
拉萨河中上游夏秋季纤毛虫群落时空变动及其与环境的关系
2019-06-04 07:31杨欣兰
生态学报 2019年9期
杨欣兰,巴 桑,*,黄 香
1 西藏大学理学院青藏高原湿地与流域生态实验室,拉萨 850000 2 西藏大学理学院化学与环境科学系,拉萨 850000
纤毛虫广泛分布于淡水、海水及潮湿的土壤中,在微型生物食物网中物质循环与能量流动传递中起着重要的枢纽作用[1-2]。该类生物的物种组成及功能结构的变动直接影响到水生态系统中食物网的组成及其他水生生物的分布和丰度[3-4]。因其繁殖周期简单而短暂,加之对水环境的变化敏感,许多种类已广泛用作水环境污染的指示者[5-6]。因此,揭示纤毛虫的群落变化规律对研究各类河流等水域的自净效能及污染评价均具有重要的意义[7]。
尽管国内外有关种类水体原生动物群落结构的研究已有大量的报道[8-9],但是关于青藏高原纤毛虫分类学组成及群落结构变动及与环境间的关系的报道甚少,尤其有关拉萨河中上游纤毛虫的研究尚处完全不详之中[10-12]。本工作于2015—2016年8月、11月分别对拉萨河中上游纤毛虫群落结构的时空变化特征与水环境之间的关系进行了研究(2016年的纤毛虫数据为2015年纤毛虫的补充数据),旨在揭示高原河流有关纤毛虫群落结构时空变化及与环境间的耦合关系,为了我国青藏高原水域环境监测与保护提供基础数据和科学依据。
1 材料和方法
1.1 样区概况
拉萨河是雅鲁藏布江五大一级支流之一,发源于念青唐古拉山的南麓、澎错东南约15 km的澎错扎玛朵山峰下。拉萨河干流全长约550 km,流域面积32896 km2,平均海拔在4 km以上。径流丰富,整个水系呈现羽毛状。主要支流有:桑曲、拉曲、雪绒藏布、墨竹玛曲和堆龙曲等。根据河谷形态和地质情况,拉萨河大致分为上、中、下游三段,从源头到桑曲汇入口为上游段。受地势的影响,气候寒冷,为高原大陆型气候。流域内人类活动相对稀少,河流水环境基本保持原生态。从桑曲汇入口到旁多拉曲汇入口为中游段,称为热振藏布,地处高原温带半干旱季风气候区[13]。该流域的主要补给水源为冰川融雪水和天然降水。
1.2 样点设置
根据拉萨河中上游的自然环境特征及水体功能的特殊性,共设置8个样点,其中S1、S2、S5和S8为拉萨河主干流,地处在高山草甸、灌木和农田生态系统,S3和S4为拉萨河一级支流——拉曲,地处高山草甸和高寒草甸生态系统;S6和S7为拉萨河一级支流——雪绒藏布,地处灌木和农田生态系统。流域内水体除S2(水库静水)河床为草甸外,其余都是砾石和砂石组成的山区稳定性河流,坡度较大,水流速度快(图1)。
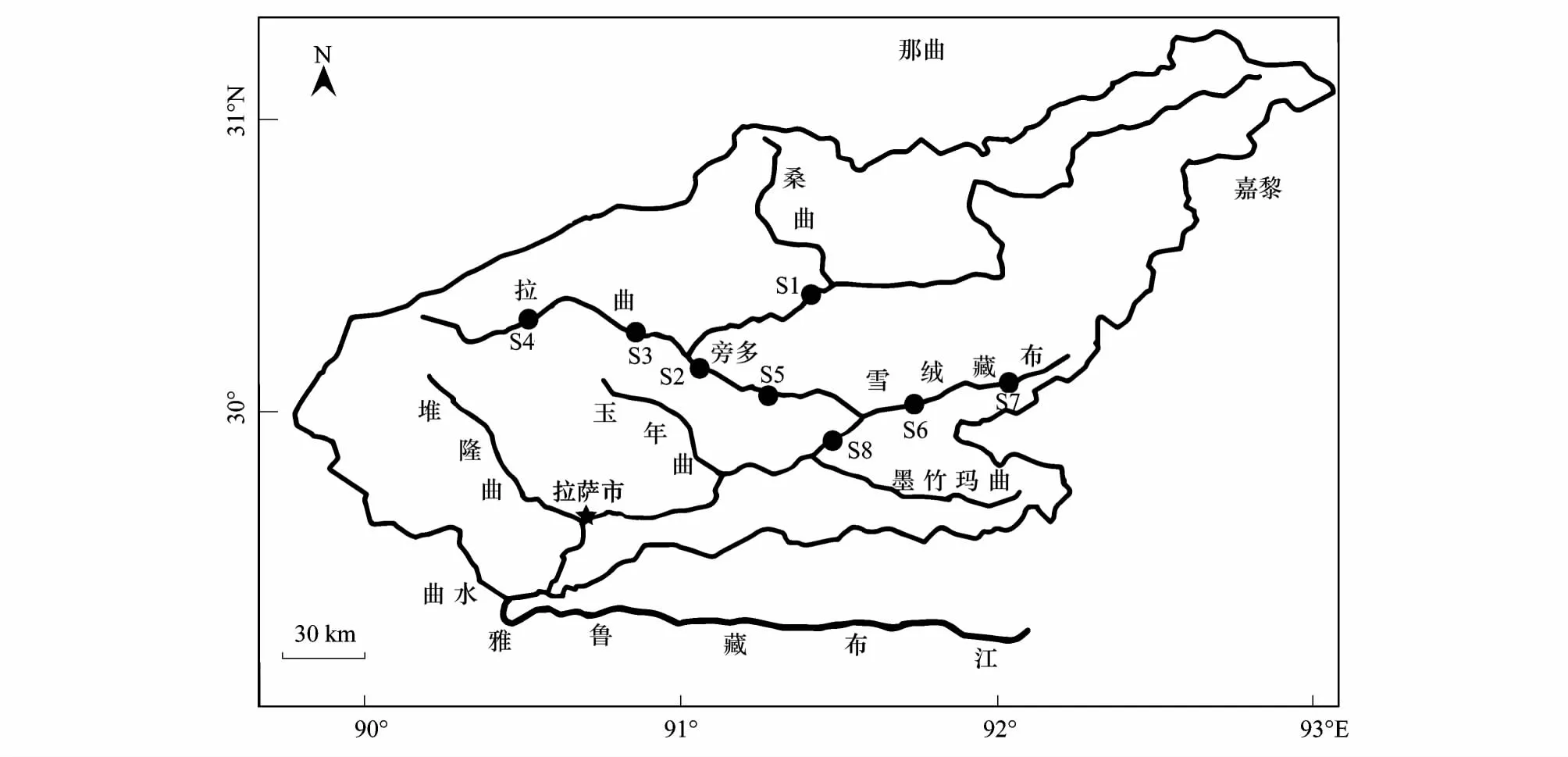
图1 拉萨河中上游样点分布图Fig.1 Sampling sites in middle and upper reaches of the Lhasa RiverS:样点,Site
1.3 样品的采集与处理
本次研究分别于2015年8月1日和10月30日与2016年8月3日和11月1日进行样品采集,每次采集时间为一周。样品采集及室内处理方法均参照《淡水浮游生物研究方法》进行[14]。定性样品用25号浮游生物网(孔径64 μm),在水面下0.8—1.2 m作∞字型拖网8—10 min;定量样品在上述深度采集1 L,随后用10%鲁哥氏液固定,静置24 h后沉淀浓缩至50 mL。移取0.1 mL浓缩液至0.1 mL计数框,在10×40倍显微镜下全片计数纤毛虫数量(平行3次)[15]。
水体pH值、水温、电导率、总溶解盐、盐度现场利用PCSTestr-35型仪器(新加坡产)测定;溶解氧采用HI-9146便携式溶解氧测定仪(德国产)测得;浊度用HI-98703高精度数据型浊度测定仪(德国产)测得;水流速度采用FP-111直读式流速仪(美国产)测得;总氮、总磷、氨氮和化学需氧量测定参照水和废水监测分析方法(第四版)和地表水环境质量标准(GB 3838—2002)于室内完成。
1.4 分类与鉴定
用活体观察、鲁哥氏液染色固定相结合的方法鉴定物种[2];鉴定依据下列文献:《原生动物学》[2],《淡水微型生物与底栖动物图谱》[16],《中国黄渤海的自由生纤毛虫》[17],《西藏水生无脊椎动物》[18]。采用Levine 分类系统进行分类地位的划分[19]。
1.5 数据统计与分析
1.5.1类群划分
对已鉴定到的各级分类单元及物种进行统计,按目级水平将物种最多的分类单元定义为优势类群,将物种次多的分类单元定义为次优势类群,将单物种或含最少种类的分类单元定义为偶见类群, 其余的为常见类群[20]。
1.5.2群落参数
用Excel计算了Shannon-Wiener多样性指数、Simpson优势度指数、Pielou 均匀度指数;用SPSS 23.0对纤毛虫群落特征参数与水体理化因子进行了Pearson相关性分析;用R软件对纤毛虫优势物种与环境因子进行了CCA 典范对应分析。根据各样点纤毛虫出现的频率和相对丰度,依据McNaughton优势度系数Y≥0.02时定为优势种。上述群落参数公式如下:
(1)Shannon-Wiener多样性指数:H=-∑Pi×lnPi(Pi=Ni/N)
(2)Simpson优势度指数:S=∑Ni(Ni-1)/(N(N-1))
(3)Pielou 均匀度指数:E=H/lnNs
(4)McNaughton优势度系数:Y=fi×Ni/N
式中,N为同一样点中个体总数;Ni为第i种的个体数;Ns为物种数;其中fi为第i种出现的频率[21-25]。
1.5.3营养功能类群划分
按照Pratt和Cairns的方法,将纤毛虫按食性分成6个营养功能类群[26],即光合自养者(P群)、食藻者(A群)、食菌—碎屑者(B群)、腐养性(S群)和食肉者(R群)。若群落中B群和S群占的比例较大,则反映水质较差[3]。
2 结果
2.1 水体理化因子
通过数据分析得出:水流速度、COD、NH4-N 和NO3-N在两个季节不存在显著性差异,其余理化因子在两个季节均存在显著性差异,详见表1。

表1 拉萨河中上游夏、秋季水环境理化因子
WT:水温,Water Temperature;TUR:浊度,Turbidity;EC:电导率,Electrical Conductivity; TDS:总溶解盐,Total Dissolved Salts; Sal:盐度,Salinity;DO:溶解氧,Dissolved Oxygen; COD:化学需氧量,Chemical Oxygen Demand; TP:总磷,Total Phosphorus; TN:总氮,Total Nitrogen;NO3-N:硝酸盐氮,Nitrate nitrogen; NH4-N:氨氮,Ammonia Nitrogen;WS:水流速度,Water Speed
2.2 群落结构及优势物种
2.2.1群落结构
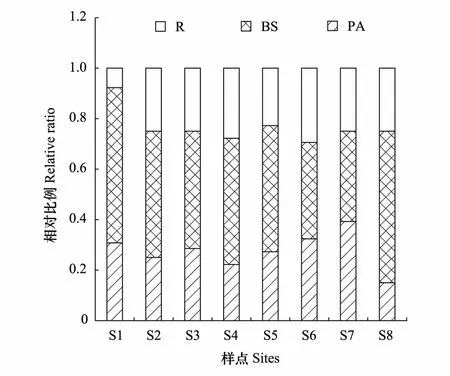
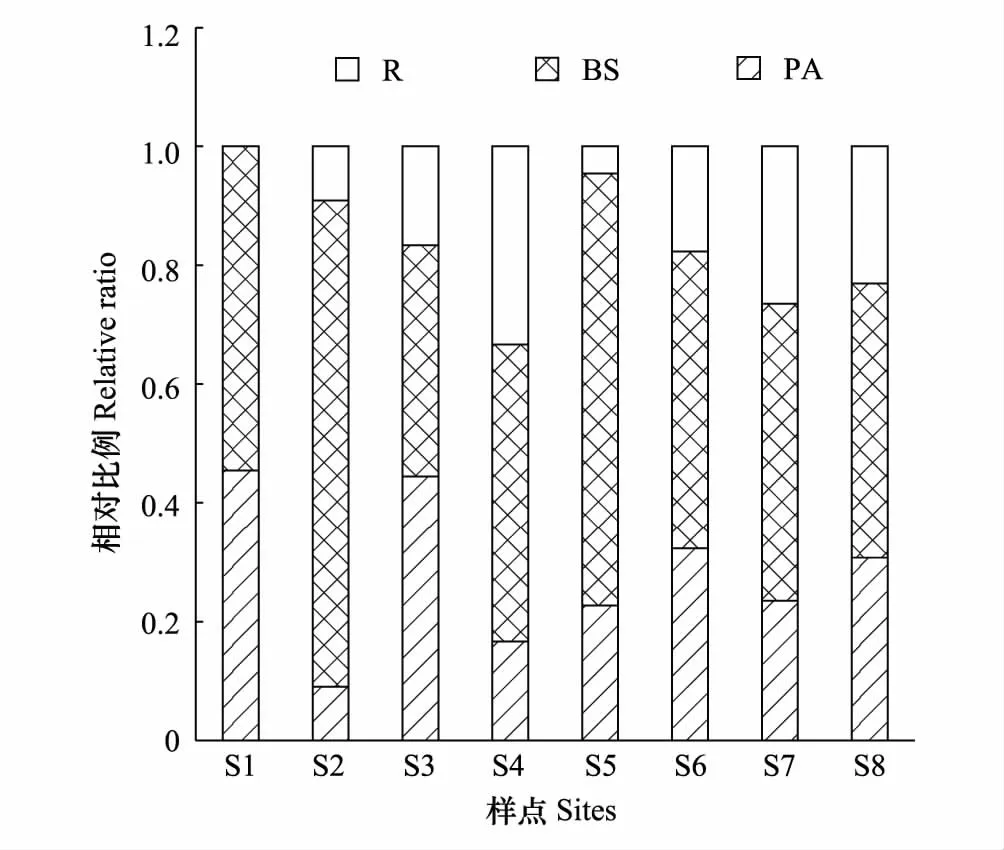
拉萨河中上游夏、秋季共鉴定出纤毛虫91种。夏季49种,隶属于3纲11目19科26属,其中,前口目为优势类群(22.45%);侧口目(14.29%)、膜口目(14.29%)和盾纤目(14.28%)为次优势类群;寡毛目(2.04%)为罕见类群,其余为常见类群。各样点纤毛虫物种数由小到大依次为:S2 表2 拉萨河中上游纤毛虫分类学组成 2.2.2优势种 夏、秋季的优势种数均为12种,但是优势种的组成及分布不同。其中,大弹跳虫、多变斜口虫和前突肾形虫是两个季节共有的种;两个季节中S4样点的优势度均值最高(0.745),S5样点的优势度均值最低(0.302);夏季的优势度均值(0.34)略高于秋季(0.18)(表3)。 表3 拉萨河中上游纤毛虫优势种 物种序列用季节的拼音首字母加数字依次排序 拉萨河中上游夏季Shannon多样性指数S6样点最高,S4最低;秋季的Shannon多样性指数S7样点最高,S4最低;总体夏季Shannon多样性指数(1.97)略高于秋季(1.86),但无显著性差异(t=0.6632,df=14,P=0.52)。夏季Pielou均匀度指数S2样点最高,S8最低;秋季Pielou均匀度指数S4最高,S3最低,总体两个季节的均值相同(0.81),无显著差异(U=32.5,P=1);夏季Simpson优势度指数S4样点最高,S2最低;秋季Simpson优势度指数S3最高,S4最低,总体两个季节的均值相同(0.22),无显著性差异(t=0.12329,df=14,P=0.9038)(表4)。 表4 拉萨河中上游纤毛虫群落特征参数 营养功能结构分析显示:B、S类群和P、A类群在两个季节的各样点中存在一定的差异。夏季B、S类群种类数(22)所占总体物种数比例(44.90%)明显高于P、A类群(20.41%);秋季B、S类群种类数(26)占的物种数比例(40.63%)明显高于P、A类群(18.75%)(图2、3)。 图2 拉萨河中上游夏季纤毛虫营养功能结构变动 Fig.2 Variation in functional-trophic structure of ciliate communities in middle and upper reaches of the Lhasa River in summerP:光合作用者,Photosynthetic autotrophs;A:食藻者,Algivores;B:食菌-碎屑者,Bactivores-detrivores;S:腐养性,Saprotrophs;R:食肉者,Predalors 图3 拉萨河中上游秋季纤毛虫营养功能结构变动 Fig.3 Variation in functional-trophic structure of ciliate communities in middle and upper reaches of the Lhasa River in autumn 通过纤毛虫群落参数与水体理化因子作Pearson双尾相关性分析可知:夏季,纤毛虫群落Pielou均匀度指数与水流速度呈显著负相关(P<0.05),纤毛虫物种数与总磷和总氮均呈显著负相关(P<0.05);秋季,纤毛虫物种数和群落Shannon多样性指数与浊度、氨氮和硝酸盐氮呈显著负相关(P<0.05),与pH呈显著正相关(P<0.05)(表5)。 为了进一步阐释和验证拉萨河中上游纤毛虫群落参数与环境因子之间的关系,本文筛选出5项主要水体理化指标,分别对两个季节的优势种与环境因子进行了CCA 分析。结果显示:总磷和浊度是影响夏季纤毛虫优势种的关键因子;水流速度、浊度和总氮是影响秋季纤毛虫优势种的关键因子(图4、5)。 图4 基于CCA方法分析拉萨河中上游夏季纤毛虫群落与环境因子之间的关系Fig.4 CCA analysis: showing relationships between ciliate community structure and environment factors in middle and upper reaches of the Lhasa River in summer 图5 基于CCA方法分析拉萨河中上游秋季纤毛虫群落与环境因子之间的关系Fig.5 CCA analysis: showing relationships between ciliate community structure and environment factors in middle and upper reaches of the Lhasa River in autumn 本文对拉萨河中上游两个季节的主要水体理化因子进行分析,并参照地表水环境质量标准《GB 3838—2002》分析得出,总磷和总氮的浓度能达到地表水环境质量标准,并且该河段水体水质良好,属于Ⅰ—Ⅲ 类水。其原因是拉萨河中上游地处青藏高原,人口稀薄,且中上游无大中型重工业,对水环境的人为干扰很少[27]。这也是环境监管与环境卫生有关部门的协调合作,政府有关环境保护政策的计划与实施,使拉萨河独有的高原生态环境得到更为有效的保护和可持续性发展的结果。 本研究中根据地理条件的特征设置了8个样点,能够充分的监测出拉萨河中上游夏秋季节的纤毛虫物种组成与丰度,进一步反应水体的状况。共鉴定出的91种纤毛虫,其中49种见于夏季,64种见于秋季。夏季各样点丰度为1.2×104—5.6×105个/L,秋季各样点丰度在1.2×104—2.6×105个/L之间,并且各样点的物种数与丰度不尽相同。可见,拉萨河中上游夏、秋季各样点的物种组成与丰度存在一定的差异。这可能与青藏高原的极端地理环境有关,即拉萨河属于冷水性河流,长期的自然进化使水生纤毛虫适应了在较低温度环境下的生存,随着季节的更替,水温降低导致纤毛虫的种类增多。这与西藏及内陆其他地区冷水型河流纤毛虫的研究结果相一致[28-31],但与其他非冷水型河流的研究结果相背[32]。另外,拉萨河中上游的物种数整体较国内相关河流偏低,说明水中营养物质相对较少,外源营养来源较单一,这也进一步说明青藏高原河流贫营养型的水环境特征。 表5 拉萨河中上游纤毛虫群落参数与水体理化因子间的相关性 **. 在 0.01 级别相关性显著;*. 在 0.05 级别相关性显著;**:P<0.01; *:P<0.05 纤毛虫能够适应的种类水环境,并形成相对稳定的群落结构。一旦环境发生变化,该群落结构的稳定性就会受到直接或间接的影响[5]。通过两季节结构参数的对比分析得出:夏季的Shannon多样性指数值与Simpson优势度指数值均比秋季的高,Pielou均匀度指数比秋季的低。秋季纤毛虫群落较夏季的结构简单,优势种明显,物种分布较夏季不均匀。其主要原因为秋季随着雨季过去水温下降,流域内水流量减少、流速缓慢,残留的有机物在河水中降解并富集,河岸植物腐败沉降并富集,增加了水中的外来营养源,导致个别耐低温与耐高盐的纤毛虫物种数增加[33-34],从而影响水中纤毛虫的群落多样性;另一方面来说,夏季水流速度较大、水温较高,流水卷起河床的氮磷等营养物质,有利于纤毛虫的生存和繁衍,从而增加了纤毛虫的数量[35-36],形成秋季纤毛虫较夏季纤毛虫优势种明显,群落结构简单,物种分布不均匀的现象。 拉萨河中上游纤毛虫营养功能结构分析显示,夏、秋两季B、S类群和P、A类群及R类群的结构模式没有明显的变化,但是夏季B、S类群的相对种类数低于秋季。这表明,拉萨河中上游水质并未受到明显的外源干扰,整体水质状况良好,符合理化指标显示的结果,并且夏季水质优于秋季[37]。这与该地区的气候环境和农业生产方式有密切关系。长期的高寒缺氧气候使西藏的植被在10 月初就开始枯萎,河流内的植物也比其他地区更早的失去其自养净化功能,使得大量的有机物在流域内残存[38],致使河流水质有所下降。另外,该地区为纯牧区,大量的牛、羊等牲畜长期在河岸周围放牧,饮取河流中水源的同时也把相应的污染物带入了该流域内[39],这也是导致拉萨河中上游秋季水质较夏季下降的原因之一。 通过Pearson双尾相关性分析可知:夏季,纤毛虫群落Pielou均匀度指数与水流速度呈显著负相关,纤毛虫物种数与总磷和总氮均呈显著负相关;秋季,纤毛虫物种数和群落Shannon多样性指数与浊度、氨氮和硝酸盐氮呈显著负相关,与pH呈显著正相关。为了查证群落结构与环境之间的复杂关系,运用CCA典范对应分析来进一步解释两者之间的关系。其结果显示:影响夏季纤毛虫物种分布的关键因子是总磷和浊度;影响秋季纤毛虫物种分布的关键因子是水流速度和浊度。夏季由于雨季的到来水流量增大,流速增快,常见种数量下降,雨水将农田地表径流水带入河中,导致河水中营养物质增多,耐营养物质的种类增多;秋季,浊度、pH、总磷和总氮皆与物种数和多样性指数呈显著负相关。前人研究表明,浊度能够促进细菌的生长繁殖,从而对水中原生动物指标有明显的影响[40];细菌与原生动物繁殖率的增加引起微生物的代谢率的增高,从而影响水环境内的氮磷浓度。这是导致一些纤毛虫增多的重要原因。括而言之,影响纤毛虫群落结构是多个环境因子共同作用的结果,而不是某个单方面的因子单独作用的结果。 1)根据地表水环境质量标准《GB 3838—2002》分析拉萨河中上游水体理化因子,该流域春、夏季水体显示为Ⅰ—Ⅲ 类水质。 2)拉萨河中上游夏、秋季纤毛虫群落结构较稳定,物种分布均匀,生物多样性指数较低。 3)拉萨河中上游纤毛虫B、S类群、P、A类群、R类群的结构组成没有明显的季节变化趋势,但是夏季B、S类群物种丰富度低于秋季。 4)纤毛虫群落特征及结构参数可以作为水环境评价的重要指标。 5)水流速度、总磷和总氮是影响拉萨河中上游夏季纤毛虫群落结构的主要因素, pH、浊度、总磷和总氮是影响该流域秋季纤毛虫群落结构的主要因素,浊度是影响纤毛虫物种分布的主要因素。
2.3 群落特征参数

2.4 营养功能类群
2.5 Pearson双尾相关性
2.6 CCA结果分析
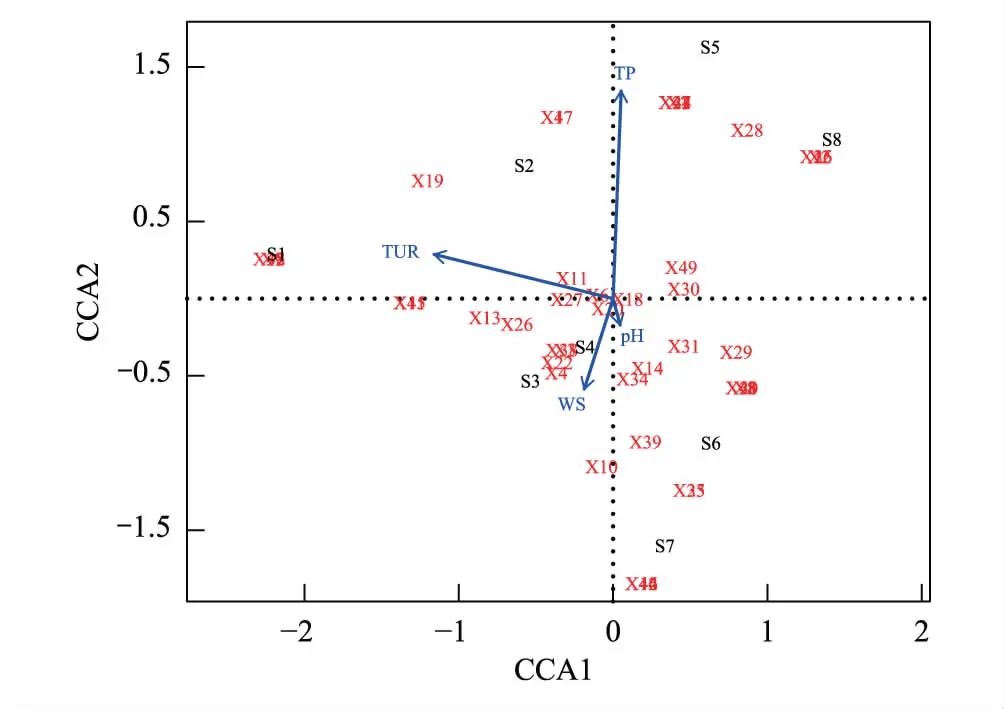

3 讨论
3.1 水体理化因子
3.2 纤毛虫物种组成与丰度

3.3 纤毛虫群落特征参数
3.4 营养功能类群
3.5 纤毛虫群落结构与环境间的关系
4 结论
猜你喜欢
海洋与湖沼(2022年5期)2022-09-21
湖北植保(2022年4期)2022-08-23
生态学报(2022年9期)2022-05-26
特区文学·诗(2021年6期)2021-12-22
土壤(2021年1期)2021-03-23
生物化工(2021年4期)2021-01-19
布达拉(2018年4期)2018-05-14
渔业科学进展(2018年1期)2018-04-03
土壤学报(2017年5期)2017-11-01
西藏大学学报(自然科学版)(2016年1期)2016-11-15
