
干旱胁迫对2种梨砧木生长及叶绿素荧光特性的影响
2019-06-05 03:14:26李俊豪朱霄飞李六林
西北农业学报 2019年5期
解 斌,李俊豪,赵 军,朱霄飞,李六林
(1.山西农业大学 园艺学院,山西太谷 030801;2.江苏省溧阳市别桥镇农业服务中心,江苏溧阳 213321;3.山西省晋中市祁县果树产业技术服务中心,山西祁县 030900)
随着全球气候变化,水分亏缺日益严重[1],干旱严重制约果树产业发展。梨是中国重要的栽培果树之一,树体高大,根系分布深,耗水量大,但其产地多为干旱、半干旱地区,降水时间分布不均[2-3],干旱缺水严重影响梨树栽培生产。在梨树苗木的嫁接繁育中,砧木对其抗旱性起决定性作用[4]。杜梨(PyrusbetulaefoliaBunge.)为北方梨树栽培常用砧木;豆梨(PyruscalleryanaDecne.)常被用作砂梨的砧木。董星光等[5]研究发现资源起源生态区不同使梨砧木间茎干组织结构形成巨大差异,杜梨导管分子管径大等的特点,保证其在干旱下的水分运输,抗旱性明显优于豆梨。目前,关于梨砧木响应干旱的研究多集中于组织结构[6],遗传多样性[7]及抗氧化酶[8]等方面,有关光合作用和叶绿素荧光参数变化规律的系统性研究较少。
叶片作为植株响应干旱最敏感的部位,其形态结构及生理变化可直接反映植株适应和抵御干旱的能力,通过叶绿素荧光特性可分析叶片对光能的吸收和利用[9]。Naidoo等[10]、麻云霞等[11]认为抗旱性强的植物能调节自身的叶绿物质含量等生理学指标主动适应干旱逆境。Wang等[12]、曹慧等[13]研究发现水分胁迫会使苹果属植物幼苗叶绿素降解加剧,光合速率大幅下降,最大光化学效率(Fv/Fm)、光化学猝灭系数(qP)降低,而非光化学猝灭系数(NPQ)不断上升以减轻干旱对树体造成的伤害。Stellfeldt等[14]在研究中发现轻度干旱胁迫对植株无明显影响,严重水分亏缺会使植物电子传递受阻,热耗散增加,从而使光合效率降低,生长减缓。王延秀等[4]在对3种苹果砧木的研究中发现干旱胁迫下抗性品种能够较好地保持细胞结构的完整性,维持光合作用相对稳定,并证明光合速率与苹果砧木抗旱能力的相关性。因此,本研究以2种梨砧木为试材进行干旱模拟试验,分析其生长状态、光合及叶绿素荧光特性等对干旱胁迫的响应与适应,探讨不同梨砧木适应干旱的能力及对策,以期为梨砧木苗期精细化水分管理及耐旱机制的深入研究提供一定的理论基础。
1 材料与方法
1.1 植物材料
供试材料分别为杜梨(PyrusbetulaefoliaBunge.)和豆梨(PyruscalleryanaDecne.)的2 a生实生幼树。杜梨的种子采自山西省绛县中条山区;豆梨的种子来自于湖北省果茶研究所。2016年11月将种子层积处理5个月后催芽、播种于穴盘中,栽培基质为细沙;2017年5月将6叶龄幼苗移栽至塑料花盆(上口径16.1 cm、下口径11 cm、高14 cm)中,盆栽土由腐熟猪粪、田园土和细砂按体积比1∶2∶3配制,每盆1株,常规管理;2018年4月中旬选择大小基本一致的苗木共计40株移植入花盆(高18 cm、上口径21 cm、下口径12 cm)中,每盆装干土2.90 kg,其土壤饱和含水量为41.3%,每盆1株,常规管理;2018年5月,将供试植株放入遮雨棚中,幼树平均株高和基径分别约为55.0 cm和0.8 cm。
1.2 处 理
试验设以下处理:轻度干旱胁迫(LD):土壤相对含水量为60%±5%;中度干旱胁迫(MD):土壤相对含水量为40%±5%;重度干旱胁迫(HD):土壤相对含水量为20%±5%。对照(CK):土壤相对含水量为80%±5%。每处理10盆。土壤相对含水量以土壤含水量占土壤饱和含水量的百分数表示[6]。试验环境温度为(25±5)℃;当控水达到预设的水分梯度后,每天18:00补充当天水分蒸发量。
1.3 测定指标与方法
1.3.1 土壤体积含水量日变化动态监测 使用托普云农土壤墒情速测仪(TZS-1K,中国)进行长期监测,每种各处理随机选择3盆,测量深度约10 cm,每隔30 min自动采集1次数据。
1.3.2 新生枝条相对生长量的测定 每隔15 d于9:00使用毫米刻度尺准确定位测定,计算相对生长量。相对生长量=(胁迫处理一定时间后新梢长度-初始新梢长度)/初始新梢长度×100%。
1.3.3 气体交换参数的测定 每隔15 d于 9:30-11:30使用便携式光合仪(LI-6400 XT,美国)测定叶片净光合速率(Pn,μmol·m-2·s-1),各处理随机选择3株,每株选择上端5~6片健康叶片待测,测定重复9次。
1.3.4 叶绿素含量的测定 叶绿素含量参考高俊凤[15]的方法,使用分光光度计(Eppendorf Bio Spectroneter fluorescence,德国)进行测定,采样间隔为15 d,每株选择3片叶待测,测定重复3次。
1.3.5 叶绿素荧光的测定 梨砧木叶片叶绿素荧光使用便携式光合仪(LI-6400 XT,美国)测定。每隔15 d在各植株上分别选择3片健康叶片作为待测叶片,重复3株,于测定当天15:00左右用锡箔纸包裹叶片使其充分暗适应约9 h后,照射检测光测定最小初始荧光F0,暗适应下最大荧光Fm,计算光系统Ⅱ中最大光化学效率[Fv/Fm=(Fm-F0)/Fm]。第2天9:00用叶室内活化光活化30 min以上,进行相关参数测定。
1.4 数据统计与分析
用Excel 2013和SAS v 9.0(SAS Institute Inc.,美国)对试验数据进行制图和统计分析。
2 结果与分析
2.1 不同程度干旱胁迫处理下土壤体积含水量动态变化
图1结果表明,正常供水和轻度干旱胁迫下土壤水分日变化波动较大,在12:00之后受植物蒸腾耗水及水分蒸发影响,土壤含水量出现明显下降,但盆内土壤含水量均处于试验设计范围内;而中度及重度干旱胁迫下土壤水分无明显变化。
2.2 不同程度干旱胁迫处理对2种梨砧木幼树生长的影响
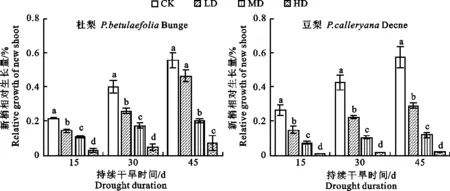
图2显示,正常供水情况下豆梨新梢相对生长量高于杜梨(P<0.05),说明豆梨幼苗较杜梨生长快。随着干旱胁迫的加重,2种砧木新梢相对生长量逐渐降低。持续性轻度干旱胁迫下,杜梨幼苗的新梢相对生长量下降了9.22%,但差异不显著(P>0.05),而豆梨降低28.47%(P< 0.05),显著低于对照;中度干旱分别使杜梨和豆梨新梢相对生长量降低35.52%和45.71%,差异达到显著水平(P<0.05);重度干旱下杜梨和豆梨的新梢相对生长量均显著低于其他水分条件,但杜梨新梢相对生长量为7.18%,豆梨新梢相对生长量仅为2.14%,说明中度及重度干旱严重影响梨砧木幼苗的生长,而豆梨受干旱胁迫影响 较重。

CK.正常供水 Normal water supply;LD.轻度干旱 Lighter drought-stress;MD.中度干旱 Moderate drought-stress;HD.重度干旱 Heavier drought-stress;下同 The same below
图1 不同程度干旱胁迫处理的土壤体积含水量
Fig.1 Soil volumetric moisture content under treatments of different drought stresses
不同小写字母表示差异显著(P<0.05) Different lowercase letters indicate significant difference(P<0.05)。下同 The same below
图2 干旱胁迫处理对2种梨砧木幼苗生长的影响
Fig.2 Effects of drought stress on growth of two kinds of pear rootstock seedlings
2.3 不同程度干旱胁迫对2种梨砧木净光合速率的影响
图3显示,与正常供水处理相比,干旱胁迫导致杜梨和豆梨幼苗净光合速率(Pn)下降,且不同程度干旱处理间存在差异。轻度干旱处理前期杜梨Pn值与正常供水植株相比,无明显差异(P>0.05),第45天时出现显著降低(P<0.05),而豆梨则从15 d时就低于对照(P<0.05);中度及重度干旱胁迫下2种砧木的Pn值均显著低于对照(P<0.05),其中杜梨45 d时较干旱处理初Pn值分别下降36.8%和74.5%,但豆梨分别下降 44.81%和78.04%,即在相同处理时间下杜梨的Pn降幅较小。
2.4 不同程度干旱胁迫对2种梨砧木叶片叶绿素和类胡萝卜素含量的影响
由表1可知,在正常供水情况下,豆梨的叶片叶绿素a(Chla)、叶绿素b(Chlb)、总叶绿素(Chl(a+b))及β-胡萝卜素(β-carotene)含量均高于杜梨,但差异不显著(P>0.05)。与正常供水相比,不同程度干旱胁迫下杜梨Chlb含量在30 d内变化均未达显著水平,在45 d时出现显著下降 (P<0.05),其他色素含量均呈不断下降的趋势;在重度干旱处理45 d时,杜梨叶片的Chla、Chlb、Chl(a+b)及β-carotene含量分别下降41.9%、27.3%、39.2%和35.4%,而豆梨分别降低了 58.6%、43.3%、55.3%和44.0%,表明在干旱胁迫下,豆梨色素含量下降幅度明显大于杜梨。
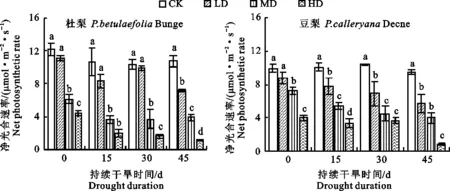
图3 干旱胁迫对2种砧木叶片净光合速率(Pn)的影响Fig.3 Effects of drought stress on net photosynthetic rate in leaves of two kinds of pear rootstock seedlings
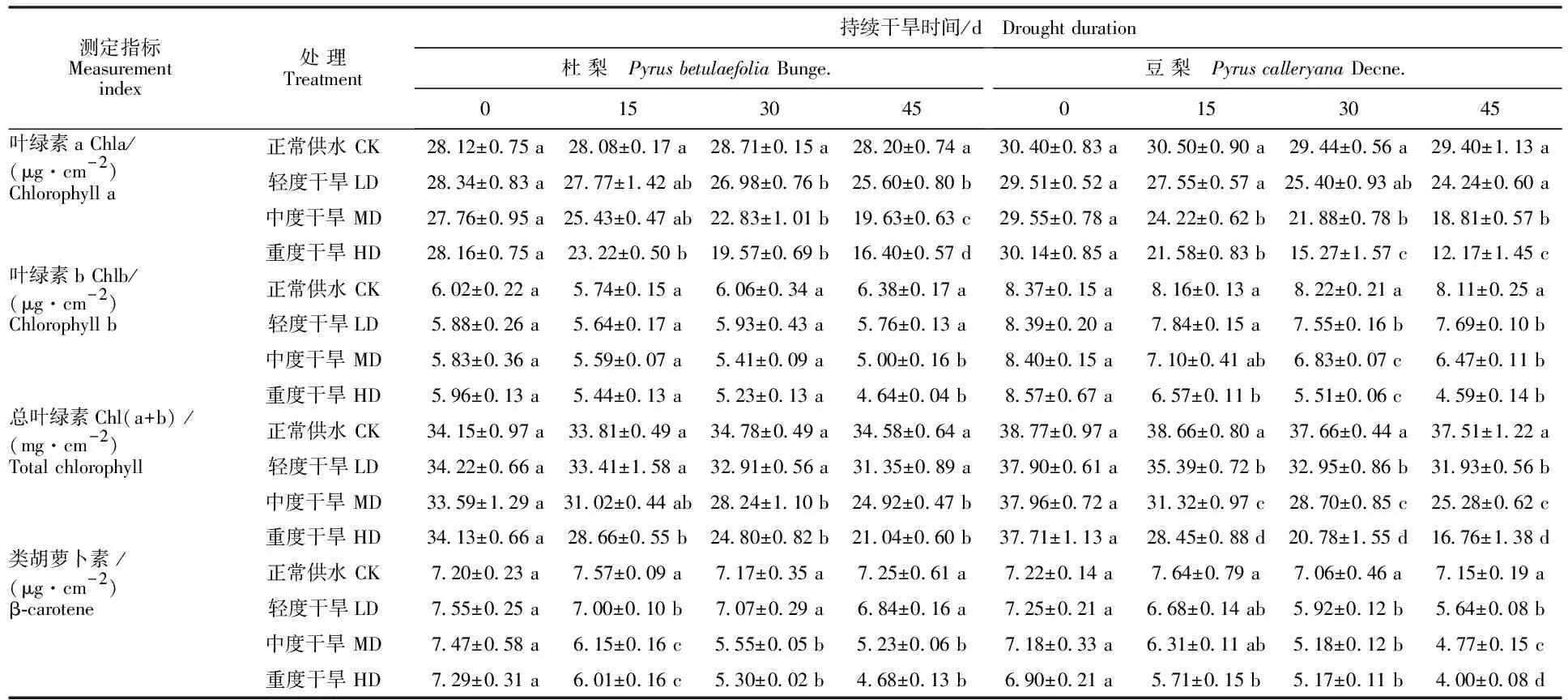
测定指标Measurement index处 理Treatment持续干旱时间/d Drought duration杜 梨 Pyrus betulaefolia Bunge.0153045豆 梨 Pyrus calleryana Decne.0153045叶绿素a Chla/(μg·cm-2)正常供水 CK28.12±0.75 a28.08±0.17 a28.71±0.15 a28.20±0.74 a30.40±0.83 a30.50±0.90 a29.44±0.56 a29.40±1.13 aChlorophyll a轻度干旱LD28.34±0.83 a27.77±1.42 ab26.98±0.76 b25.60±0.80 b29.51±0.52 a27.55±0.57 a25.40±0.93 ab24.24±0.60 a中度干旱 MD27.76±0.95 a25.43±0.47 ab22.83±1.01 b19.63±0.63 c29.55±0.78 a24.22±0.62 b21.88±0.78 b18.81±0.57 b重度干旱 HD28.16±0.75 a23.22±0.50 b19.57±0.69 b16.40±0.57 d30.14±0.85 a21.58±0.83 b15.27±1.57 c12.17±1.45 c叶绿素b Chlb/(μg·cm-2)正常供水 CK6.02±0.22 a5.74±0.15 a6.06±0.34 a6.38±0.17 a8.37±0.15 a8.16±0.13 a8.22±0.21 a8.11±0.25 aChlorophyll b 轻度干旱LD5.88±0.26 a5.64±0.17 a5.93±0.43 a5.76±0.13 a8.39±0.20 a7.84±0.15 a7.55±0.16 b7.69±0.10 b中度干旱 MD5.83±0.36 a5.59±0.07 a5.41±0.09 a5.00±0.16 b8.40±0.15 a7.10±0.41 ab6.83±0.07 c6.47±0.11 b重度干旱 HD5.96±0.13 a5.44±0.13 a5.23±0.13 a4.64±0.04 b8.57±0.67 a6.57±0.11 b5.51±0.06 c4.59±0.14 b总叶绿素Chl(a+b) /(mg·cm-2)正常供水 CK34.15±0.97 a33.81±0.49 a34.78±0.49 a34.58±0.64 a38.77±0.97 a38.66±0.80 a37.66±0.44 a37.51±1.22 aTotal chlorophyll轻度干旱LD34.22±0.66 a33.41±1.58 a32.91±0.56 a31.35±0.89 a37.90±0.61 a35.39±0.72 b32.95±0.86 b31.93±0.56 b中度干旱 MD33.59±1.29 a31.02±0.44 ab28.24±1.10 b24.92±0.47 b37.96±0.72 a31.32±0.97 c28.70±0.85 c25.28±0.62 c重度干旱 HD34.13±0.66 a28.66±0.55 b24.80±0.82 b21.04±0.60 b37.71±1.13 a28.45±0.88 d20.78±1.55 d16.76±1.38 d类胡萝卜素 /(μg·cm-2)正常供水 CK7.20±0.23 a7.57±0.09 a7.17±0.35 a7.25±0.61 a7.22±0.14 a7.64±0.79 a7.06±0.46 a7.15±0.19 aβ-carotene 轻度干旱LD7.55±0.25 a7.00±0.10 b7.07±0.29 a6.84±0.16 a7.25±0.21 a6.68±0.14 ab5.92±0.12 b5.64±0.08 b中度干旱 MD7.47±0.58 a6.15±0.16 c5.55±0.05 b5.23±0.06 b7.18±0.33 a6.31±0.11 ab5.18±0.12 b4.77±0.15 c重度干旱 HD7.29±0.31 a6.01±0.16 c5.30±0.02 b4.68±0.13 b6.90±0.21 a5.71±0.15 b5.17±0.11 b4.00±0.08 d
注:同列不同小写字母表示同一指标在相同处理时间内不同干旱处理间差异显著(P<0.05)。
Note:Different lowercase letters in the same columns indicate significant difference among different drought treatments at the same treatment time at 0.05 level.
2.5 不同程度干旱胁迫对2种梨砧木叶绿素荧光参数的影响
由图4可知,在正常供水情况下,2个品种间光系统Ⅱ(PSⅡ)荧光参数无明显差异(P> 0.05),干旱胁迫前期(0 d、15 d)对于杜梨的最小初始荧光(F0)、实际光化学量子效率(ΦPSⅡ)、非光化学猝灭系数(NPQ)和电子传递效率(ETR)均无明显影响,持续的低土壤含水量条件下,2个品种的最大光化学效率(Fv/Fm)、ΦPSⅡ、光化学猝灭系数(qP)和ETR值均显著下降(P<0.05),F0和NPQ则显著升高(P<0.05)。
2.5.1 最大光化学效率(Fv/Fm) 由图4可知,干旱胁迫后杜梨和豆梨的Fv/Fm均有所减小,且胁迫越重,降幅越大。轻度干旱胁迫下杜梨Fv/Fm在第45天时相较于CK下降1.33%,豆梨则是在胁迫第30天已经降低2.06%;中度和重度水分胁迫下杜梨、豆梨分别在第30天和第15天时出现下降;重度胁迫处理45 d时,杜梨和豆梨相较于CK分别降低6.67%和7.31%。
2种梨砧木的F0在胁迫后均有所增加,增加幅度在各处理间均存在显著差异。在轻度、中度和重度干旱处理15 d时,豆梨F0均显著高于CK,之后随时间延长,差异愈加显著,45 d时F0分别增加至CK的1.1倍、1.25倍和1.36倍;而在45 d时,轻度胁迫处理下的杜梨与CK相比无显著差异,中度和重度干旱使杜梨F0增加至对照的1.06倍和1.11倍。

图4 干旱胁迫对2种砧木叶绿素荧光参数的影响Fig.4 Effects of drought stress on chlorophyll fluorescence parameters of two kinds of pear rootstock seedlings
2.5.2 荧光猝灭动力学 由图4可知,干旱胁迫下,杜梨和豆梨的ΦPSⅡ相较于正常供水处理均呈下降趋势(P<0.05),其中,中度及重度胁迫下杜梨从第30天开始显著低于正常供水(P<0.05),而豆梨则在第15天即出现显著下降。
qP为光适应光化学淬灭系数,可以反映光合反应中心的开放程度。由图4可知,轻度干旱胁迫下,杜梨qP值在45 d时显著降低,豆梨在15 d时出现显著下降;在中度和重度干旱胁迫下杜梨和豆梨均于第15天出现下降。与CK相比,长期轻度、中度和重度干旱处理45 d使得杜梨qP分别下降14.4%、34.3%和36.5%,而豆梨则分别下降 23.7%、35.8%和46.3%。
NPQ可反映PSⅡ吸收的能量以热耗散形式释放的部分。由图4可知,随着土壤水分含量降低及处理时间延长,杜梨和豆梨的NPQ值均显著上升(P<0.05)。干旱处理45 d时,轻度、中度和重度干旱使杜梨的NPQ值相较于CK分别增长 21.7%、42.7%和68.9%,而使豆梨分别增长12.0%、 42.4%和53.0%。
轻度干旱胁迫对于杜梨的电子传递效率(ETR)无明显影响,但豆梨却呈下降趋势,差异显著(P<0.05)。在中度和重度干旱胁迫下2种砧木的ETR表现相似的变化规律,即均呈显著下降趋势。
3 讨论与结论
3.1 干旱胁迫对梨砧木幼苗生长特性的影响
杜梨和豆梨幼苗的水分消耗和生长情况均与土壤体积含水量紧密相关(图1,图2)。水分亏缺下2种梨砧木新梢相对生长量均低于正常供水处理。干旱条件下植株的新梢相对生长量是其抗旱性反应的综合体现,本研究发现,杜梨对轻度干旱胁迫适应性较强,该结果与李泽等[16]的研究结果类似;随土壤水分含量的降低,杜梨和豆梨幼苗的生长受抑制程度逐渐加重,新梢生长愈加减缓,长期干旱条件下豆梨的新梢生长速率明显低于杜梨,换言之,杜梨表现出较好的抗旱能力。
3.2 干旱胁迫对梨砧木幼苗叶片净光合速率的影响
梨砧木在应对干旱胁迫时表现形态和生理变化等多种防御机制[17]。本研究发现干旱胁迫后杜梨叶绿素b含量变化较小,但随着干旱胁迫的加剧,2种梨砧木叶绿素a、总叶绿素及类胡萝卜素含量均不断降低,这可能是由于水分亏缺使活性氧累积,器官受损,影响叶绿素的生物合成,同时叶绿素分解加快,从而导致植株叶片叶绿素减少[8];相较于叶绿素b而言,叶绿素a对活性氧反应更敏感,致使叶绿素a含量受干旱胁迫影响较大[18]。梨砧木幼苗的叶绿体结构破坏,直接影响光合作用的正常进行,因此干旱胁迫以对光合作用的影响最为突出。井大炜等[19]、Limbu等[20]研究发现,随干旱胁迫程度的增加,植物叶片的Pn随之下降,抗旱性强的植物Pn较抗旱性弱的植物降幅小。本研究结果同样表明,重度和中度干旱胁迫下2种砧木幼苗的光合作用明显减弱,这可能主要是由于渗透胁迫导致叶片气孔关闭,环境中CO2向细胞内扩散难度增大,使得光合碳固定底物减少所致。杜梨的光合参数在干旱胁迫后变化幅度明显小于豆梨,表现出较好的抗旱能力,可能与杜梨在胁迫后能较好地维持正常叶片气孔形态相关。
3.3 干旱胁迫对梨砧木幼苗光系统Ⅱ叶绿素荧光参数的影响
植物叶绿素所吸收的光能需经叶绿素荧光发射、光合电子传递和热耗散消耗,因此,利用叶绿素荧光特性能够有效探测植株的光合作用效率[21]。逆境中植物品种内或品种间遗传物质的差异会使其叶绿素荧光参数从本质上产生不同[22-25]。Fv/Fm响应逆境胁迫较为敏感[26],本试验中杜梨和豆梨的Fv/Fm均逐渐下降,表明长期水分亏缺使梨砧木叶片发生了光抑制[27-28]。F0既和叶绿素含量有关,同时也与PSⅡ反应中心的活性密切相关[29-30]。前人研究发现,干旱[31]和盐胁迫[32]等逆境均使F0升高。本研究发现,土壤含水量降低使豆梨的F0增幅显著大于杜梨,说明水分亏缺使豆梨光合器官受到更严重损伤,影响PSⅡ的正常活动。杜梨的F0在不同程度干旱处理30 d时才逐渐升高,而豆梨在15 d就发生了显著升高,证实杜梨可维持PSⅡ活性以增强对干旱的耐受性。
Jalink等[22]、Razavi等[33]认为,植物的抗旱能力与非光化学猝灭系数NPQ的上升幅度呈正比,此系数升高表明植株在干旱胁迫下会启动自我保护机制。本研究结果显示,干旱胁迫45 d后杜梨qP和ETR下降幅度低于豆梨,NPQ增加幅度明显大于豆梨,这可能是由于在严重水分亏缺的情况下,抗旱性强的砧木杜梨一方面可以保护光系统Ⅱ,维持原初光能转换效率和潜在活性的稳态,另一方面非辐射热能耗散体系能够有效响应干旱,从而使光合机构得到保护。
综上所述,干旱胁迫使2种梨砧木均受到光抑制,叶片叶绿素含量下降,叶绿体结构发育受阻,净光合速率减弱,从而导致生长减缓,但杜梨的光合系统对干旱胁迫的防卫能力明显优于豆梨,轻度干旱胁迫对杜梨不产生明显影响。
猜你喜欢
绿色科技(2021年21期)2021-11-26 09:12:06
内蒙古林业调查设计(2021年3期)2021-07-06 14:12:00
落叶果树(2021年6期)2021-02-12 01:29:12
北方果树(2020年6期)2020-11-14 01:35:42
现代园艺(2018年1期)2018-03-15 07:56:13
现代园艺(2017年21期)2018-01-03 06:41:45
作文周刊·小学二年级版(2017年11期)2017-06-21 03:47:16
湖南农业(2017年1期)2017-03-20 14:04:46
爱你·阳光少年(2016年10期)2016-05-30 09:06:07
儿童绘本(2015年4期)2015-05-25 17:59:48
