
冬麦免耕覆盖栽培对土壤丛枝菌根真菌多样性的影响
2019-05-30 07:18:20汪志琴王小玲魏常慧
核农学报 2019年5期
汪志琴 马 琨 王小玲 李 越 魏常慧
(1 宁夏大学农学院,宁夏回族自治区 银川 750021;2 宁夏大学西北土地退化与生态恢复国家重点实验室培育基地,宁夏回族自治区 银川 750021)
丛枝菌根(arbuscularmycorrhizae,AM)真菌是自然界分布广泛的一种植物根系和真菌共生体[1],能通过碳和养分吸收、传递,强化生物和非生物生态系统组分之间的联系[2-4]。随着植物与微生物相互作用研究的深入,发现AM真菌能直接或间接地影响土壤生态过程;AM真菌是维持陆地生态系统稳定性的重要组分,在植物生长、土壤肥力与作物生产力形成等方面发挥着至关重要的作用[5-6]。因此,了解不同农业管理活动对AM真菌的影响,对农业生产和土壤的可持续利用具有重要意义。
免耕是农田管理活动中经常采用的一种保护性耕作措施。与传统耕作相比,免耕能通过减少土壤的扰动提高土壤微生物生物量和生物多样性,改变微生物群落组成,优化土壤环境,改善土壤质量,间接影响作物养分吸收,从而影响作物产量[7-8]。研究表明,AM真菌受耕作措施和施肥制度的影响,在扰动较少的土壤中能表现出更高的多样性[9]。与耕地相比,持久草地管理下土壤AM真菌具有更高的多样性,这2种利用方式下的土壤AM真菌群落组成和多样性存在明显差异,集约化农业生产管理活动会对AM真菌的增殖产生不利影响[10]。相对未开垦的土地而言,传统耕作由于破坏了AM真菌菌根网络,导致AM真菌丰富度、多样性均有所降低;而在集约化农业生产活动影响下,AM真菌的多样性通常更低、丰富度更小,甚至土壤在受到扰动后会导致某些物种消失[11]。然而,关于AM真菌群落对土地利用方式响应的研究结论不一。Plaza等[12]研究表明,土壤耕作扰动有利于少数AM真菌孢子快速生长;而高萍等[9]和Kautz等[13]则认为,有机耕作强度与土壤AM真菌群落结构及其多样性呈负相关关系,免耕条件下的土壤AM真菌多样性均高于耕作。
秸秆覆盖可以通过提高AM真菌生物量来促进植物对养分的吸收[14]。Bending等[15]研究发现在免耕秸秆覆盖措施下,土壤AM真菌多样性指数、土壤微生物碳源利用能力均显著提升;而植被清除、土壤耕作、化学肥料施用均显著降低了土壤AM真菌生物量;在高肥力下,土壤氮、磷、速效钾的增加是导致AM真菌群落组成变化的驱动因子。Dai等[16]和Köhl等[17]研究表明,农业生产中肥料的投入是导致AM真菌群落结构改变的主要因素。有机肥施用对土壤AM真菌生长繁殖有促进作用,这主要是因为有机肥可以通过改变土壤矿质营养浓度、植物组织氮磷比来影响AM真菌的生长和侵染能力,进而刺激AM真菌种群的生长,使其更适应新的营养条件。朱晨[18]也证实化学氮肥的施用对AM真菌群落结构与多样性均具有显著影响。氮肥施用水平的提高会导致AM真菌多样性的降低,进而导致AM真菌群落结构简单、功能单一,而磷肥施用则无显著影响;此外,施用有机肥或生物有机肥,对调控土壤微生物区系有突出作用,说明有机肥也是影响AM真菌的重要因素之一[19]。
土壤AM真菌在维持农田生产力方面发挥着不可替代的作用,研究农业综合管理活动对AM真菌群落组成、多样性以及不同AM真菌间功能差异的影响具有重要意义。但目前相关研究[16-19]多集中在单一土地管理要素对AM真菌的影响上,而关于AM真菌在免耕、覆盖、有机肥管理等综合因素及交互作用的响应上研究尚鲜见报道。本研究以不同农业综合管理活动的农田生态系统,包括传统耕作有机生产、免耕覆盖耕作有机生产的冬麦农田土壤为对象;利用基于Illumina MiSeq平台的高通量测序方法,旨在揭示耕作、施肥及覆盖三者之间交互关系对AM真菌多样性及群落组成的影响,以及AM真菌群落组成和多样性对土壤环境因子的变化产生的响应情况,为宁夏南部山区合理农作物免耕覆盖、有机栽培管理提供一定的理论依据。
1 材料与方法
1.1 研究区概况
试验在宁夏回族自治区固原市隆德县沙塘镇和平村(105°28′48″E,35°12′36″N)进行,该区域海拔1 883 m,年均降水量410 mm,年均蒸发量 1 370 mm,年均气温6℃,供试土壤为黑垆土。试验于2015年4月开始,前茬作物为蚕豆,品种为临蚕6号,冬麦品种为蓝天32号,均购自银川种子销售公司。供试土壤基本理化性状为全氮0.52 g·kg-1、全磷0.65 g·kg-1、有机质10.63 g·kg-1,碱解氮41.0 mg·kg-1、速效磷45.5 mg·kg-1、速效钾170.5 mg·kg-1、pH值9.0。
1.2 试验设计
分别于2015年、2016年秋季种植冬麦,冬麦前茬作物为蚕豆。采用双因素随机区组设计,设置2个因素:因素1为耕作方式,包括免耕覆盖(记作N)、传统耕作不覆盖(记作T) 2个水平;因素2为施肥方式,包括施有机肥(记作F)和不施有机肥(记作C)2个水平。共4个处理,即免耕覆盖+施有机肥(记作NF)、免耕覆盖+不施有机肥(记作NC)、传统耕作不覆盖+施有机肥(记作TF)和传统耕作不覆盖+不施有机肥(记作TC);每个处理4次重复。每个小区面积为12 m2,共16个小区。
2015年开始,免耕覆盖处理的小区均不进行土壤翻耕,传统耕作处理进行人工翻耕。2015年免耕覆盖处理冬麦播前将200 kg·667 m-2小麦脱粒后的颖壳均匀的覆盖于土壤表面,后续试验均不再覆盖。免耕覆盖处理在冬麦生育期内定期将地上杂草齐地表割掉,覆盖在冬麦行间;耕作处理定期将田间杂草连根拔掉。2015年9月中旬种植冬麦,于2016年7月中旬收获。2016年9月中旬种植冬麦,于2017年7月中旬收获。
2015年、2016年冬麦播种量均为25 kg·667 m-2,每小区种12行,每行播56.25 g。免耕覆盖处理,播前将有机肥均匀地撒在冬麦行间,耕作处理有机肥在播前随耕翻施入土壤。所施用的生物有机肥(黄腐酸≥12%、有机质≥45%、N+P2O5+K2O≥4%),每个小区施肥量折合纯氮(N%)施量为108 g、纯磷(P2O5%)施量为23.6 g、纯钾(K2O%)施量为22.3 g。
1.3 样品采集与分析
1.3.1 取样方法 2017年冬麦收获前利用多点取样的方法,采集各小区0~20 cm土层土壤,各小区土壤样本独立,土壤样本用冰盒保鲜带回。土壤样品立即过1 mm筛,一部分置于-80℃冰箱保存备用,用于土壤AM真菌多样性测定;其余样品自然风干,用于土壤理化性状分析。
1.3.2 土壤AM真菌DNA 提取、PCR 扩增及回收纯化 采用Fast DNA Spin Kit试剂盒(Illumina公司,美国)提取土壤AM真菌DNA,按照说明书步骤进行提取。引物信息如表1所示。扩增片段大小为280 bp。PCR扩增在ABI-2720扩增仪(美国ABI公司)上进行。扩增体系:5×反应缓冲液5 μL,5×GC缓冲液5 μL,2.5 mmol·L-1dNTP 2 μL,10 μmol·L-1前、后引物各1 μL,DNA模板40 ng,Q5 DNA聚合酶(NEB公司,美国)0.25 μL,ddH2O补足至25 μL,扩增程序:98℃预变性2 min;98℃变性15 s,55℃退火30 s,72℃延伸30 s,共30个循环;72℃终延伸5 min,10℃保存;扩增产物利用2%琼脂糖凝胶电泳检测,采用凝胶回收试剂盒(美国AXYGEN公司)对目标片段进行切胶回收。由上海派森诺生物科技有限公司Illumina MiSeq测序平台进行双端测序分析。

表1 本研究中所用引物Table 1 The primer sequences used in this study
1.3.3 土壤理化性状的分析 土壤全氮采用半微量凯式法测定;碱解氮采用碱解扩散法测定;速效磷采用NaHCO3浸提-钼锑抗比色法测定;速效钾采用NH4OAc浸提,火焰光度法测定;有机质采用重铬酸钾外加热法测定;pH值利用PHSJ-4F pH计(上海仪电科学仪器股份有限公司)测定(1∶5土水比)[21]。
1.4 数据处理
采用QIIME软件和UCLUST序列比对工具[22],对根据质量控制筛选出的优质序列按97%的序列相似度进行归并和划分为不同的操作分类单元(operational taxonomic unit,OTUs),并选取每个OTU中丰度最高的序列作为该OTU的代表序列,根据获得的OTU丰度矩阵,使用R软件计算各样本(组)共有OTU的数量,在OTU聚类基础上对样品进行Alpha多样性分析,计算样品丰富度指数、多样性指数,根据分类学分析结果绘制样品菌群分布图比较差异。
采用两因素方差分析检验免耕覆盖、有机肥措施对AM真菌多样性及群落组成间的显著性(SPSS 19.0);Canoco 5.0软件进行AM真菌多样性、丰富度及基于属水平上AM真菌群落组成与土壤环境要素间的多元分析。
2 结果与分析
2.1 AM真菌测序深度分析
由图1可知,样品的AM真菌物种累计曲线逐渐趋于平缓,说明样本量足够大,测序数量趋于饱和,取样样品数基本合理,测序文库能够较真实地反映土壤样品中AM真菌微生物群落的物种数,测序结果基本能充分发掘分析样品中的大部分AM真菌种类,但仍可能会有少量AM真菌种类未被发现。
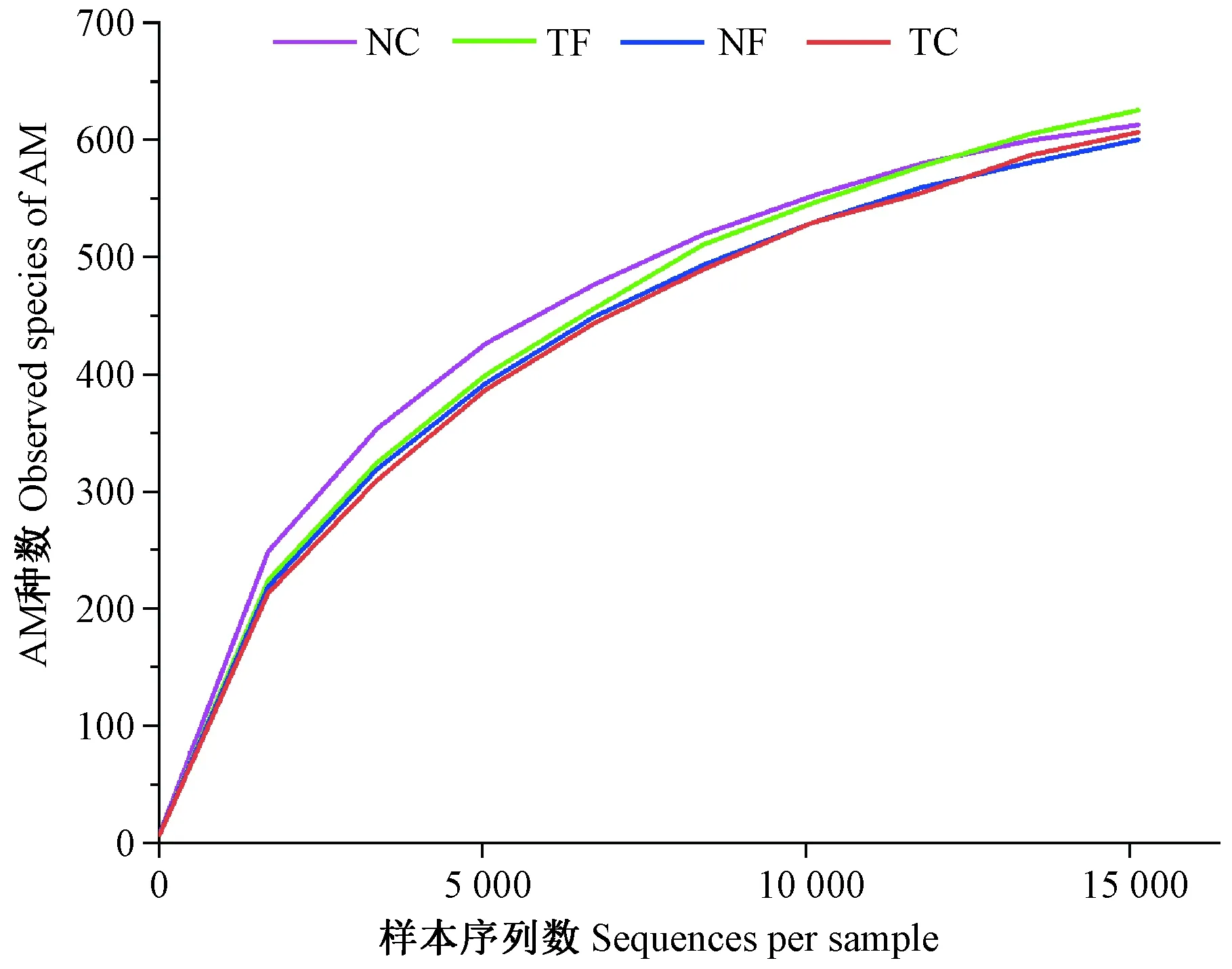
图1 AM真菌物种稀释曲线图Fig.1 Species rarefaction curve of AM fungi
2.2 免耕覆盖、有机肥施用对土壤AM真菌群落共有及特有OTUs的影响
将丰度值低于全体样本测序总量0.001%(1/100 000)的OTU去除,并将去除稀有OTU的丰度矩阵用于后续分析,在所有样本中共检测到OTU数为 4 515个。NC、NF、TC、TF处理分别获得1 187、1 106、1 075、1 147个OUT,各处理间共有OTU数为450个(图2)。NC、NF、TC、TF特有的OTU数分别为238、162、155、240个。结果表明,与免耕覆盖不施肥相比,有机肥施用后土壤AM真菌的总OTU数和特有OTU数分别降低了6.8%和31.93%。然而,在传统耕作条件下,有机肥的施用却提高了土壤AM真菌的总OTU数和特有OTU数,分别为6.27%和35.41%,表现出与免耕覆盖相反的变化趋势。有机肥的施用对各处理土壤特有AM真菌OTU数有明显的抑制或促进作用,其作用强度明显高于对土壤AM真菌总OTU数的影响。
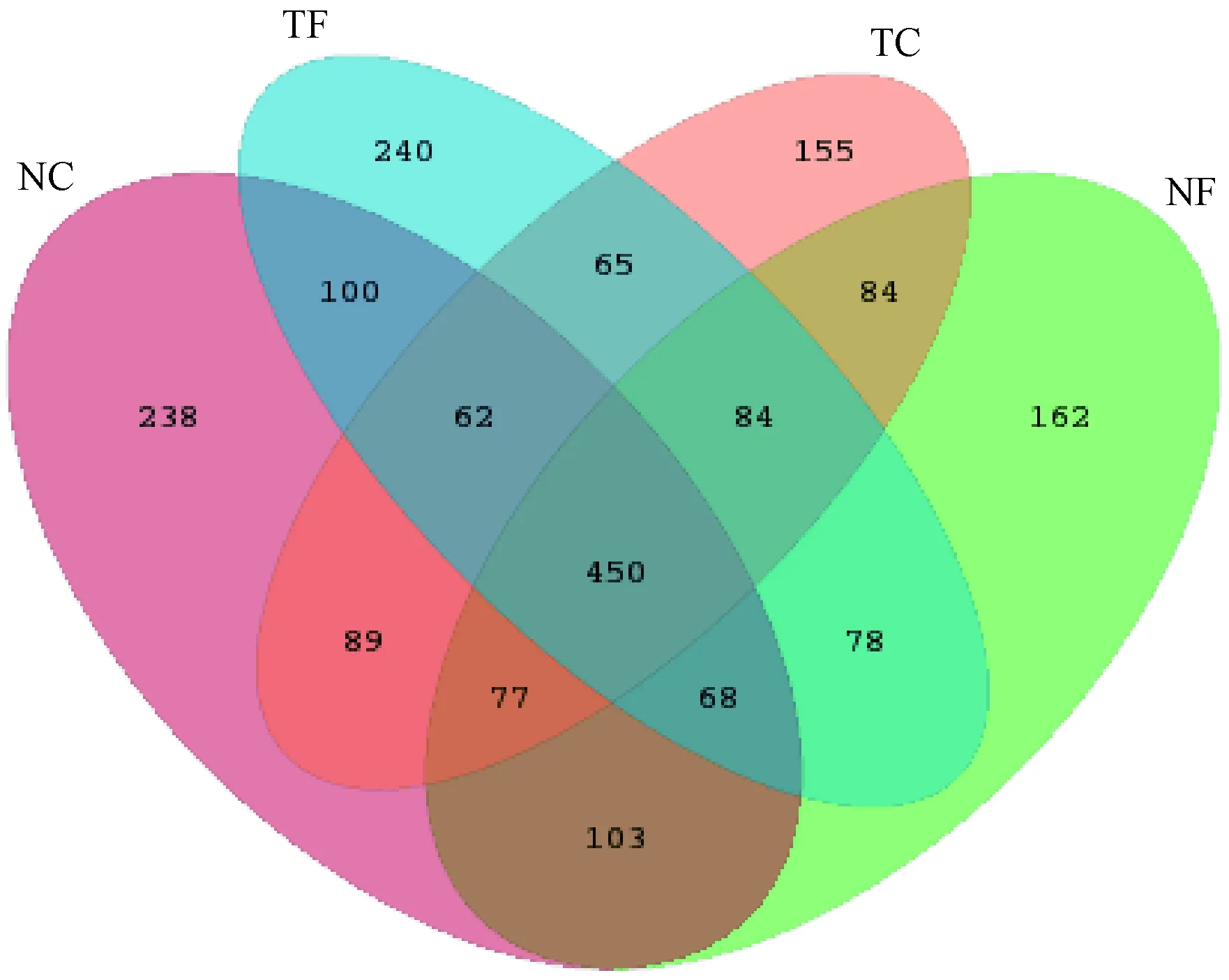
图2 不同处理下共有AM真菌OTUs的Venn图Fig.2 Venn diagrams of the number of unique and shared AM fungal OTUs under different treatment
2.3 免耕覆盖、有机栽培对土壤HM真菌多样性的影响
由表2可知,免耕覆盖(NC)及传统耕作(TC)处理下,有机肥的施用均降低了土壤AM真菌的多样性指数,其中Shannon指数降低了3.70%~8.40%,Simpson指数降低了2.11%~3.26%,而AM真菌丰富度中Chao1指数则增加了1.94%~2.63%、ACE指数增加了3.84%~5.62%。从施肥方式来看,不施肥及施有机肥下,传统耕作模式均降低了AM真菌的多样性指数,而提高了AM真菌的丰富度指数。与NC相比,TC的Shannon指数降低了8.40%,Simpson指数下降了3.16%,Chao1指数增加了5.01%、ACE指数增加了8.31%;TF与NF间也表现出类似的变化趋势,但各处理间均无显著差异。结合图2各处理共有或独有的OTUs可知,农业综合管理活动中耕作覆盖方式,有机肥的施用与否均会对土壤AM真菌的多样性及丰富由于C=O、C-H、N-H的伸缩振动引起的,其中C-H的伸缩振动也与胶稠度紧密相关[29]。度产生一定影响,其中多样性变化趋势可能与农业综合管理活动时间的长短以及群落组成中AM种属分布特征有关。
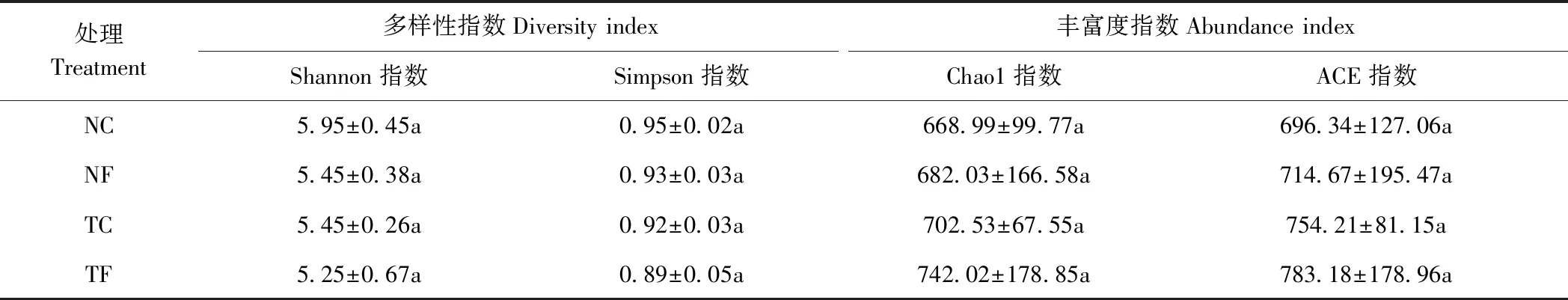
表2 免耕覆盖、有机栽培对土壤AM真菌多样性的影响Table 2 Effect of no-tillage, mulching and organic fertilization management to AM fungi diversity
注:同列不同小写字母表示各处理间差异显著(P<0.05)。
Note: Different lowercase letters in the same column indicate significant difference among treatment.
2.4 免耕覆盖、有机栽培影响下土壤AM真菌在属水平的分类特征
不同处理土壤中AM真菌共获得4 515个OTUs,分属于1门3纲4目8科10属155种;其中,类球囊霉属(Paraglomus)、球囊霉属(Glomus)、近明球囊霉属(Claroideoglomus),为各处理AM真菌群落的优势属,其AM真菌平均相对丰度占AM真菌OUT总丰度的94.5%,而原囊霉属(Archaeospora)、盾巨孢囊霉属(Scutellospora)、双型囊霉属(Ambispora)、多孢囊霉属(Diversispora)、有隔球囊霉属(Septoglomus)、巨孢囊霉属(Gigaspora)、无梗囊霉属(Acaulospora)仅占总丰度的5.5%。
由图3可知,免耕覆盖下,有机肥的施用(NF)提高了类球囊霉属(Paraglomus)、多孢囊霉属(Diversispora)AM真菌的相对丰度。其中多孢囊霉属的相对丰度提高了1.14倍,原囊霉属也提高了8.2%,其他AM真菌属水平上OTU相对丰度则分别下降了5.1%~44.4%。传统耕作不覆盖条件下,有机肥施用后(TF),类球囊霉属的相对丰度增加了36.4%,较免耕处理提高了4.45倍。优势属中,球囊霉属的相对丰度也提高了26.7%,但近明球囊霉属的相对丰度表现出与免耕处理相同的下降趋势;其他各属的丰度变化幅度较大,平均在20%~300%。总体来看,无论在免耕覆盖还是传统耕作条件下,有机肥的施用均会影响AM真菌属水平上的相对丰度。有机肥的施用提高了类球囊霉属的相对丰度,对近明球囊霉属有明显的抑制作用,降低了其丰度;丰度较低的其他7属(原囊霉属、盾巨孢囊霉属、双型囊霉属、多孢囊霉属、有隔球囊霉属、无梗囊霉属、巨孢囊霉属)AM真菌更易受到有机肥施用的明显影响。
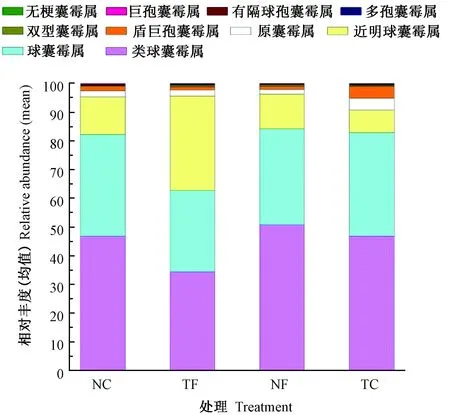
图3 免耕覆盖有机肥管理对基于属水平上AM真菌相对丰度组成的影响Fig.3 Effect of no-tillage, mulching and organic fertilization management to the relative abundances of AM fungi based on the genus level
不施肥条件下,与传统耕作(TC)相比,免耕覆盖(NC)的土壤AM真菌类球囊霉属、球囊霉属的相对丰度分别下降了26.55%、19.79%,但近明球囊霉的相对丰度却提高了1.53倍。随着有机肥的施用,传统耕作(TF)较免耕覆盖(NF)而言,类球囊霉属的相对丰度约降低了7.37%,近明球囊霉和球囊霉属的相对丰度分别增加和降低了7.05%、35.54%。综上,耕作管理活动明显抑制了AM真菌类球囊霉属的相对丰度;施肥或不施肥条件下,耕作与否对球囊霉属和近明球囊霉相对丰度的变化会产生相反的作用效果。
AM真菌属水平上相对丰度分布的方差显著性检验结果表明(图4),受耕作覆盖及有机肥施用方式影响,传统耕作不施肥(TC)条件下土壤AM真菌中近明球囊霉相对丰度与免耕施肥(NF)、免耕无肥(NC)、传统耕作施肥(TF)间均存在显著差异(P=0.02、0.04、0.006)。两因素方差分析结果表明,肥料因素(P=0.014)和两因素间(肥料因素×耕作覆盖)的交互(P=0.019)对不同处理下近明球囊霉相对丰度有显著影响。免耕覆盖、不施肥(NC)处理中,有隔球囊霉属的相对丰度与传统耕作施用有机肥(TF)间存在极显著差异(P=0.009);不同处理下肥料因素、耕作覆盖因子对土壤AM真菌有隔球囊霉属的相对丰度均有显著影响(P1=0.029,P2=0.029)。
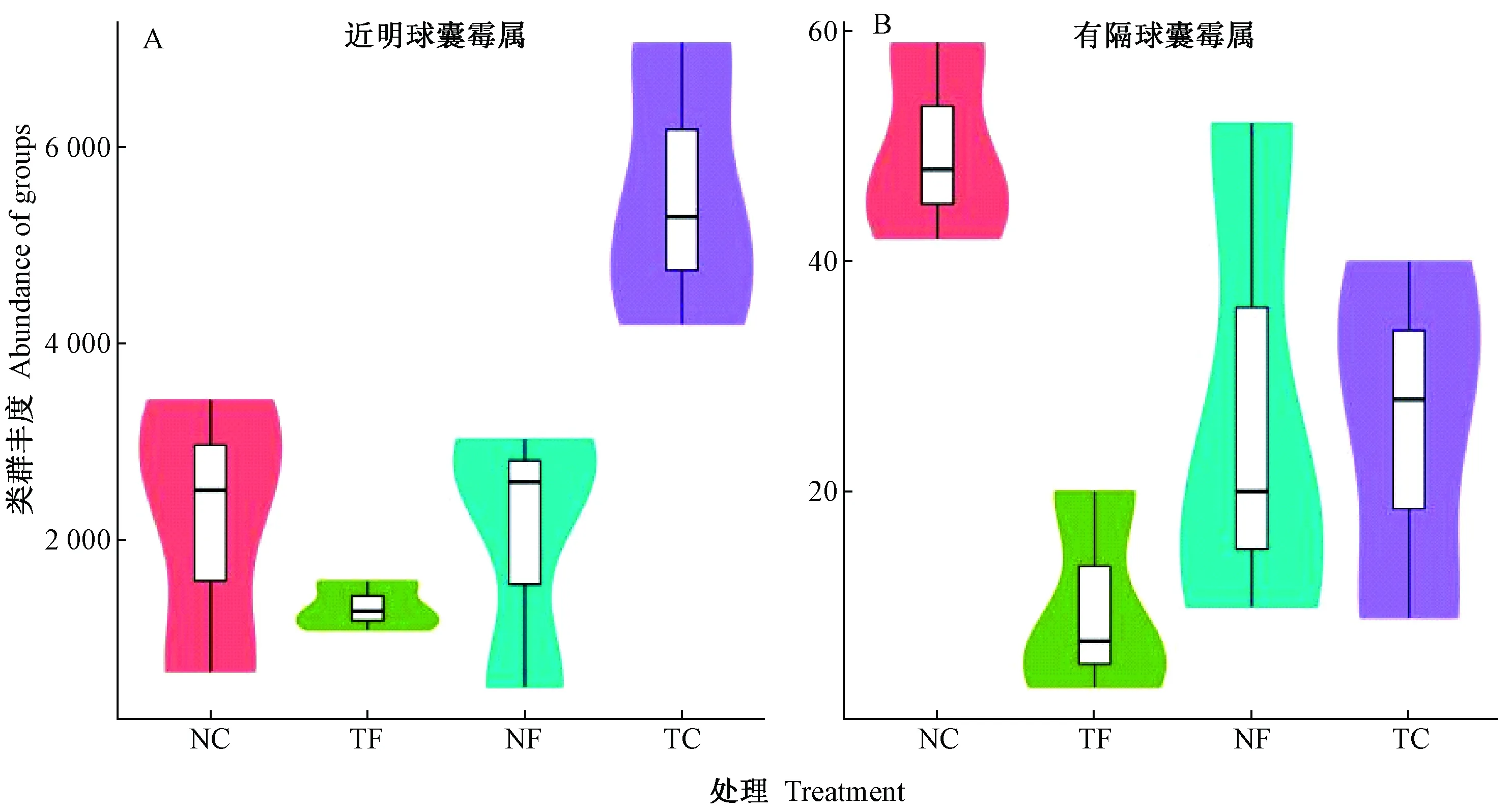
图4 免耕覆盖有机肥管理影响下的土壤AM真菌属水平类群丰度差异Fig.4 Difference in abundance of AM fungi in soil under no-tillage and organic management
2.5 免耕覆盖有机肥管理下AM真菌属水平的相对丰度、多样性与土壤环境因子的关系
由图5可知,第1、第2排序轴的累计贡献率均大于70%,说明土壤环境因子指标能在70%以上解释AM真菌多样性、属水平的相对丰度与土壤环境因子间的相关关系及差异性。
由图5-A可知,第1排序轴(43.46%)、第2排序轴(27.59%)能够在累计变量71.05%上揭示土壤中AM真菌多样性指数、丰富度指数的变化。不同处理下AM真菌多样性及丰富度的空间变异受全磷、速效磷、全氮浓度及土壤pH值的影响;而两类用以表征AM真菌多样性与丰富度的参数均与速效磷、全磷浓度呈正相关,与土壤pH值、全氮浓度呈负相关;其中,全磷浓度与AM真菌多样性(Shannon指数、Simpson指数)相关性最大,而与丰富度(ACE指数、Chao1指数)相关性最大的是土壤速效磷浓度,说明影响AM真菌多样性与丰富度的首要因子是土壤速效磷浓度。
由图5-B可知,第1、第2排序轴能够在累计变量72.89%上解释土壤中AM真菌属水平相对丰度的空间变化,全磷、速效磷、全氮含量、碱解氮、速效钾等均对AM真菌优势属的丰度有一定的影响。对优势属而言,类球囊霉属与全磷呈正相关,且相关性最大,而球囊霉属与速效磷呈显著正相关,相对丰度仅次于前两者的近明球囊霉,其相对丰度高低与土壤速效钾、碱解氮含量呈负相关。
3 讨论
农田生态系统中,土壤AM真菌群落结构及多样性变化受耕作措施及施肥制度等多种因素的影响[23-24]。本研究中,不同耕作与施肥措施下土壤AM真菌群落的组成及多样性变化存在一定差异。从基于OTU数的群落结构分析可知,4种处理下的土壤AM真菌类群分布有部分一致的类群;但从耕作覆盖及施肥因素来看,不同处理间AM真菌的存在较为明显的差异,免耕覆盖处理2组样本共有的OTU数高于其他样本两两共有,达到最大,施肥处理次之,说明免耕条件下,免耕施肥和免耕土壤AM真菌群落分布相似度高于耕作处理。这与Verena 等[25]的研究结果一致。表明免耕条件下的土壤环境更适宜于AM真菌群落生长和繁殖,与传统耕作相比,免耕能提高AM真菌物种丰度与多样性,形成独特的AM真菌群落结构。Irene等[26]用覆盖替代冬季休耕,在随后种植向日葵或玉米的土壤中发现AM真菌生物量及多样性均有所提高;也有研究通过将常规耕作与免耕系统进行比较,发现土壤频繁耕作会对AM真菌丰富度的产生不利影响,这是传统耕作稀释了AM真菌繁殖体造成的结果[25]。高萍等[9]对14年保护性耕作条件下的AM真菌多样性进行研究,也发现免耕土壤中的AM真菌多样性显著高于耕作处理。上述研究均说明实施保护性耕作措施的时间越长,越有利于AM真菌多样性的提高[27-28],土壤扰动会降低AM真菌的多样性[28-29]。这与本研究结果基本一致。研究发现在不考虑耕作管理对AM真菌影响的条件下,土壤施肥方法、制度与AM真菌的多样性和组成也存在较为密切的关系。Liu等[30]研究表明,施肥减少了AM真菌的多样性,使AM真菌群落向特定物种或区系的优势种转移。而关于施肥对AM真菌多样性的中性或者负作用影响也有报道,如刘佳[31]认为,施有机肥和化肥均能提高AM真菌多样性。这与本研究结果相反。这可能与本试验中施用有机肥导致土壤AM真菌类球囊霉属、球囊霉属、近明球囊霉等优势种发生变化,为其他属的AM真菌成功定殖提供了更多的机会有关。研究发现有机肥施用导致的土壤环境变化,主要表现为改善土壤结构,增强土壤保水性和透气性,促进寄主植物的生长发育[32],这也可能是导致AM真菌多样性降低,丰富度提高的重要因素之一,说明AM真菌群落对有机肥施用的响应受多种因素相互作用的影响。
本研究中,土壤AM真菌优势属有类球囊霉属与球囊霉属。这与李雪静等[33]对荒漠和草原生态系统的研究结果一致。前人对森林[34]、草原[35]、湿地[36]、农田生态系统[37]的AM真菌群落结构及多样性进行研究,发现AM真菌群落组成主要由球囊霉属决定。这与本研究结果基本相似。球囊霉属具有独特的繁殖方式,能够耐受各种环境的变化,因此分布广泛[38]。与传统耕作相比,在受干扰少的土壤(免耕)中,类球囊霉属与球囊霉属相对丰度的分布更集中、更均匀。耕作和施肥管理改变了AM真菌属水平的相对丰度,免耕覆盖条件下施肥提高了类球囊霉属相对丰度;但耕作条件下有机肥施用后则表现出相反的变化趋势,其中近明球囊霉属的相对丰度变化与类球囊霉属相对丰度变化规律相似。无论耕作与否,施肥均降低了球囊霉属相对丰度,而免耕条件下施肥增加了其相对丰度。属水平的显著性差异分析结果表明,施肥和耕作、覆盖措施都对土壤中有隔球囊霉属造成了较大程度的破坏和抑制,降低了其丰度,近明球囊霉属相对丰度对有机肥施用也表现出相似的响应。
研究表明,环境因子(气候、土壤理化因子)对AM真菌群落结构的影响较大[39-40]。本研究中,土壤速效磷、全磷与AM真菌多样性和群落结构存在显著相关关系,这与Smilauer[41]和Mathimaran等[42]研究认为AM真菌多样性、群落结构组成与土壤磷元素浓度并无显著相关关系的结论不一致。但也有研究结果表明,土壤磷与AM真菌群落结构或多样性显著相关[43],这与本研究结果相似。分析认为,在一定的土壤环境条件下,AM真菌丰富度随着磷含量的增加而呈上升的趋势,当浓度升高到一定程度时,AM真菌丰富度与多样性呈下降的趋势,其主要原因是土壤的高磷浓度使宿主植物根系分泌物发生了改变,从而导致AM真菌生存环境发生变化。此外,土壤pH值与有机质也是影响AM多样性的重要因素。我国东南沿海地区AM真菌群落结构的研究表明,球囊霉属对土壤pH值的适应范围较宽,在pH值为5~9的土壤中均广泛分布[25]。本试验结果表明,土壤AM真菌多样性和群落结构与土壤pH值呈显著负相关,而有机质、速效磷与AM真菌多样性呈显著正相关。研究发现由于有机质、速效磷等土壤养分与生态系统中物质的循环和土壤中各类生物的活性有关[44],有机质能够通过影响地上生物群落组成和多样性而间接影响与之共生的AM真菌多样性与群落组成,并在一定范围内对AM真菌群落组成起到显著的促进作用。因此,随着土壤中有机质的增加,其AM真菌多样性也随之升高,从而通过相互促进来调节作物生长过程中养分的吸收及利用。
4 结论
免耕覆盖、有机肥综合管理下,土壤AM真菌优势属主要为类球囊霉属、球囊霉属、近明球囊霉属;免耕措施增加了类球囊霉属相对丰度,施肥活动增加了近明球囊霉相对丰度。冬麦免耕不施肥栽培管理提高了AM真菌多样性,而耕作施肥管理降低了AM真菌的丰富度。近明球囊霉相对丰度受农业综合管理措施中有机肥施用以及免耕覆盖、有机肥施用交互作用的影响显著。土壤磷含量是影响AM真菌群落结构与多样性的重要因素,AM真菌群落对耕作、覆盖、有机肥施用的响应受多因素相互作用的影响。
猜你喜欢
现代园艺(2017年21期)2018-01-03 06:41:42
水土保持研究(2016年4期)2016-10-21 02:28:59
绿色科技(2016年16期)2016-10-11 06:53:51
新农业(2016年15期)2016-08-16 03:40:48
土壤与作物(2015年3期)2015-12-08 00:46:58
亚热带资源与环境学报(2015年1期)2015-01-22 07:04:58
水土保持研究(2014年4期)2014-09-21 02:24:12
作物研究(2014年6期)2014-03-01 03:39:12
新疆农垦科技(2014年1期)2014-02-28 19:18:59
