浓度过大会抑制菌体细胞生长以及细胞内的氧化还原酶活性,因此确定最适底物浓度为60mmol/L。
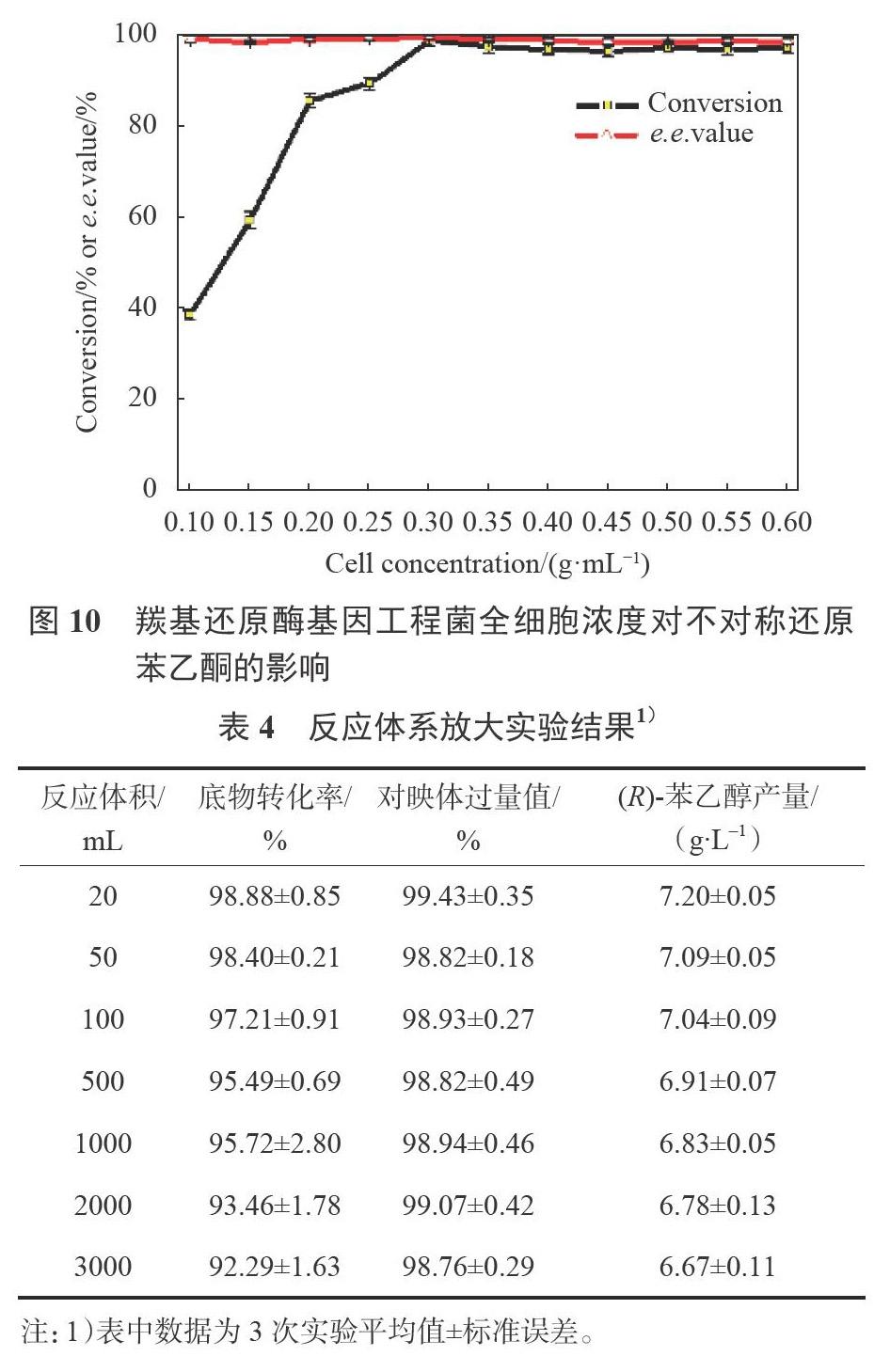
3.3.5最适细胞浓度
如图10所示,反应体系中基因工程菌细胞含量逐渐增加,底物转化率也逐渐增高。这是因为当底物浓度不变时,增加菌体细胞的量相当于增加反应体系中催化剂的数量,进而加快了转化进程,底物转化率增加。当细胞浓度增加到0.3g/mL时,底物转化率最大,若再增加菌体细胞的含量,底物转化率变化不大。分析原因可能是反应体系中过多的菌体细胞,不能很好地分散在磷酸盐反应体系中,有的菌体细胞在振摇过程中还发生贴壁现象,导致发挥催化作用的细胞含量实际并没有增加,甚至减少了。考虑到细胞浓度对产物e.e.值的影响不大,故转化反应的最适细胞浓度为0.3g/mL。
3.4反应体系放大实验
最佳转化条件下反应体系扩大至3000mL,产物e.e值仍保持在99.0%左右,底物转化率为92%以上,(R)-苯乙醇的产量也可达6.67g/L,见表4。
4结束语
本文利用前期构建的羰基还原酶基因工程菌全细胞为催化剂,探讨发酵培养基、诱导条件等因素对重组羰基还原酶活性的影响,建立最佳产酶条件,羰基还原酶基因工程菌粗酶活最高达6.65U/mL,是优化前的1.93倍。鉴于重组羰基还原酶在纯化过程中酶活性损伤较大,故利用羰基还原酶基因工程菌全细胞为催化剂,深入探究了转化时间、温度、pH等多种因素对转化反应影响,建立最佳转化条件,在此基础上将反应体系扩大至3000mL,产物e.e.值仍保持在99.0%左右,底物转化率为92%以上,(R)-苯乙醇的产量也可达6.67g/L。后续为获得稳定的羰基还原酶制剂,将进一步研究酶的纯化工艺,以提高其催化活性的稳定性。同时,还将尝试细胞固定化等方法,解决生物催化剂回收和重复利用等技术问题,为该方法的工业化应用奠定基础。
