
湖北红椿天然种群小叶表型性状变异研究
2019-06-11 06:03:50陈文学明安觉宋丛文付秋生
植物资源与环境学报 2019年2期
汪 洋, 陈文学, 明安觉, 雍 军, 宋丛文,①, 付秋生,①
(1. 湖北生态工程职业技术学院, 湖北 武汉 430200; 2. 竹山县林业局, 湖北 竹山 442200;3. 竹山县国土资源局, 湖北 竹山 442200)
植物表型性状反映了基因型对环境变化的适应性[1]。 植物在漫长的进化过程中,应对环境差异产生的表型变异体现了植物的生存智慧,其表型变异往往在适应和进化上有重要意义[1],是遗传多样性研究的重要内容[2]。 植物表型研究主要关注植物在其分布区内各种环境下的表型变异[3],被广泛用于揭示天然种群的遗传变异及其与地理格局的关系[4-8]。
红椿(ToonaciliataRoem.) 隶属于楝科(Meliaceae)香椿属(ToonaRoem.),为落叶或半落叶乔木,其木材优良,是珍贵用材树种[9]。 由于过度开发且天然更新较慢,红椿天然分布区日益减少[10],其天然种群内植株数量也随之减少,目前红椿已被列为国家二级重点保护野生植物[11]。 鉴于红椿的濒危现状,研究人员已对其生理[12-15]、生长特性[16-17]、造林[18]、种群生态学[19-20]、优树选择[21-23]、表型遗传研究的取样技术[24]以及红椿提取物抗癌能力[25]等方面进行了深入研究。 然而,对红椿的科学保护应结合其现存种群规模与遗传多样性水平进行[26]。 李培等[27]利用SRAP 标记对来自中国的29 个种源和来自澳大利亚的1 个种源的红椿进行系统地遗传多样性分析,结果表明红椿不同种源间的分化程度较高。叶片是植物进化过程中对环境变化比较敏感且可塑性较大的器官[28],植物叶片的形态与其营养和其他生理、生态因子以及繁殖密切相关[29],因此,不同程度的环境异质性,影响植物叶片表型的可塑性或植物对环境的适应性。 红椿为强阳性植物,叶片是光合作用的主要器官,研究红椿叶片的性状和功能,有助于分析其种群内和种群间的表型变异多样性水平,对了解其生长发育和逆境响应具有重要意义。
在湖北地区,红椿主要分布于西南部、西北部和东南部的山区以及中北部的荆山山脉,中南部的江汉平原尚未发现有分布。 由于红椿分布区生境差异较大,地理分断特征明显,研究红椿天然种群的表型变异及其地理变异格局具有重要意义。 本文对湖北16个红椿天然种群的10 个小叶表型性状进行分析,探讨了种群间和种群内小叶表型性状的变异状况及其与地理环境间的关系,以期为湖北红椿种质资源保护、种源选择和开发利用提供基础数据。
1 材料和方法
1.1 材料
在全面系统普查湖北红椿种质资源的基础上,综合考虑种群位置和大小等实际情况,尽量覆盖湖北红椿天然分布区,最终确定16 个红椿天然种群为采样点,其基本概况见表1,表中气象数据来源于中国气象数据网(https:∥data. cma. cn/data/cdindex. html)、地方气象站和部分种群所在自然保护区官网。
1.2 小叶表型性状的选取及测定
红椿复叶特征较为稳定,其表型差异及变种间鉴别主要依据小叶的叶长、叶宽、叶柄长与叶尖角等指标的差异[9]。 于2015 年11 月,在16 个红椿种群中分别选择10 株无病虫害且生长正常的成年植株,株间距大于等于35 m,按东、西、南、北4 个方向均匀采集各植株树冠中部的枝条。 每株采集20 枝复叶,在叶轴右侧中部取1 枚小叶,每个种群200 枚小叶。 用电子游标卡尺(精度0.01 mm)测量小叶柄长、小叶长(含小叶柄)、小叶宽、宽基距(小叶最宽处距小叶基的距离)和脉左宽(小叶最宽处左缘到中脉的距离),用量角器(精度0.1°)测量小叶尖角(小叶尖与叶片边缘的夹角)[2,6,30],并计算小叶长小叶宽比、小叶柄长小叶长比、脉左宽小叶宽比和宽基距小叶长比[30]。
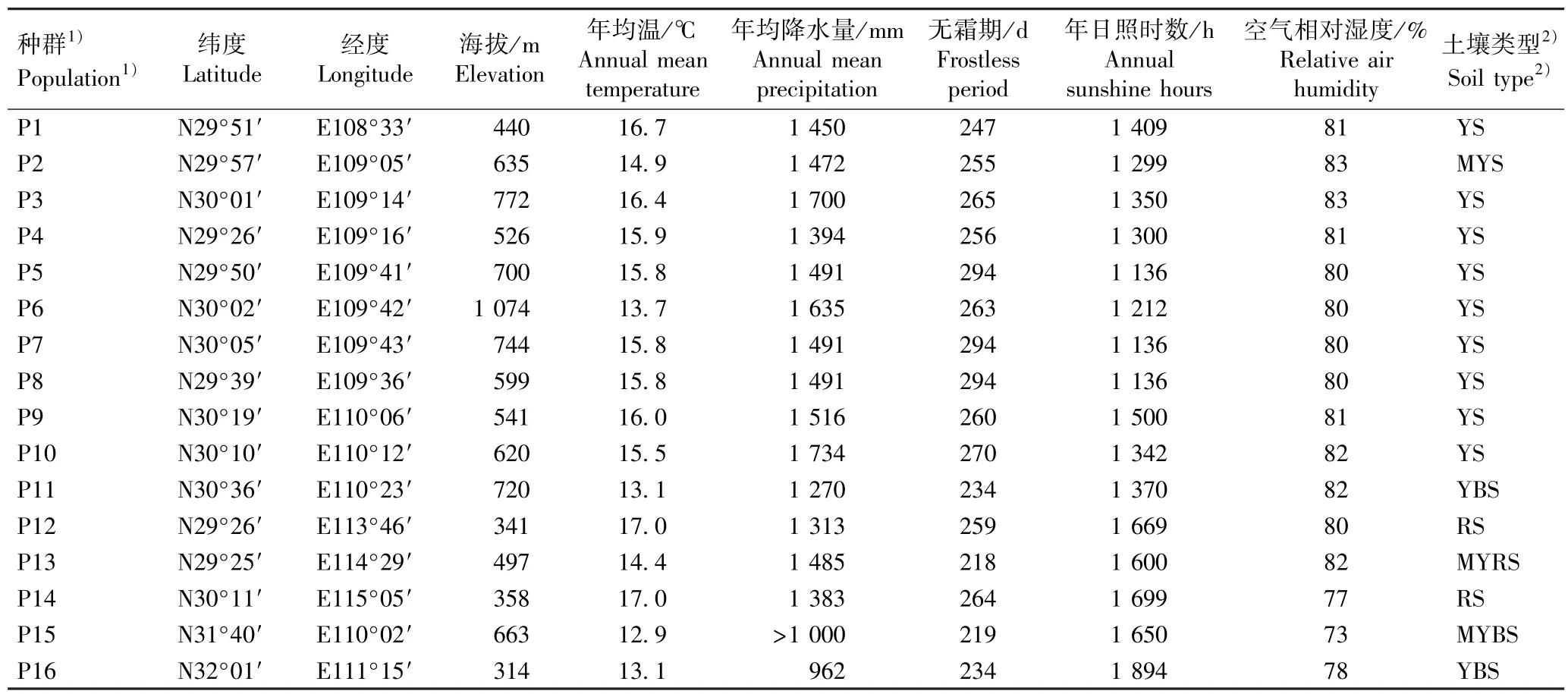
表1 湖北16 个红椿天然种群的基本概况Table 1 Basic situation of 16 natural populations of Toona ciliata Roem. in Hubei Province
1.3 数据统计分析
对16 个红椿种群小叶表型性状在种群间和种群内的差异显著性进行巢式方差分析[31],线性模型为Yijk=μ+Si+T(i)j+ε(ij)k。 式中,Yijk为第i个种群第j个单株第k个观测值,μ 为总体均值,Si为第i个种群的效应值,T(i)j为第i个种群第j个单株的效应值,ε(ij)k为随机误差。 表型分化系数(Vst)可以近似解释种群间表型分化程度,计算公式为式中,为种群间方差分量为种群内方差分量[32]。 计算各小叶表型性状的均值、标准差和变异系数。 采用Duncan's 新复极差法进行多重比较。 对小叶表型性状间以及其与采样点环境因子进行相关性分析。 用非加权平均法(UPGMA)进行系统聚类分析,运用Mantel 检验[33]研究地理距离的自然对数与遗传距离间的相关性。 巢式方差和表型分化系数采用SAS 9.4 软件计算,多重比较和相关性分析采用SPSS 22.0 软件,聚类分析采用NTSYS-pc2.1 软件。
2 结果和分析
2.1 不同红椿种群小叶表型性状的比较
湖北红椿种群间及种群内小叶表型性状的方差分析结果见表2。 由表2 可以看出:红椿10 个小叶表型性状在种群间存在显著(P<0.05)或极显著(P<0.01)差异,表明红椿小叶表型性状在种群间变异程度较高。 小叶长、小叶柄长、小叶宽、宽基距、小叶长小叶宽比和小叶柄长小叶长比在种群内存在极显著差异,其余4 个小叶表型性状在种群内差异不显著。总体来看,湖北红椿小叶表型性状在种群间的变异大于种群内。
湖北16 个红椿天然种群小叶表型性状的比较结果见表3。 由表3 可以看出:红椿10 个小叶表型性状在种群间存在显著差异。 小叶长的变化范围为15.070~22.221 cm,均值为18.794 cm,其中,黄石黄荆山(P14)种群的小叶最长,宣恩金盆村(P5)种群的小叶最短。 小叶柄长的变化范围为5.35 ~8.53 mm,均值为7.09 mm,其中,宣恩大卧龙(P7)种群的小叶柄最长,通山九宫山(P13)种群的小叶柄最短。小叶宽的变化范围为5.827~7.828 cm,均值为7.044 cm,其中,宣恩肖家湾(P6)种群的小叶最宽,P7 种群的小叶最窄。 宽基距的变化范围为4.345 ~6.477 cm,均值为5.736 cm,其中,P14 种群的宽基距最大,P5 种群的宽基距最小。 脉左宽的变化范围为3.106~4.158 cm,均值为3.769 cm,其中,P6 种群的脉左宽最大,P7 种群的脉左宽最小。 小叶尖角的变化范围为19.70°~38.35°,均值为30.69°,其中,利川堡上(P1)种群的小叶尖角最大,P14 种群的小叶尖角最小。 小叶长小叶宽比的变化范围为2.455 ~3.374,均值为2.690,其中,P14 种群的小叶长小叶宽比最大,P13 种群的小叶长小叶宽比最小。 小叶柄长小叶长比的变化范围为0.028 ~0.054,均值为0.038,其中,P7 种群的小叶柄长小叶长比最大,竹山洪坪(P15)种群的小叶柄长小叶长比最小。 脉左宽小叶宽比的变化范围为0.496 ~0.559,均值为0.536,其中,宣恩红旗坪(P8)种群的脉左宽小叶宽比最大,P13 种群的脉左宽小叶宽比最小。 宽基距小叶长比的变化范围为0.272 ~0.344,均值为0.306,其中,P13 种群的宽基距小叶长比最大,P15 种群的宽基距小叶长比最小。

表2 湖北红椿种群间及种群内小叶表型性状的方差分析结果1)Table 2 Result of variance analysis on leaflet phenotypic traits of Toona ciliata Roem. among populations and within population in Hubei Province1)
表3 湖北红椿16 个天然种群小叶表型性状的比较(±SD)Table 3 Comparison on leaflet phenotypic traits of 16 natural populations of Toona ciliata Roem. in Hubei Province (±SD)

表3 湖北红椿16 个天然种群小叶表型性状的比较(±SD)Table 3 Comparison on leaflet phenotypic traits of 16 natural populations of Toona ciliata Roem. in Hubei Province (±SD)
种群1)Population1)小叶表型性状2) Leaflet phenotypic traits2)LL/cm LLP/mm WL/cm LMWLB/cm WLBEM/cm P1 16.891±1.011d 7.99±1.25a 6.680±0.583ef 5.052±0.443gh 3.668±0.306de P2 19.186±1.065c 8.16±3.04a 6.959±0.749def 5.508±0.834fgh 3.837±0.613bcd P3 17.449±1.786d 5.98±1.11cde 6.736±0.988ef 5.626±0.829defg 3.429±0.435efg P4 19.242±2.044bc 8.28±1.06a 7.497±1.000abc 5.575±0.990efgh 3.970±0.558abc P5 15.070±0.110f 7.50±1.40ab 5.837±0.580g 4.345±0.656i 3.239±0.361gh P6 19.564±1.332bc 8.05±0.94a 7.828±0.443a 6.263±0.770abc 4.158±0.218a P7 15.862±1.726ef 8.53±1.42a 5.827±0.730g 4.984±0.978h 3.106±0.387h P8 20.311±2.348b 8.50±1.75a 7.335±0.829bc 6.271±1.061abc 4.109±0.554ab P9 19.492±1.693bc 6.27±1.19cde 7.152±0.894cde 5.788±0.758bcdef 3.869±0.435abcd P10 19.903±1.051bc 6.53±1.43cd 7.653±0.662ab 6.221±0.707abcd 4.092±0.270ab P11 19.540±0.768bc 5.61±1.50de 7.317±0.464bc 6.054±0.834abcdef 3.949±0.590abc P12 19.304±1.284bc 7.78±1.30ab 7.721±0.780ab 6.139±1.097abcde 4.056±0.335ab P13 16.639±1.107de 5.35±0.89e 6.790±0.485ef 5.682±0.978cdef 3.362±0.296fgh P14 22.221±1.751a 6.49±1.36cd 6.597±0.466f 6.477±1.149a 3.615±0.258def P15 20.158±1.673bc 5.51±1.38de 7.123±0.450cde 5.464±0.537fgh 3.772±0.242cd P16 19.885±1.264bc 6.98±1.32bc 7.658±0.407ab 6.329±0.683ab 4.079±0.179ab均值Mean 18.794±2.339 7.09±1.81 7.044±0.895 5.736±1.008 3.769±0.508
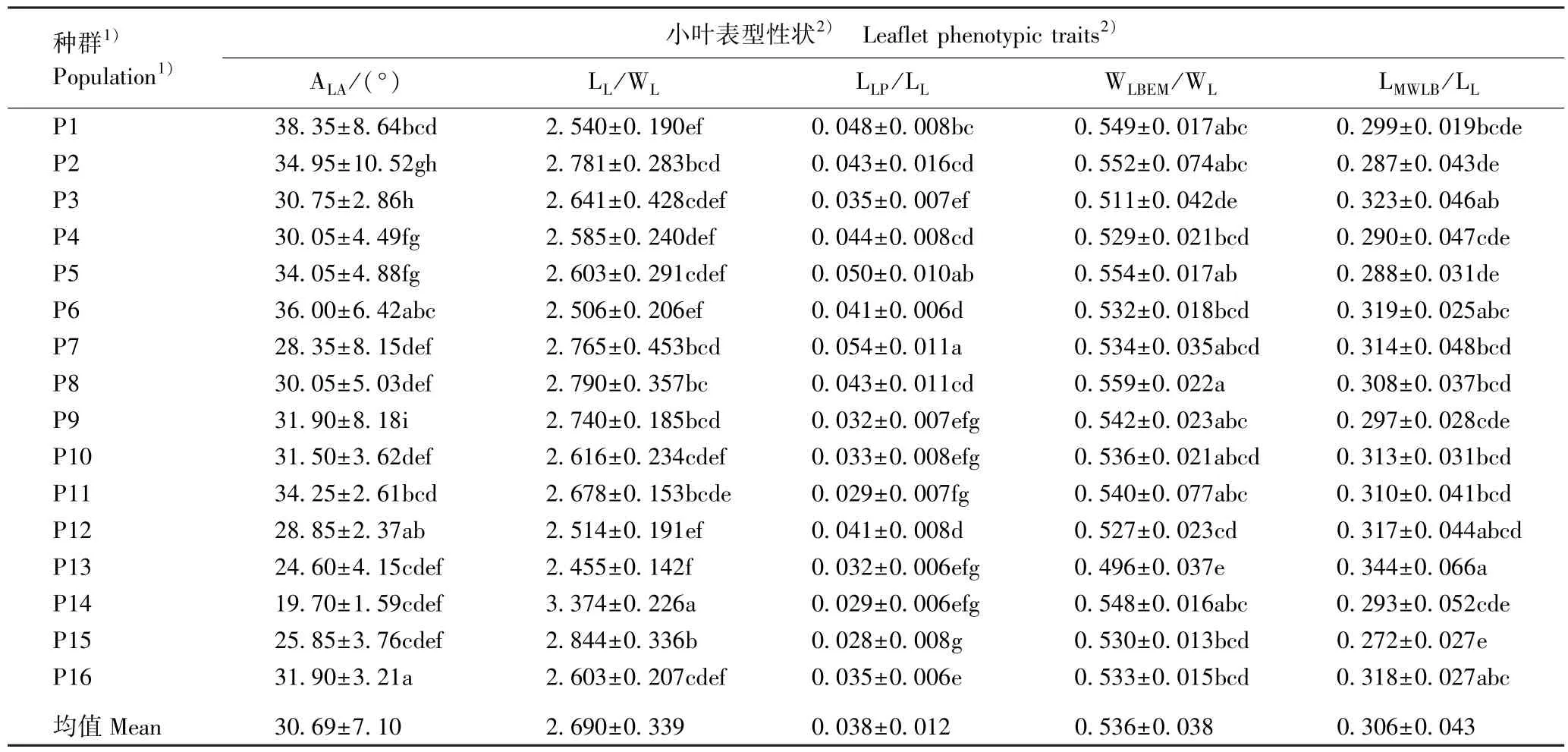
续表3 Table 3 (Continued)
2.2 不同红椿种群小叶表型性状的变异分析
湖北16 个红椿天然种群小叶表型性状的变异系数见表4。 由表4 可以看出:红椿10 个小叶表型性状变异系数的均值在3.31%~21.99%之间,由大到小依次为小叶柄长小叶长比(21.99%)、小叶柄长(19.84%)、小叶尖角(15.35%)、宽基距(13.44%)、宽基距小叶长比(11.28%)、脉左宽(9.64%)、小叶长小叶宽比(9.51%)、小叶宽(9.40%)、小叶长(7.56%)、脉左宽小叶宽比(3.31%),其中,小叶柄长和小叶尖角变异系数的均值较大,小叶长变异系数的均值较小。
由表4 还可以看出: 16 个红椿种群内小叶表型性状变异系数的均值从大到小依次为咸丰横石梁(P2) 种 群(17.59%)、 宣 恩 大 卧 龙(P7) 种 群(14.52%)、宣恩红旗坪(P8)种群(14.34%)、恩施马鹿河(P3)种群(14.14%)、建始青龙河(P9)种群(13.33%)、来凤三寨坪(P4)种群(13.01%)、宣恩金盆村(P5) 种群(12.15%)、竹山洪坪(P15) 种群(11.85%)、崇阳庙圃(P12)种群(11.37%)、鹤峰彭家湾(P10)种群(11.22%)、巴东野三关(P11)种群(11.10%)、通山九宫山(P13)种群(10.95%)、利川堡上(P1)种群(10.41%)、黄石黄荆山(P14)种群(9.45%)、宣恩肖家湾(P6)种群(9.37%)、谷城玛瑙观(P16)种群(9.32%)。 红椿种群内小叶表型性状变异系数的均值大多未达到15%,变异水平较低。
2.3 红椿小叶表型性状的表型分化分析
湖北16 个红椿天然种群小叶表型性状的方差分量和表型分化系数见表5。 由表5 可以看出:红椿种群间10 个小叶表型性状方差分量百分比的均值为32.36%,种群内10 个小叶表型性状方差分量百分比的均值为6.27%,种群间10 个小叶表型性状的方差
分量明显大于种群内。 种群间10 个小叶表型性状的表型分化系数在63.51% ~98.29%之间,均值为80.73%,其中,种群间小叶尖角的表型分化系数最大,为98.29%,种群间脉左宽小叶宽比的表型分化系数最小,为63.51%,说明红椿小叶表型变异主要来源于种群间。

表4 湖北16 个红椿天然种群小叶表型性状的变异系数Table 4 Coefficient of variation of leaflet phenotypic traits of 16 natural populations of Toona ciliata Roem. in Hubei Province
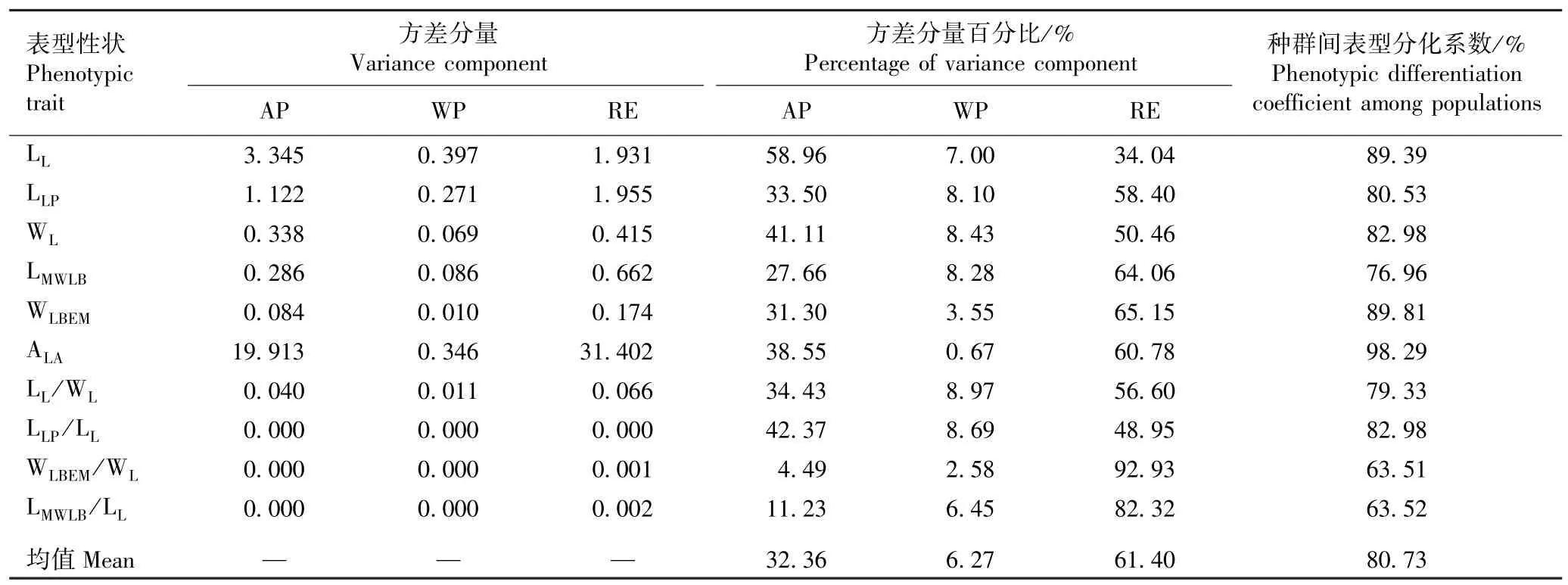
表5 湖北16 个红椿天然种群小叶表型性状的方差分量和表型分化系数1)Table 5 Variance component and phenotypic differentiation coefficient of leaflet phenotypic traits of 16 natural populations of Toona ciliata Roem. in Hubei Province1)
2.4 红椿小叶表型性状间及其与环境因子的相关性分析
湖北红椿小叶表型性状间及其与环境因子的相关系数分别见表6 和表7。
由表6 可以看出:红椿小叶长与小叶宽、宽基距和脉左宽呈极显著(P<0.01)正相关,与小叶柄长小叶长比呈极显著负相关;小叶柄长与小叶柄长小叶长比呈极显著正相关,与脉左宽小叶宽比呈显著(P<0.05)正相关;小叶宽与宽基距和脉左宽呈极显著正相关;宽基距与脉左宽呈极显著正相关,与小叶尖角和小叶柄长小叶长比呈显著负相关;小叶柄长小叶长比与脉左宽小叶宽比呈显著正相关。 小叶尖角、小叶长小叶宽比和宽基距小叶长比与其他小叶表型性状相关性较低,表现为相对独立的性状。
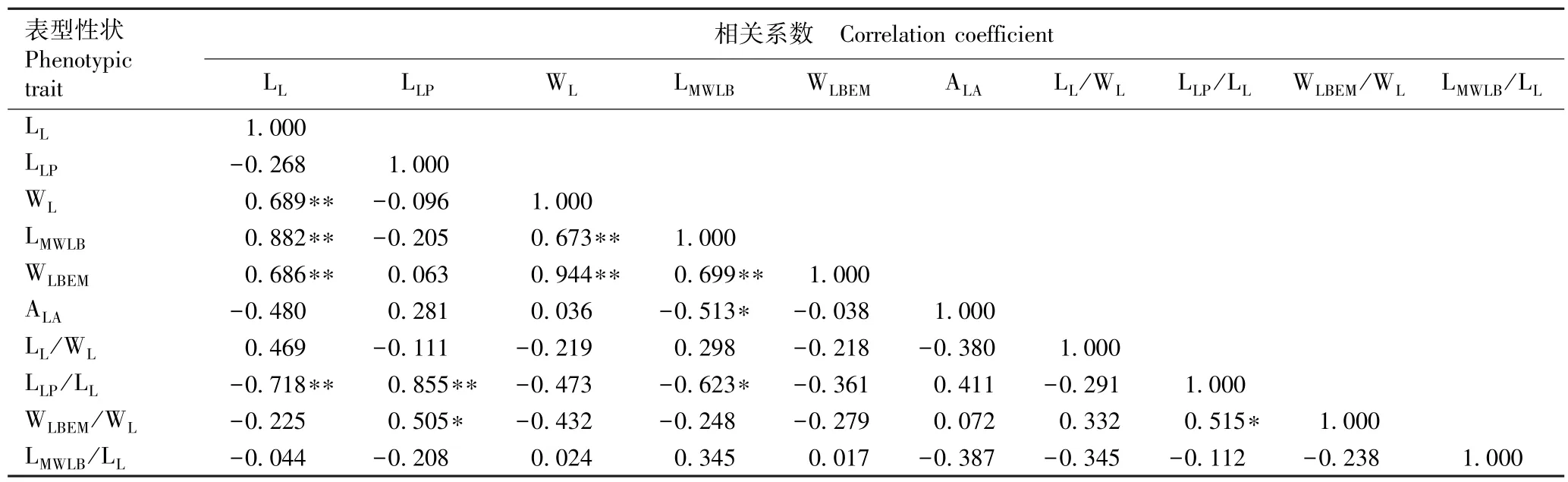
表6 湖北红椿小叶表型性状间的相关系数1)Table 6 Correlation coefficient among leaflet phenotypic traits of Toona ciliata Roem. in Hubei Province1)
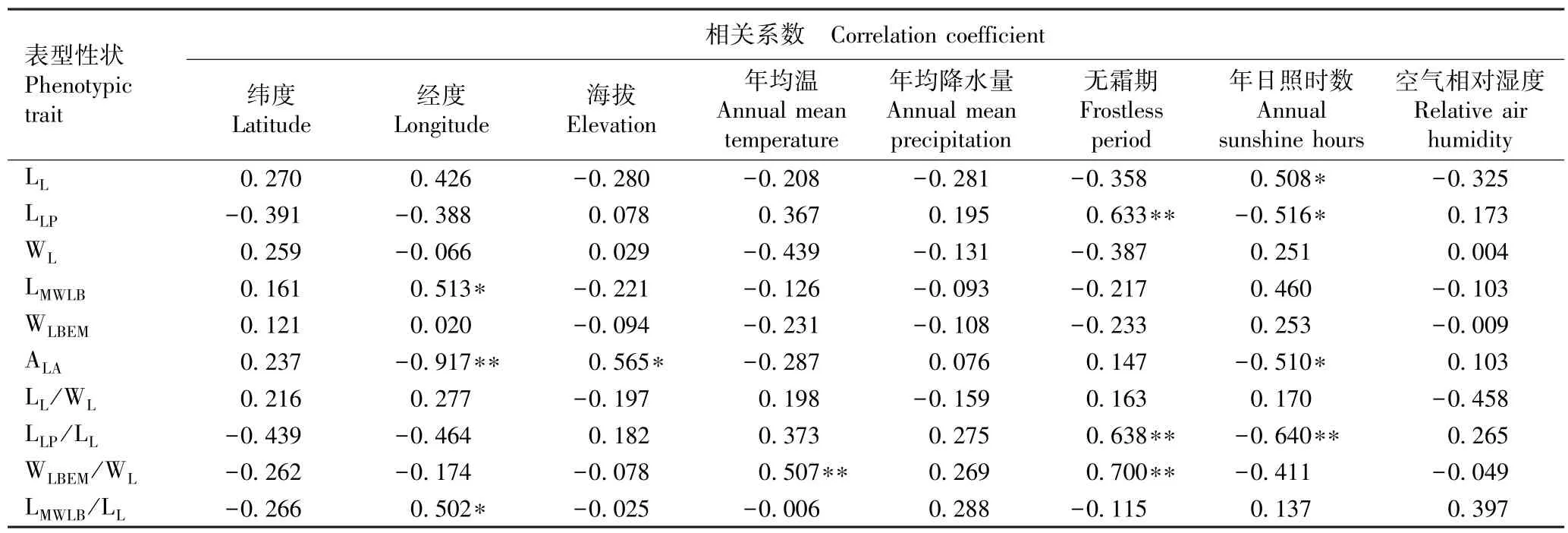
表7 湖北红椿小叶表型性状与环境因子的相关系数1)Table 7 Correlation coefficient of leaflet phenotypic traits of Toona ciliata Roem. with environmental factors in Hubei Province1)
由表7 可以看出:红椿小叶长与年日照时数呈显著正相关;小叶柄长与无霜期呈极显著正相关,与年日照时数呈显著负相关;宽基距与经度呈显著正相关;小叶尖角与经度和年日照时数分别呈极显著和显著负相关,与海拔呈显著正相关;小叶柄长小叶长比与无霜期呈极显著正相关,与年日照时数呈极显著负相关;脉左宽小叶宽比与年均温和无霜期呈极显著正相关;宽基距小叶长比与经度呈显著正相关。 各小叶表型性状与纬度、年均降水量和空气相对湿度的相关性均不显著。
2.5 不同红椿种群的聚类分析
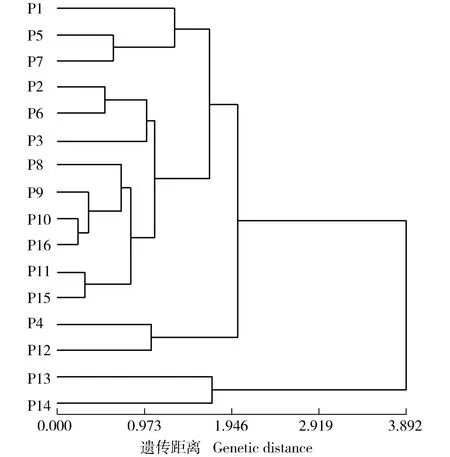
图1 基于小叶表型性状的湖北16 个红椿天然种群的聚类图Fig. 1 Dendrogram of 16 natural populations of Toona ciliata Roem.in Hubei Province based on leaflet phenotypic traits
基于供试的10 个小叶表型性状,采用非加权平均法(UPGMA)对湖北16 个红椿天然种群进行聚类分析,结果见图1。 由图1 可以看出:在遗传距离3.892 处,通山九宫山(P13) 种群与黄石黄荆山(P14)种群聚为一类,其他14 个种群聚为另一类。 在遗传距离2.018 处,后一类可划分为2 个亚类,其中,鄂西南的来凤三寨坪(P4)种群和鄂东南的崇阳庙圃(P12)种群聚为一个亚类,鄂西南的利川堡上(P1)种群、宣恩金盆村(P5)种群、宣恩大卧龙(P7)种群、咸丰横石梁(P2)种群、宣恩肖家湾(P6)种群、恩施马鹿河(P3)种群、宣恩红旗坪(P8)种群、建始青龙河(P9)种群、鹤峰彭家湾(P10)种群和巴东野三关(P11)种群以及鄂西北的竹山洪坪(P15)种群和谷城玛瑙观(P16)种群聚为另一个亚类。 16 个种群间的地理距离的自然对数与遗传距离的Mantel 检验结果表明:二者间的相关性不显著(R= 0.205,P=0.120)。
3 讨论和结论
3.1 红椿种群表型变异来源
本研究中,红椿种群间小叶表型性状表型分化系 数的均值为80.73%,高于无患子(SapindussaponariaLinn.) (62.21%)[34]、 蒙 古 栎(QuercusmongolicaFisch. exLedeb.) ( 53.97%)[35]、 长 柄 扁 桃(AmygdaluspedunculataPall.)(45.90%)[7]、白皮松(PinusbungeanaZucc. ex Endl.)(22.86%)[31]和青梅(VaticamangachapoiBlanco)(18.31%)[6],与滇 龙胆 (GentianarigescensFranch.ex Hemsl.)(73.14%)[36]和山杏〔Armeniacasibirica(Linn.)Lam.〕(73.03%)[37]接近,但低于夏蜡梅(Calycanthus chinensisCheng et S. Y. Chang)(89.30%)[38],说明红椿种群间表型分化水平较高。 为了应对不同地区的环境差异,红椿小叶形成了稳定的表型遗传特征。首先,湖北整体为西部高于东部、中部为平原和丘陵,地貌差异较大。 红椿主要分布在湖北东、西两端,中部江汉平原地区尚未发现天然分布,种群间的环境异质性差异程度远大于种群内,因此,种群间遗传变异的可能性高于种群内。 其次,地理隔离会造成种群间基因交流不频繁,红椿小叶表型性状种群间表型分化系数达到80.73%,远高于种群内表型分化系数,说明红椿天然种群小叶表型性状变异主要源自种群间,也反映出红椿不同天然种群基因与环境互作的复杂性及其适应环境选择压力的广泛程度,是不同环境选择的结果,也是种群分化的源泉[39]。
3.2 红椿小叶表型性状的变异特征
本研究中,红椿小叶表型性状变异系数均值的变化范围较大(3.31%~21.99%),变异主要表现在小叶柄长(19.84%)、小叶尖角(15.35%) 和宽基距(13.44%)上。 湖北16 个红椿种群中,仅咸丰横石梁(P2) 种群内小叶表型性状变异系数的均值(17.59%)大于15%,其他种群均低于15%,表明湖北红椿种群内小叶表型性状变异水平均较低。 推测造成该结果的原因包括2 个方面:其一,虽然研究选样时尽量考虑了取样距离的最大化,但香椿属植物的小种群特征会造成种群内遗传水平下降,使种群内表型性状变异水平较低;其二,种群内较低变异水平较低可能与大部分种群内微生境异质性较低有关[40]。
3.3 红椿小叶表型性状间及其与环境因子的关系
红椿小叶表型性状间的相关性分析结果表明:小叶越长,宽基距越长,小叶柄越短,小叶尖角越小,小叶面由卵形向披针形变化;经度与红椿小叶长呈正相关,与宽基距呈显著正相关,但与小叶尖角呈极显著负相关,进一步说明小叶型向披针形变化的显著程度与地理经度变化密切相关。 随经度增大,鄂西南、鄂西北种群与鄂东南种群所处地区海拔的差异较大(鄂东南3 个种群的平均海拔低于400 m,而鄂西南和鄂西北地区平均海拔为640 m),鄂东南种群的年均温明显高于鄂西南和鄂西北种群,同时年日照时数也明显变长。 在日照时数较短时,小叶可能通过增大叶尖角和增长小叶柄来获取较大光合面积以提高光合效率。 红椿小叶表型性状与环境因子的相关性很可能与湖北地区的纬度跨度远小于经度跨度以致行政区划内取样不足有关,因此,应增加采样点,以获得更准确可靠的研究结果。
3.4 红椿表型变异趋势及种质资源的利用
由于分布和环境条件等因子的综合作用,植物会形成连续变异、不连续变异以及随机变异等多种地理变异模式[41]。 红椿小叶表型性状变异既有连续性又有随机性,如通山九宫山(P13)种群与黄石黄荆山(14)种群聚为一类,鄂西南种群〔利川堡上(P1)种群、咸丰横石梁(P2)种群、恩施马鹿河(P3)种群、宣恩金盆村(P5)种群、宣恩肖家湾(P6)种群、宣恩大卧龙(P7)种群、宣恩红旗坪(P8)种群、建始青龙河(P9)种群、鹤峰彭家湾(P10)种群和巴东野三关(P11)种群〕与鄂西北种群〔竹山洪坪(P15)种群和谷城玛瑙观(P16)种群〕聚为一类,均为相邻地理种群聚类,表现为局部区域化变异和连续变异。 而鄂西南的来凤三寨坪(P4)种群与鄂东南的崇阳庙圃(P12)种群聚为一类,表现出种群间较高的变异水平,但Mantel 检验结果表明:红椿种群间的变异与地理距离相关性不显著,地理变异中存在随机性或不连续性。 小叶尖角和宽基距等小叶表型性状与经度显著相关,说明虽然红椿种群间没有严格按照地理距离聚类,但湖北红椿小叶表型性状变异仍存在沿经度梯度变异的规律性。
逐步累积的随机变异和连续变异是较大分布区域上红椿显著地理隔离模式的基础,因此,在红椿种质资源保护与遗传资源改良研究时,对于变异较小的种群,收集种质材料时应尽可能增加种群的数量,减少种群内的取样数;如果种群内的变异较为丰富,应尽量增加该种群内个体的取样数,如在P2 种群的种质资源收集时,应尽量多考虑不同个体的选择。 P14种群作为鄂东南红椿种质保存的“孤岛”,处于较为独特的地理位置,是连接湖北东南部和周边省份红椿天然种群之间的“驿站”,对于减少居留种灭亡的风险和物种保护有重要意义,应该重点保护。
猜你喜欢
红蜻蜓·高年级(2022年10期)2022-12-31 06:38:50
东北林业大学学报(2022年7期)2022-08-04 01:48:02
小天使·四年级语数英综合(2022年6期)2022-06-21 12:49:36
High Technology Letters(2021年4期)2022-01-09 02:08:16
作文小学中年级(2021年4期)2021-05-07 01:17:50
河北果树(2020年4期)2020-11-26 06:05:00
海峡姐妹(2019年1期)2019-03-23 02:42:40
中成药(2018年9期)2018-10-09 07:18:46
乡村科技(2018年17期)2018-08-16 02:27:50
湖北林业科技(2016年5期)2016-12-02 10:23:28
