
雷州半岛尾叶桉和湿加松人工林的蒸腾耗水规律
2019-05-13 02:06王志超许宇星竹万宽杜阿朋
生态学报 2019年6期
王志超,许宇星,竹万宽,杜阿朋
国家林业和草原局桉树研究开发中心,广东湛江桉树林生态系统国家定位观测研究站,湛江 524022
蒸腾是植物生命活动必需的生理代谢过程[1],其耗水占根部吸收水分的99%以上[2],树干液流是蒸腾作用在植物体内产生的上升流,提供99.8%以上的蒸腾耗水,因此可以通过精确测定树干液流来反映植株的蒸腾耗水状况[3]。树干液流受植物生物学结构、气象因子及土壤水分等因素的综合影响[4],其中生物学结构决定树干液流的潜在能力[5],而气象因子和土壤水分决定树干液流的变化趋势和瞬时波动特征[6- 8],因此准确分析掌握单株树干液流变化规律及其影响因素,并结合林分特征进行尺度转换,对于正确评价大面积人工林蒸腾耗水量及其对局域水资源的影响具有重要作用。
树木的蒸腾耗水量是人工林树种选择的重要参数[9],桉树和松树作为公认的速生林种,已成为雷州半岛地区重要的造林树种。湿加松(Pinuselliottii×P.caribaea)是湿地松(Pinuselliottii)与加勒比松(P.caribaea)的杂交子代,具有生长快、生长量大、树干圆满通直、抗逆性强等优点[10],20世纪50年代已在澳大利亚地区广泛种植,并取得成功[11- 12]。80年代开始在广东开展杂交实验,并随后在广东,广西,福建等亚热带南缘省份大面积推广。目前对于湿加松的研究主要集中在生长表现[13]、木材材性[14]、以及育种方面[15],而对于其蒸腾耗水特征方面的研究尚未见报道。桉树由于其本身的生物学结构及速生性决定了其单位时间单位面积的耗水量高于一般树种,再加上该地区降雨时空分配不均引起的季节性干旱,导致桉树耗水问题成为广泛关注和争论的焦点[16- 17]。目前已有很多学者进行了相关方面的研究,如张宁南等[18]研究了4年生尾叶桉(Eucalyptusurophylla)的液流密度和蒸腾耗水特征;孙振伟等[19]对柠檬桉(Corymbiacitriodora)树干液流及蒸腾耗水季节变异进行了研究,王志超等[20]对2年生—3年生尾巨桉(E.urophylla×E.grandis)旱雨两季树干液流特征进行了分析,Vertessy等[21]与Wullscheger等[22]研究了王桉(Eucalyptusregnans)的树干液流动态。但国内对于10年生以后大径材桉树蒸腾耗水特征的研究较少,更未有人对大径材阶段的尾叶桉和湿加松的蒸腾耗水规律进行对比分析。
本研究利用精确度高、对植株损伤小、可在树木自然生长状态下对树干液流的连续观测的Granier热扩散探针法(TDP)[23]对雷州半岛地区10年生尾叶桉树干液流进行持续观测,并与同期10年生湿加松树干液流对比分析,并结合自动气象观测站对环境因子作同步测定,以期揭示雷州半岛地区尾叶桉和湿加松人工林蒸腾耗水规律,为正确认识桉树的耗水问题及地区人工林树种选择提供数据支持和理论指导。
1 研究区概况
研究区位于广东雷州半岛地区,试验样地设置在广东湛江桉树森林生态系统国家定位观测站内(21°30′N,111°38′E),海拔90 m,地势平坦,属海洋性季风气候,年均降雨量约1500 mm,多集中在5—10月份,为雨季,占全年降雨量的77%—85%;年均温度23℃,年总辐射量4240 MJ/m2,年均相对湿度80.4%。试验区土壤类型主要是玄武岩砖红壤,土层厚度2 m以上,0—80 cm土层内平均有机质含量1.6%以上,pH 4.5—5.3,土壤肥力属中等水平。乔木层主要有尾叶桉、湿加松和尾巨桉;林下植被丰富,主要灌木有五色梅(Lantanacamara)、野牡丹(Melastomacandidum)、槲栎(Quercusaliena)等;主要草本植物有飞扬草(Euphorbiahirta)、飞机草(Eupatoriumodoratum)、蟛蜞菊(Wedeliachinensis)以及白花鬼针草(HerbaBidentisPilosa)等。
2 研究对象及方法
2.1 研究对象
研究对象为尾叶桉人工林和湿加松人工林,林龄均为10年,立地条件一致;其中尾叶桉人工林现存密度1245株/hm2,叶面积指数1.37,平均胸径为22.5 cm,平均树高21.7 m,平均冠幅3.61 m;湿加松人工林现存密度1375株/hm2,叶面积指数0.95,平均胸径为18.9 cm,平均树高12.3 m,平均冠幅2.45 m。在对尾叶桉和湿加松人工林林分调查的基础上,分别在各林分内设置20 m×20 m标准样地1个,作为观测试验区。
2.2 研究方法
2.2.1树干液流的测定
在尾叶桉和湿加松人工林标准样地内,分别选择5株生长良好、无病虫害、树干圆满通直无挤压的标准木作为液流持续观测样木,观测周期为1年(4月到次年3月),样木的各项参数详见表1。
树干液流测定方法采用Granier热扩散探针法,传感器采用德国Ecomatik公司SF-L4针型热扩散探针(33 mm),数据采集器采用DL2e数据采集器,采集时间间隔10 min,与气象监测时间同步。为避免不同高度、不同方位间液流的差异以及太阳辐射的影响,探针均安装在同一高度的北侧,同时用防辐射铝箔覆盖。
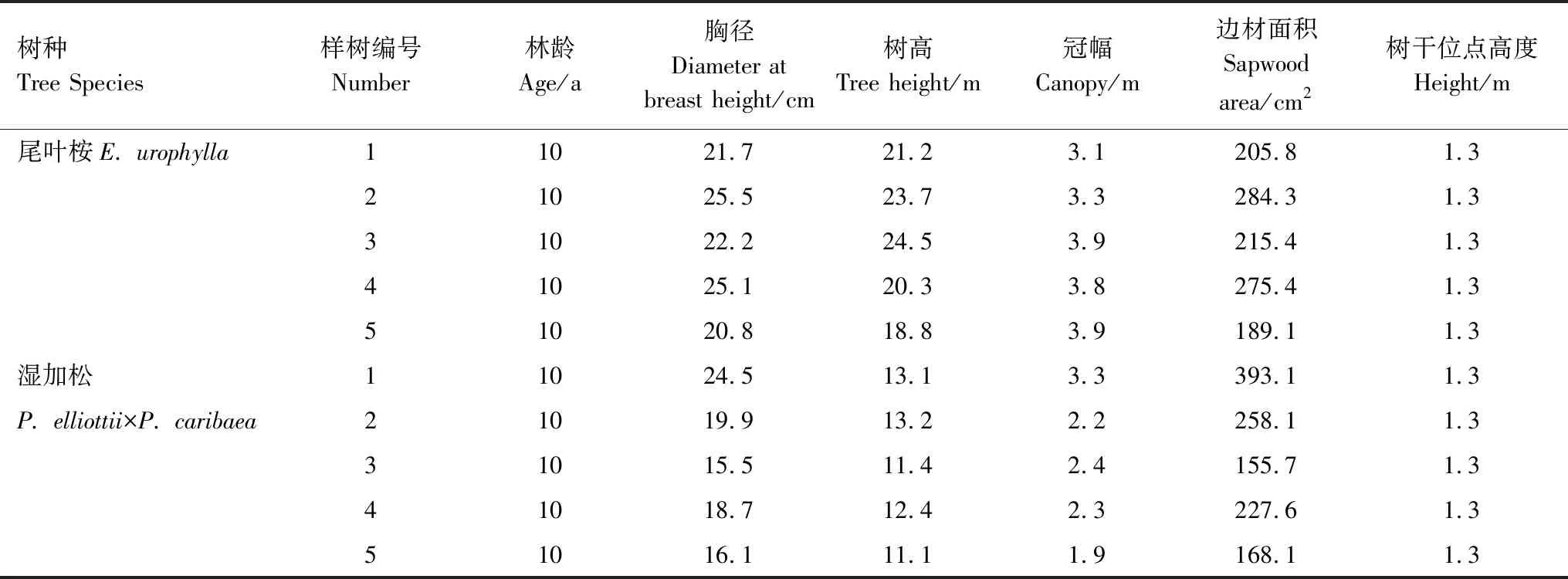
表1 被测样木基本参数
根据Granier[24]液流计算公式:
Js=0.0119×(dtmax/dtact-1)1.231×60
(1)
dt=T1-0-(T1-2+T1-3)/2
(2)
式中,Js为液流速率,cm/min;dtmax、dtact均由式(2)计算得到,dtmax一般指夜间空气湿度为100%长达2 d或树干直径停止变化、处于相对稳定状态时算得的dt值;T1-0、T1-2、T1-3分别为探针S0、S2、S3分别与S1间的温度差,℃。
2.2.2边材面积的测定
样木的边材面积无法直接测量,只能通过生长锥测出边材厚度,进而根据取芯处的树干直径以及树皮厚度进行计算。为避免生长锥对样木造成损伤进而影响液流速率的监测,在2处试验地内分别随机选取30株树,测量其胸径和边材厚度,每株树分东西、南北两个方向分别取木芯一条,测其边材厚度取平均值,计算边材面积,并建立胸径与边材面积的关系方程。经拟合回归我们得出:
尾叶桉人工林关系方程为:
As= 0.4382D2-2.701D+18.48
(3)
湿加松人工林关系方程为:
As′= 0.608D2.0232
(4)
两树种边材拟合方程决定系数R2均达到0.8以上,拟合效果较好。式中As、As′为边材面积,cm2,D为胸径,cm。根据关系方程计算出样树的边材面积(表1),进而计算液流通量。
2.2.3气象因子的测定
两处试验地相距较近,因此在尾叶桉和湿加松试验区之间开阔地带安置美国Campbell公司的CR3000型自动气象观测系统,连续观测试验期间的大气温度(T,℃)、空气相对湿度(RH,%)、太阳辐射(Slr,W/m2)、降雨量(Rain,mm)等气象指标,数据采集时间间隔为10 min,与液流监测时间间隔一致。同时为考虑大气温度和湿度的协同效应,利用大气温度和相对湿度计算空气饱和水汽压差(VPD,Kpa),公式如下:
(5)
式中,RH为空气相对湿度,%;T为空气温度,℃。
2.3 统计分析
采用Excel 2003、SPSS 19.0以及SigmaPlot 12.5统计软件对所有数据进行分析并作图。
3 结果与分析
3.1 尾叶桉、湿加松边材液流速率特征研究
3.1.1两种速丰林液流速率日变化特征
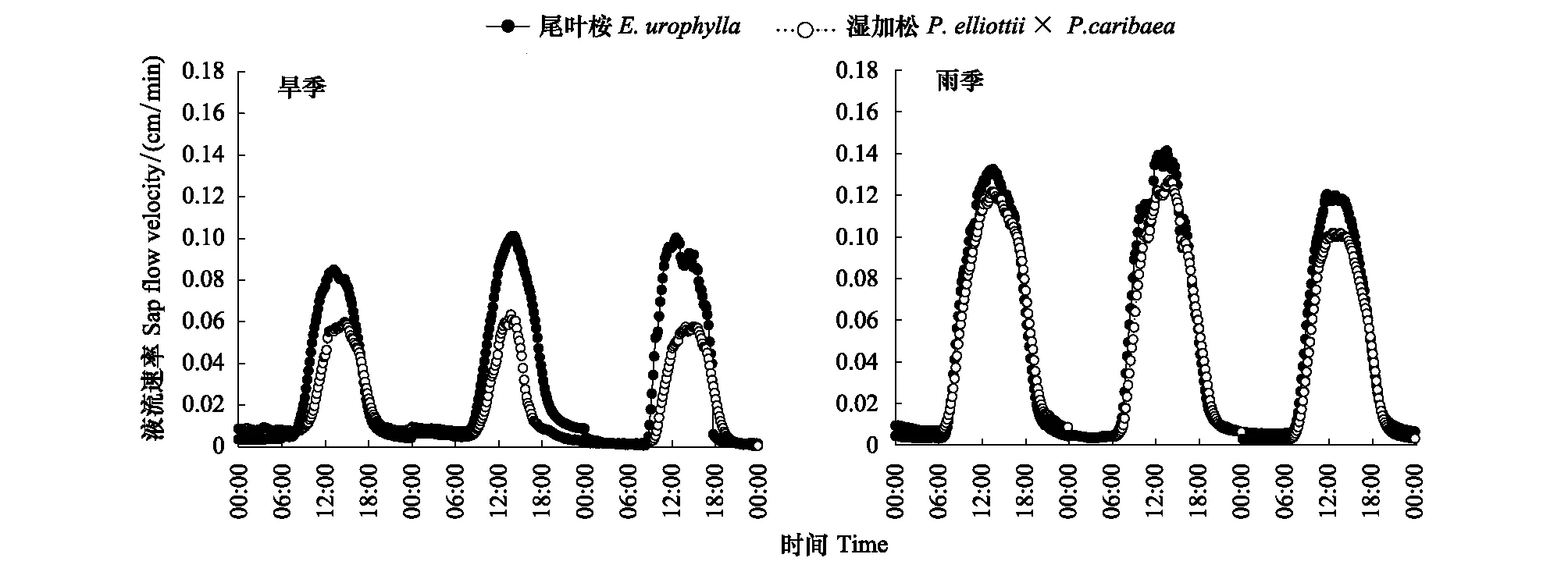
图1 尾叶桉、湿加松旱雨两季液流速率日变化特征Fig.1 Features and differences of sap flow in daytime during both rainy and dry seasons of E.urophylla and P.elliottii×P.caribaea
分别选取雨季(降雨量为1500.2 mm)和旱季(降雨量为204.5 mm)中典型月份(雨季为7月—9月;旱季为12月—次年2月)的日变化液流数据进行每10 min平均,得出两种速丰林旱雨季的平均日变化进程,如图1所示。由图可知,两种速丰林树干液流均表现出典型的昼夜变化规律,白天变化幅度大,呈典型的单峰型曲线。尾叶桉、湿加松雨季与旱季液流差异均较大,主要表现在日平均液流速率、日峰值,启动时间及液流迅速下降时间上,其中雨季日平均液流速率尾叶桉(0.045 cm/min)及湿加松(0.036 cm/min)分别是旱季的1.5倍和1.8倍,两个树种间日平均液流速率雨季尾叶桉是湿加松的1.27倍,旱季为1.54倍,存在较大差异;日峰值雨季尾叶桉(0.127 cm/min)是旱季的1.32倍,湿加松雨季(0.097 cm/min)是旱季(0.061 cm/min)的1.6倍,而尾叶桉雨季是湿加松的1.30倍,旱季为1.57倍;从启动时间来看,尾叶桉雨季启动时间在6:00—7:30之间,比旱季提前约1 h左右,湿加松雨季启动时间与尾叶桉差异不大,而旱季启动时间比尾叶桉旱季启动时间滞后约1—1.5 h;液流迅速下降时间雨季两树种间差异不大,均在16:30—17:30之间,而旱季尾叶桉较雨季要提前1—1.5 h,湿加松较尾叶桉旱季液流迅速下降时间仍提前约0.5—1 h左右。因此从图上看雨季两者差异不大,仅峰值尾叶桉略高于湿加松,而旱季尾叶桉峰型要高宽于湿加松。
分析夜间液流发现,两树种旱雨季均存在较大的夜间液流,且前半夜液流速率及变化幅度均较后半夜大,后半夜夜间液流均较小且平稳,推测原因可能是后半夜树干水分接近饱和,夜晚水分补充的时间主要为前半夜。尾叶桉雨季夜间平均液流速率(0.0059 cm/min)与旱季(0.0062 cm/min)无显著性差异;湿加松雨季夜间平均液流速率(0.0063 cm/min)与旱季(0.0060 cm/min)差异不显著,且两树种间夜间液流无论是旱季还是雨季差异均未达到显著水平。
3.1.2尾叶桉、湿加松平均液流速率月动态规律
树木边材液流速率受气象因素、土壤状况及自身生理特性的影响,呈现出明显的季节变化规律[25]。将尾叶桉、湿加松各月份液流速率进行平均,得出一年内各月份平均液流速率变化图(图2),由图中可以看出,研究期间尾叶桉月平均液流速率12月份最小,而湿加松1月最小,分别为0.027 cm/min和0.016 cm/min,最大值月份尾叶桉为8月,为0.051 cm/min,是最小月份的1.9倍;湿加松最大月平均液流速率出现在7月,为0.048 cm/min,是最小月份的3.0倍。两树种各月份平均液流速率大小依次为:尾叶桉8月>7月>10月>9月>6月>5月>1月>11月>4月>2月>3月>12月,湿加松为7月>8月>6月>9月>11月>5月>4月>12月>10月>3月>2月>1月,各月平均液流速率尾叶桉均大于湿加松,其中雨季月份两树种间差距小于旱季月份间的差距。

图2 不同月份尾叶桉、湿加松平均边材液流速率Fig.2 The monthly average of stem sap velocity in different month
3.2 两种速丰林边材液流速率与气象因子间的关系
树木边材液流速率的日变化特征主要受气象因子的影响,为了更好的分析液流速率与气象因子间的关系,分别选取雨季和旱季各连续6天(雨季7月15—20日,旱季11月10—15日;其中阴雨天气5天,晴天7天)的液流速率和气象因子数据进行分析作图(图3),通过Pearson相关分析我们得出,湿加松和尾叶桉均与空气温度、风速、太阳辐射、光合有效辐射和水汽压亏缺呈极显著正相关(P<0.01),而与相对湿度呈极显著负相关(P<0.01),与降雨量无显著相关性。其中湿加松各影响因子相关系数大小为VPD>相对湿度(负相关)>光合有效辐射>太阳辐射>大气温度>风速;尾叶桉为相对湿度(负相关)>VPD>光合有效辐射、太阳辐射>大气温度>风速。
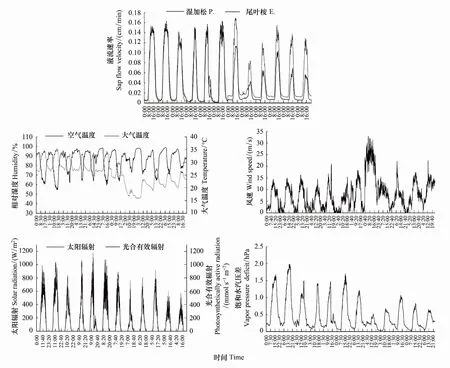
图3 湿加松和尾叶桉液流速率与气象因子日变化Fig.3 Relationship between stem sap flow rate and meteorological factors
为了进一步揭示环境因子对液流速率的综合影响建立树干液流速率与上述环境因子的多元线性模型采用逐步法进行回归分析,以5%和10% 的可靠性作为因变量入选和剔除临界值最终的回归模型参数见表2。
湿加松:VC=-0.0659+9.14×10-2VPD+2.66×10-4PAR-2.80×10-4SR+5.99×10-4RH+2.58×10-4Vwind;R2=0.95,n=1728
尾叶桉:VC=0.2482-2.58×10-3RH+8.31×10-5PAR+4.37×10-4Vwind;R2=0.924,n=1728
式中,VC为边材液流速率;PAR为光合有效辐射;VPD为水汽压亏缺;RH空气相对湿度;SR为太阳辐射;Vwind为风速。
3.3 林分的树木蒸腾耗水量估算
如表3所示,观测期间尾叶桉人工林年平均单株日耗水量为12.79 L/d,是湿加松的1.33倍;旱雨两季两树种均差异显著,其中尾叶桉雨季平均单株日耗水量为15.31 L/d,是旱季的1.50倍,湿加松雨季平均单株日耗水量为12.38 L/d,是旱季的1.81倍;两种速丰林各月的平均单株日耗水量均存在差异,其中尾叶桉最小为9.01 L/d,出现在12月份,最大为8月份16.92 L/d;湿加松平均单株日耗水量最小出现在1月份为5.64 L/d,最大值为7月份16.94 L/d。通过林分现存密度进行尺度转换我们得出:尾叶桉和湿加松人工林林分年蒸腾耗水量分别为582.16 mm和483.24 mm,分别占同期年降雨量的34.2%和28.4%。雨季尾叶桉林分蒸腾量占年总蒸腾量的3/5,是同期降雨量的23.4%,旱季蒸腾量为231.4 mm,是同期降雨量的113%;湿加松林分雨季蒸腾量为313.04 mm,是旱季的1.84倍,占同期降雨量的20.9%,旱雨两季蒸腾量分别为同期降雨量的20.9%和83.2%(见图4)。
*代表观测期间的日均耗水量

图4 湿加松、尾叶桉旱雨季林分蒸腾耗水量及同期降雨量比较/mmFig.4 Comparison of transpiration water consumption and rainfall during the same rainy season and dry season
4 结论与讨论
尾叶桉和湿加松边材液流速率受气象因素的瞬时影响,均表现出明显的昼夜变化规律:白天变化幅度大,呈明显的单峰型曲线。蒋文伟等[26]认为白天随着太阳辐射的增强,叶片气孔导度增大,植物气体交换加快,同时空气温度升高,叶片和空气中水汽压梯度增大,水分汽化加快,从而加快蒸腾,增大液流速率,很多学者的研究均证明了这一点[27-28]。两树种夜间均存在夜间液流且较白天微弱,依据Goldstein等[29]的理论,夜间液流活动是植物体为弥补日间蒸腾造成的水分亏缺,为植物体补充水分的过程。研究发现,两树种旱雨季夜间液流速率及变化幅度均显著大于后半夜,说明前半夜是两树种水分补充的主要时期。夜间液流的产生原因目前尚有争论,孙振伟等[19]、蒋文伟等[26]认为夜晚蒸腾作用为零,根压是夜间液流产生的主要因素。还有很多学者认为夜间液流的产生不仅仅是根压造成的,如果有足够的外部环境,夜间也会有明显的蒸腾作用[30],鱼腾飞等[31]对胡杨(Populuseuphratica)蒸腾耗水的研究中发现,胡杨夜间有明显的蒸腾作用,平均蒸腾速率为0.7 mmol m-2s-1。尾叶桉和湿加松夜间液流的产生原因,还需进一步分别对其夜间气孔蒸腾进行监测,同时对夜间液流速率与气象因素进行相关分析,从而确定夜间液流是否因单纯的夜间补水还是夜间蒸腾和树干补水同时进行造成的。
受气象因素和土壤供水等因素的影响,尾叶桉和湿加松各月平均液流速率均存在较大差异,尤其是旱雨两季间的日变化特征差异更大,两树种雨季启动时间均早于旱季,峰值、维持高液流速率的时间及日平均液流速率均雨季大于旱季。这与王志超等[20]对尾巨桉旱雨两季液流特征的研究结果保持一致。造成这一结果的原因主要是雷州半岛地区雨热同期,较高的太阳辐射、空气温度和充足的供水条件使得雨季蒸腾速率大于旱季。这在其他树种如大叶相思(Acaciaauriculiformis)[32]、马占相思(Acaciamangium)[33]的研究中也证明了这一观点。尾叶桉和湿加松两树种间日变化特征也存在较大差异,雨季两树种间差异较小,旱季差异较大,这主要是由树种本身的生物学特性决定的。
研究表明,尾叶桉和湿加松边材液流速率是多个气象因子综合影响的结果,两树种均与空气温度、风速、太阳辐射、光合有效辐射以及水汽压亏缺呈极显著正相关,与空气相对湿度呈极显著负相关,而与降雨量无显著相关性,这与刘国粹等[17]对白天尾叶桉与气象因子的关系研究结果保持一致。熊伟等[34]研究认为影响林木蒸腾的最主要的微气象因子是太阳辐射、空气相对湿度和气温,其中与冠层太阳辐射、饱和水气压差、气温呈正相关关系,与空气湿度呈负相关关系,本研究的结果也与此观点保持一致。
本研究对10年生尾叶桉和湿加松林分蒸腾耗水量的研究表明:尾叶桉年平均单株耗水量为12.79L/d(1.59 mm/d),湿加松为9.61 L/d(1.31 mm/d),林分年耗水量尾叶桉和湿加松分别为582.16 mm和483.24 mm,分别占同期降雨量的34.2%和28.4%。张宁南[35]对河头、纪家4a生尾叶桉人工林耗水量研究中两林分日均耗水量分别为1.49、1.53 mm/d,年耗水量为542、559 mm,与本研究10年生林分耗水量相当,原因是10年生尾叶桉日平均液流速率小于4年生液流速率,但边材面积却显著大于4年生,综合起来,平均单株日耗水量两林龄相差不大。湿加松蒸腾耗水研究目前较少,但对25年生加勒比松蒸腾耗水量研究发现其日耗水量为 0.35—3.52 mm/d[36];刘琪璟等[37]对20年生湿地松耗水量研究发现,其林分年蒸腾量为477 mm(1.31 mm/d)。
尾叶桉和湿加松旱雨季的蒸腾量也有较大差异,其中尾叶桉雨季蒸腾量为350.76 mm,占年总蒸腾量的60%,日平均耗水量为1.91 mm/d,是旱季日均耗水量的1.5倍;而湿加松雨季蒸腾量(313.04 mm)是旱季蒸腾量的1.84倍。张宁南等[35]对河头纪家尾叶桉旱雨季的蒸腾量研究中同样发现,两林分的日耗水量均是雨季显著大于旱季,这与本文的研究结果相一致。导致这一结果的原因是旱季土壤有效水的减少及较低的VPD等在不同程度上都限制了林木的耗水量。
猜你喜欢
绿色科技(2022年7期)2022-11-16
农业科技与信息(2021年8期)2021-05-17
北方果树(2020年6期)2020-11-14
中国水土保持科学(2019年6期)2019-04-26
现代园艺(2017年19期)2018-01-19
现代园艺(2017年21期)2018-01-03
农业环境科学学报(2017年2期)2017-03-20
Advances in Polar Science(2015年1期)2015-02-06
中国质量与标准导报(2014年5期)2014-02-28
储能科学与技术(2014年5期)2014-02-27
