
大黄素抑制胰腺癌裸鼠原位移植瘤新生血管形成的机制研究
2019-05-17 07:44:50陈敏远徐锦波王兆洪胡万乐
浙江医学 2019年8期
陈敏远 徐锦波 王兆洪 胡万乐
近30年大量研究证实,血管生成是许多肿瘤包括胰腺癌发生的早期事件,也是肿瘤侵袭和转移的必要条件。微血管密度(MVD)作为检测肿瘤新生血管形成的金标准由来已久,其是由CD31或CD34等内皮细胞标记物染色后显微镜下观察、计算所得,与肿瘤的侵袭性密切相关[1]。血管内皮生长因子(VEGF)作为血管新生重要的促血管生成因子,与肿瘤新生血管形成密切相关[1]。血管生成素(angiopoietin,Ang)系统及其相关的因子是继VEGF之后,人们发现的又一重要的内皮特异性的促血管生成因子。与VEGF不同,Ang系统作用于血管生成的晚期阶段,诱导和维持内皮细胞迁移存活,在机体和肿瘤血管生成中发挥着重要的作用。Ang家族由Ang-1、Ang-2、Ang-3 和 Ang-4 组成,其中 Ang-1、Ang-2与它们共同的受体酪氨酸激酶受体2(Tie-2)与血管生成密切相关[2]。胰腺癌是典型的富血管性的肿瘤,其生长和转移都依赖血管生成调节因子的调控。血管生成不仅为肿瘤细胞的生长提供了营养,更为肿瘤的浸润和转移提供了通路。本研究旨在探讨大黄素对胰腺癌裸鼠原位移植瘤新生血管形成相关因子VEGF、Ang-1、Ang-2及Tie-2表达的影响及其作用机制。
1 材料和方法
1.1 主要药物和试剂 大黄素(纯度>98%,美国Sigma公司),用二甲基亚砜(DMSO)溶解(DMSO的终浓度小于 0.1%);FBS、含乙二胺四乙酸(EDTA)的胰酶、RPMI-1640培养基(美国Gibco公司);Trizol(美国Invitrogen公司);RevertAidTMFirst Strand cDNA Synthesis Kit(加拿大Fermentas公司);Ang-1、Ang-2和VEGF抗体(美国Santa Cruz公司);Tie-2抗体(美国 Abcam公司);RT-qPCR反应所需引物由上海基康生物有限公司合成。
1.2 细胞株和动物 人胰腺癌细胞株SW1990购自美国组织培养库。雌性BALB/c-nu/nu品系的裸小鼠40只,4~6周龄,体重18~20g,均购自中国科学院上海动物实验中心。饲养于SPF级屏障系统的洁净层流架内,室温控制在(25±1)℃,相对湿度40%~60%。
1.3 细胞培养 人胰腺癌细胞株SW1990培养于含10%FBS的RPMI 1640培养基中,置于含5%CO2、37℃细胞培养箱中。待细胞单层贴壁稳定生长并铺满整个培养瓶时用胰蛋白酶消化传代、分瓶培养,待建立动物模型。
1.4 动物模型制备
1.4.1 皮下移植瘤制备 取对数生长期的SW1990细胞(细胞数为2×106)进行裸鼠皮下注射,待生长成1cm3皮下移植瘤后取出,无菌条件下去除中央坏死组织,选取周围健康肿瘤组织剪成1mm3的组织块待用。
1.4.2 胰腺癌裸鼠原位移植瘤模型制备 戊巴比妥钠(50mg/kg)腹腔麻醉裸鼠,左上腹直肌旁切口,暴露脾脏和胰尾,剪开胰腺被膜,将瘤块植入胰尾,缝合胰尾被膜。共制备40只裸鼠原位移植瘤模型。
1.5 分组与给药方法 手术后第3周开始给药,将实验动物采用随机数字表法分为N组(腹腔注射,DMSO终浓度<0.1%的氯化钠注射液)、E20组(腹腔注射,20mg/kg大黄素)、E40组(腹腔注射,40mg/kg大黄素)、E80组(腹腔注射,80mg/kg大黄素)4组,每组10只,每周均治疗3次,共治疗2周[3]。
1.6 胰腺肿瘤重量测定及抑瘤率计算 末次用药后1周颈椎脱臼法处死4组裸鼠,留取原位移植瘤并称重,计算抑瘤率,抑瘤率=(1-给药组平均瘤质量/对照组平均瘤质量)×100%。
1.7 原位移植瘤微血管密度(MVD)测定 采用免疫组化法。取固定好的肿瘤组织,经脱蜡、水化、3%H2O2封闭以及高压热抗原修复后,加入兔血清封闭,一抗体4℃孵育过夜,PBS洗涤后滴加辣根过氧化物酶(HRP)标记的二抗室温孵育30min,DAB显色3~5 min,苏木素复染后树脂封片镜检。低倍镜(×100)下寻取血管热点区域,然后高倍镜(×400)下随机选择5个视野,计数每个视野中CD31阳性细胞数,其平均值即为MVD[4]。
1.8 原位移植瘤Ang-1、Ang-2、Tie-2及VEGF mRNA表达水平测定 采用RT-qPCR法。按照Trizol试剂盒使用说明书从肿瘤组织提取总RNA,并计算RNA纯度和浓度。选取OD260/OD280比值在1.8~2.0的RNA进行后续试验。根据说明书配置好反应液,加入随机引物进行逆转录,采用罗氏染料进行荧光定量,β-actin为内参。由LightCycler Real Time PCR扩增仪获取实验数据。RT-qPCR引物序列见表1。

表1 RT-qPCR引物序列
1.9 原位移植瘤中VEGF、Ang-1、Ang-2及Tie-2蛋白表达水平测定 采用Western blot法。取新鲜肿瘤组织,RIPA裂解液裂解组织,提取上清液,采用BCA方法测量定量蛋白浓度后取等量蛋白质样品(50μg/孔),8%SDS-聚丙烯酰氨凝胶电泳电泳,湿转膜法转膜,用含5%无脂牛奶TBST封闭,滴加一抗4℃孵育过夜,HRP标记的二抗孵育2h,用ECL发光液显色。甘油醛-3-磷酸脱氢酶(GAPDH)为内参。采用Totallab 2.1软件分析Western印迹图像。以目的蛋白条带与内参条带吸光度值的比值反映VEGF、Ang-1、Ang-2及Tie-2蛋白的相对表达量。
1.10 统计学处理 采用SPSS 13.0统计软件。计量资料以表示,多组间比较采用单因素方差分析,两两比较采用LSD-t检验。P<0.05为差异有统计学意义。
2 结果
2.1 4组裸鼠原位移植瘤重量和抑瘤率的比较 E40组、E80组与对照组相比,差异均有统计学意义(均P<0.05),见表 2。
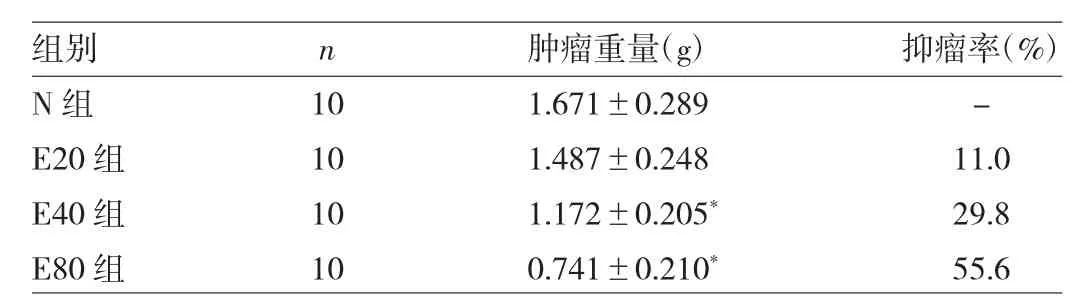
表2 4组裸鼠原位移植瘤重量和抑瘤率的比较
2.2 4组裸鼠原位移植瘤组织MVD的比较 4组裸鼠的MVD表达见图1。其中N组MVD为19.3±1.9,E20组 MVD 为 11.7±1.6,E40 组 MVD 为 7.9±2.3,E80 组MVD为7.2±1.8。各试验组MVD与对照组比较,差异均有统计学意义(均P<0.01)。
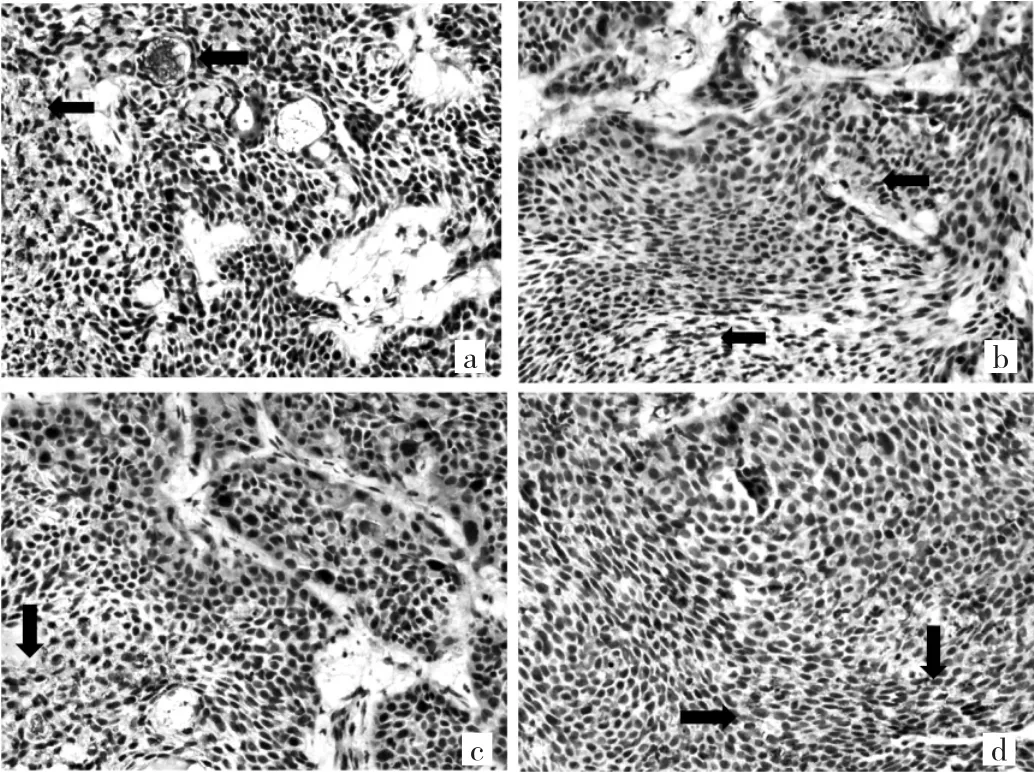
图1 4组裸鼠原位移植瘤组织MVD的表达(a:N组;b:E20组;c:E40 组;d:E80 组;箭头提示血管热点区域;DBA 显色,×400)
2.3 4组裸鼠原位移植瘤组织中VEGF、Ang-1、Ang-2及Tie-2 mRNA表达水平的比较 E20组、E40组、E80组的VEGF、Ang-1、Ang-2及Tie-2 mRNA表达与N组相比,差异均有统计学意义(均P<0.05),见图2。
2.4 4组裸鼠原位移植瘤组织中VEGF、Ang-1、Ang-2及Tie-2蛋白表达水平的比较 E20组、E40组及E80组的VEGF、Ang-1、Ang-2及Tie-2蛋白与N组比较差异均有统计学意义(均P<0.05),见图3。

图2 4组裸鼠原位移植瘤组织中VEGF、Ang-1、Ang-2及Tie-2 mRNA表达水平的比较(与对照组比较,*P<0.05)

图3 4组裸鼠原位移植瘤组织中VEGF、Ang-1、Ang-2及Tie-2蛋白表达水平的比较(a:4组 VEGF、Ang-1、Ang-2及 Tie-2蛋白表达水平的比较;b:4组 VEGF、Ang-1、Ang-2及 Tie-2蛋白表达的电泳图;与对照组比较,*P<0.05)
3 讨论
大量实验已经证实,肿瘤的生长依赖于血管新生[4]。随着肿瘤MVD的增加,其侵袭周围组织以及转移至其他脏器的能力明显增强。MVD被认为是预测肿瘤转移、复发和预后的一项重要指标[5]。本研究通过对移植瘤组织中CD31的检测,从而确定其MVD。在本研究中,大黄素各剂量组的MVD均<N组,差异均有统计学意义。这一结果与之前的文献报道相一致[3],因此笔者推断,大黄素可通过抑制肿瘤血管新生而抑制肿瘤的生长、发展。
VEGF作为血管新生重要的促血管生成因子,与肿瘤新生血管形成密切相关。本研究中课题组通过对胰腺癌裸鼠原位移植瘤组织中VEGF蛋白和mRNA的分析得到,VEGF在N组中蛋白以及mRNA均高表达,而在大黄素各剂量组中均低表达,且与大黄素浓度呈负相关。大黄素可能通过抑制VEGF的表达,使VEGF与其受体的结合减少[1],从而降低其促血管生成的作用。
在血管形成的早期,VEGF可促进内皮细胞增殖和迁移,形成初级血管网,但是VEGF单纯过表达诱导的新生血管是不成熟的,血管的成熟有赖于Ang/Tie通路的作用。生理状态下,Ang-1和VEGF的作用相辅相成,在产生新生血管的过程中发挥着各自的功能。Ang-1与血管内皮细胞表面Tie-2受体结合,促进其磷酸化启动这一信号通路,进而使血管周围细胞围绕内皮细胞,形成支架结构,使血管壁连续而完整,完成血管再构这一过程。所以Ang-1能维持血管的完整,对血管形成必不可少[6]。Ang-1在乳腺癌、前列腺癌、膀胱癌、肝癌、胆管细胞癌以及胰腺癌等多种肿瘤中扮演着不同的角色[7-9]。Ang-2是Ang-1的天然拮抗剂,通过与Ang-1竞争同一受体阻断Ang-1的作用[6]。Ang-2与Tie-2受体结合后,可阻断其磷酸化。Ang-2可抑制血管周围细胞的聚集,使其不再围绕内皮细胞形成支撑,也能促进内皮细胞的凋亡,减少周细胞覆盖,导致血管处于一种不稳定状态,从而促进血管新生的发生。Ang-2的促血管生成作用与局部微环境以及VEGF等生长因子密切相关[6,10]。当存在VEGF时,其可与VEGF协同刺激血管的出芽生长,形成新生血管[11];而在没有VEGF等生长因子存在的条件下,其可阻断Ang-1稳定血管的作用,导致内皮细胞凋亡,显微镜下可见血管退化,微血管密度降低[8]。
本研究发现,大黄素可抑制Ang-1的表达,且大黄素各剂量组与N组相比差异有统计学意义。合理的解释是,Ang-1表达减少使得Ang-1与VEGF协同形成新生血管的能力降低,减少了肿瘤组织微血管的形成。本研究还发现,大黄素可抑制Ang-2、Tie-2的表达,且大黄素各剂量组与对照组比较差异均有统计学意义。这表明大黄素可能一方面通过对VEGF表达的影响,从而影响Ang-2、Tie-2的表达以及作用,另一方直接作用于Ang-2、Tie-2,从而抑制其表达,达到抑制胰腺癌血管新生的作用。
本研究表明大黄素可抑制胰腺癌裸鼠原位移植瘤的血管新生,大黄素对血管新抑制方式可能是以通过抑制 VEGF、Ang-1、Ang-2、Tie-2 为主。该研究结果将为大黄素用于临床胰腺癌治疗提供一定的实验依据,但还需进一步的动物实验和临床试验加以验证。
猜你喜欢
幼儿100(2024年19期)2024-05-29 07:43:34
保健医苑(2022年6期)2022-07-08 01:25:22
海峡姐妹(2019年8期)2019-09-03 01:01:02
天然产物研究与开发(2019年1期)2019-03-01 05:41:22
天然产物研究与开发(2018年5期)2018-06-13 03:23:54
天然产物研究与开发(2018年1期)2018-02-02 07:21:19
中成药(2017年12期)2018-01-19 02:06:32
天津医药(2016年9期)2016-10-20 03:19:39
上海工运(2015年11期)2015-08-21 07:27:00
中国中医药现代远程教育(2014年22期)2014-03-01 04:33:08
