
增温对青藏高原高寒草原生态系统碳交换的影响
2019-05-13 01:49干珠扎布胡国铮万运帆李玉娥旦久罗布白玛玉珍高清竹
生态学报 2019年6期
李 岩,干珠扎布,*,胡国铮,万运帆,李玉娥,旦久罗布,白玛玉珍,高清竹
1 中国农业科学院农业环境与可持续发展研究所,北京 1000812 农业部农业环境重点实验室,北京 1000813 那曲市草原站,那曲 852100
草地是重要的陆地生态系统碳库,草地生态系统碳循环是目前全球变暖研究中的热点。草地生态系统与大气之间的碳交换是碳循环的主要过程之一。碳交换组分包括总生态系统生产力(GEP,Gross ecosystem productivity)、净生态系统碳交换(NEE,Net ecosystem carbon exchange)、生态系统呼吸(ER,Ecosystem respiration)和土壤呼吸(SR,Soil respiration)等。气候变暖对草地生态系统碳交换各组分均有不同程度的影响,从而改变碳循环过程[1]。青藏高原平均海拔超过4000 m,被称为“世界第三极”[2],其面积约占中国陆地面积的 26.8%[3]。高寒草地生态系统是青藏高原分布最广、面积最大的生态系统,其自然条件复杂,生态环境脆弱,抵御灾害能力差,对气候变化极其敏感[4]。近年来,青藏高原气候变暖趋势明显,其增温幅度明显高于全球和全国平均水平,达到每10年0.32℃[5]。青藏高原高寒草地因其独特的地理位置和下垫面特点,气候变化导致的碳收支不平衡可能对我国中东部、亚洲其他地区乃至全球的气候产生极为重要的影响[6- 7]。
温度是限制青藏高原草地生产力的重要生态因子之一。温度通过影响植物的生长、繁殖、生物量分配,从而引起群落结构及物候等变化,进而间接影响生态系统的碳循环过程[8- 9]。目前,学者对水分条件较好的青藏高原高寒草甸碳循环做了大量研究[10-13]。亓伟伟等[10]认为相比土壤水分,温度对青藏高原高寒草甸碳通量的变化影响更大,增温使生态系统年平均总初级生产力增加了34.3%。Lin等[11]研究表明土壤温度是高寒草甸碳循环的主要控制因素,增温促进了生态系统呼吸和土壤呼吸,而土壤水分对呼吸作用无显著的影响。但也有研究认为,土壤水分在特定条件下对高寒草地生态系统碳交换具有显著的影响。例如,耿晓东等[12]发现增温幅度超过3℃时,将对土壤形成干旱胁迫,从而抑制GEP并降低NEE。Ganjurjav等[13]认为,增温尽管促进生长季平均GEP、ER和NEE,但在雨季来临之前,由于增温引起干旱,高寒草甸生态系统碳交换各组分均有不同程度的降低。综上所述,增温对水分条件较好的高寒草甸生态系统碳交换总体上具有促进作用,但增温超过一定阈值或者在干旱季节,增温将对生态系统产生干旱胁迫,从而抑制碳交换。因此,为了进一步明确增温对高寒草地碳交换的影响,本研究选取了水分条件较差的高寒草原生态系统,采用开顶增温室(Open Top Chamber,OTC)进行模拟增温,研究增温是否会对高寒草原产生干旱胁迫,是否对生态系统碳交换产生负面影响,为评估未来气候变化条件下青藏高原高寒草原碳储存能力提供理论依据。
1 材料与方法
1.1 试验区概况
试验区位于那曲市班戈县普保镇,地处念青唐古拉山和唐古拉山之间以及藏北高原纳木错、色林错两大湖泊之间,地理坐标为92°03′E,31°39′N,平均海拔4700 m。试验区为高原亚寒带半干旱季风气候,高寒缺氧,气候干燥,昼夜温差大,年均气温0℃,生长季(5—9月)平均气温为8.0℃;全年日照时数为2900 h,年降水量为289—390 mm,主要集中在6—9月份,年蒸发量为2000 mm。
1.2 试验设计
本试验采用开顶增温室对高寒草原进行全年模拟增温。该设备是以聚碳酸酯为原料的透明圆台,底部直径1.2 m,顶部直径0.6 m,高0.45 m,顶部和底部与外界相通,保持空气流动。OTC固定在样地内,进行全年增温。
本试验设置增温(W,Warming)和对照(CK,Control check)两个处理,每个处理重复4次,共8个小区,相邻试验样方间隔2 m的缓冲带。试验地位于地势较为平坦,群落结构较为均匀的自然高寒草原,主要建群种为禾本科植物紫花针茅(Stipapurpurea),其他伴生种包括银洽草(Koeleriaargentea)、羊茅(Festucaovina)、矮火绒草(Leontopodiumnanum)、小叶棘豆(Oxytropismicrophylla)等。土壤类型为高寒草原土,土壤质地为砂壤土。试验地于2011年6月进行围封禁牧,2011年7月开始进行增温试验,数据采集在2012—2014年6—9月份进行。
1.3 数据观测方法
采用温湿度记录器(北京佳乐宣科技有限公司)对高寒草原生态系统空气温度、土壤温度(5 cm)和土壤湿度(5 cm)进行全年动态观测(测定频率为15 min),以监测模拟增温条件下群落环境因子的变化。
采用Li- 6400便携式光合作用测定仪(LI-COR Inc.,Lincoln,NE,USA)和箱式法对高寒草原NEE和ER进行测定。实验开始前,将面积为30 cm×30 cm的底座嵌入各小区中,以备日后的定点测定。在植物生长季,选择晴朗天气在10:00―20:00(10:00之前露水较多,导致仪器测定效果不佳),每隔2 h进行1次测定,每个月测定1次。测定时将体积为30 cm ×30 cm×40 cm的透明有机玻璃箱放在底座上,箱子顶部采用风扇对箱内进行气体混匀,测定NEE,测定时间为90 s。NEE测定结束后,将箱子抬起,待箱内水分和CO2接近外界值时将箱子重新放在底座上,并用遮光布(内黑外白)盖在箱上,开始测定ER,测定时间同样为90 s。测定结束后通过NEE与ER之和计算出GEP。
土壤呼吸测定采用Portable Flux Meter(West systems,Italy)和箱式法测定。在每个小区内设置一个永久性土壤呼吸底座,并嵌入土壤。测定前24 h将底座内的绿色植物齐地面剪下,以消除植物冠层呼吸对土壤呼吸测定的影响。土壤呼吸测定选择晴朗天气,10:00—20:00进行动态观测,每2 h测定一次,每次测定时间为90 s,每个月测定1—2次,与NEE和ER同时测定。
地上生物量采用估算法测定:每年的生长季初(5月初),在试验地开顶增温室旁随机设置20个0.5 m×0.5 m矫正样方,每年的生长旺季(7月底或8月初),采用样方法在矫正样方内对每种植物进行群落特征调查,首先用直尺测定植物高度,随后在每个样方的冠层上方放置一个0.5 m×0.5 m的框架,框架中有100个均匀分布的网格(5 m×5 cm),然后估算每种植物的分盖度和群落总盖度。随后将植物齐地面剪下,分物种装入信封,在实验室烘箱中105℃杀青30 min,70℃烘干至恒重,测定干重。用每种植物的高度和盖度与所测生物量之间的关系建立回归方程,并运用该方程估算对照和增温处理的地上生物量[14],具体方程如下:
B=4.772H+0.408C-7.202
(1)
B=4.916H+1.228C-11.313
(2)
B=6.677H+1.015C-16.628
(3)
公式1、2、3分别为2012、2013和2014年生物量方程。其中,B代表生物量(g/m2),H代表植株高度(cm),C代表植物盖度(%)。
1.4 数据处理方法
采用IBM SPSS Statistics 19软件中单样本t检验分析比较不同处理间空气温度、土壤温度和土壤湿度的差异,采用重复测定方差分析比较不同处理对高寒草原GEP、ER、SR、NEE和地上生物量的影响,采用回归分析方法分析碳交换与空气温度、土壤温度、土壤湿度和地上生物量的关系。
2 结果与分析
2.1 高寒草原环境因子对模拟增温的响应
通过对青藏高原高寒草原生长季空气温度、土壤温度和土壤水分连续3年(2012—2014年)的观测,发现增温显著增加了空气温度、土壤温度并降低了土壤含水量(P<0.05)。2012年,2013年,2014年对照样地生长季空气平均温度分别为10.8,10.8,10.5℃,增温样地分别为12.8,12.5,12.5℃,较对照增加了2.0,1.7,2.0℃,增长率分别为18.5%,15.8%,19.0%(图1);同样,对照样地3年土壤温度分别为13.4,13.6,13.4℃,增温样地分别为15.3,14.5,15.0℃,较对照样地增加了1.9,0.9,1.6℃,增长率为14.2%,6.7%,11.9%(图1)。在土壤水分方面,增温样地的土壤含水率普遍低于对照处理(图1)。2012年,2013年,2014年对照样地生长季土壤含水率分别为11.99%,10.82%,10.77%,增温处理下分别为10.7%,10.55%,10.22%,较对照处理分别下降了1.29,0.27,0.55个百分点。在年际间,空气温度和土壤温度没有明显的变化,2013年,2014年的土壤含水率较2012年略有下降,在增温之后,3年之间的土壤含水率没有明显的变化。
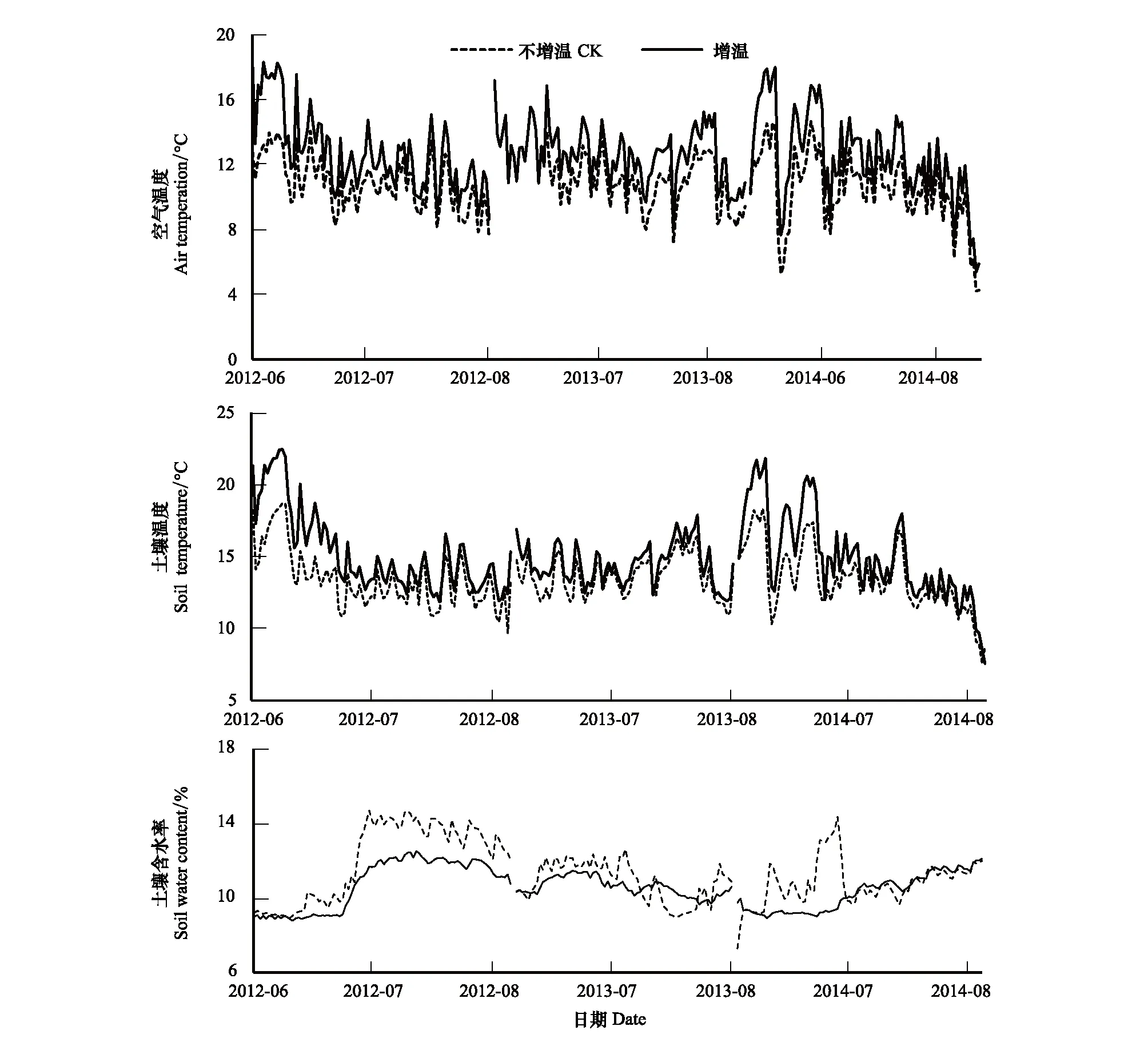
图1 增温对空气温度、土壤温度和土壤含水率的影响Fig.1 The air temperature and soil temperature and soil water content under warming conditionCK:不增温Control check;W:增温Warming
2.2 增温对高寒草原地上生物量的影响
增温对高寒草原地上生物量的影响存在年际差异,但总体上表现出其对地上生物量具有负效应(图2)。其中,2013年和2014年对照样地地上生物量分别为73.2 g/m2和66.1 g/m2,增温样地为58.8 g/m2和52.9 g/m2;较对照而言,增温处理地上生物量分别降低了19.7%和20.0%,达到显著性差异(P<0.05)。增温样地3年平均地上生物量为59.7 g/m2,对照样地为70.2 g/m2,增温使地上生物量降低了15.1%,达到显著性差异(P<0.05)。年际变化方面,对照样地内3年地上生物量分别为71.5,73.2 g/m2和66.1 g/m2,未达到显著性差异(P>0.05);增温样地内,2012年地上生物量最大,为67.4 g/m2,2014年最小,为52.9 g/m2,年际间差异性显著(P<0.05),表现出随着增温年限的增加,地上生物量逐渐降低的趋势。
2.3 增温对总生态系统生产力(GEP)的影响
在高寒草原,增温条件下不同年份GEP均低于对照(图3),其中,2013年和2014年达到显著性差异(P<0.05)。2013年和2014年,对照处理GEP分别为4.4 μmol m-2s-1和5.5 μmol m-2s-1,增温处理为2.6 μmol m-2s-1和3.1 μmol m-2s-1,增温处理GEP较对照分别减少了40.9%和43.6%。增温样地内3年平均GEP为3.8 μmol m-2s-1,对照样地为5.8 μmol m-2s-1,增温样地GEP较对照降低了36.8%,达到显著性差异(P<0.05)。在年际间,2012年的GEP显著高于2013年和2014年(P<0.05),对照样地内,2012年GEP分别是2013年和2014年的1.7倍和1.4倍,增温样地内,2012年的GEP是2013年和2014年的2.2倍和1.8倍。

图2 增温对生物量的影响Fig.2 The above-ground biomass under warming condition

图3 增温对总生态系统生产力的影响Fig.3 The gross ecosystem productivity under warming condition
2.4 增温对生态系统呼吸(ER)和土壤呼吸(SR)的影响
2012—2014年,增温条件下ER低于对照(图4),其中2013年达到显著水平(P<0.05),降低幅度达到22.7%。增温样地内3年平均ER为2.1 μmol m-2s-1,对照样地为2.6 μmol m-2s-1,增温样地ER较对照降低幅度达19.2%,达到显著性差异(P<0.05)。年际间,同GEP一致,较2012年,2013年和2014年的ER较弱,而在增温条件下其年际差异进一步增加。
高寒草原SR对增温的响应存在年际差异(图4)。2012年和2014年SR在增温与对照之间无显著差异(P>0.05),但在2013年,增温条件下SR显著降低(P<0.05),降幅为18.1%;增温样地内3年SR平均为1.5 μmol m-2s-1,对照样地为1.6 μmol m-2s-1,增温样地SR较对照降低了1.5%,未达到显著性差异(P>0.05)。在不同年份,增温条件下SR/ER均有所升高,但只有在2014年达到显著性水平(P<0.05),达到0.91,较对照的0.65增加了40.0%(图4);在增温条件下,3年平均SR/ER为0.8,较对照的0.6升高幅度达18.8%。
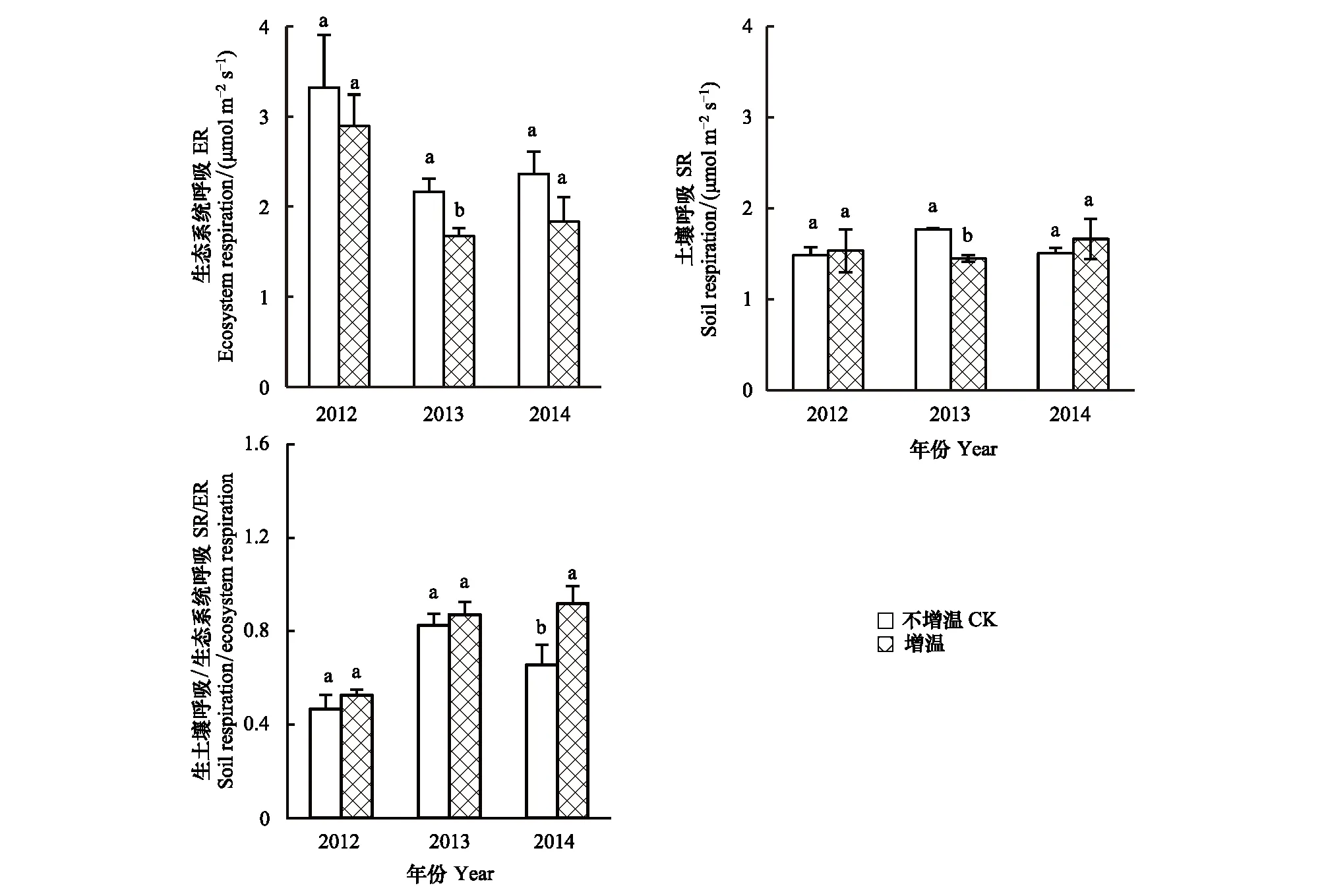
图4 增温对生态系统呼吸和土壤呼吸的影响Fig.4 The ecosystem respiration and soil respiration under warming condition
2.5 增温对净生态系统碳交换量(NEE)的影响
在高寒草原,增温对NEE的影响存在年际差异(图5),其中,2013年和2014年达到显著性差异(P<0.05),对照样地NEE分别为2.3,3.1 μmol m-2s-1,增温样地为0.9,1.2 μmol m-2s-1,增温较对照NEE分别降低了60.9%,61.3%;2012年对照样地内NEE为4.2 μmol m-2s-1,增温样地为2.7 μmol m-2s-1,增温较对照降低了35.7%。增温样地内3年NEE平均为1.6 μmol m-2s-1,对照样地为3.2 μmol m-2s-1,增温样地NEE平均降低幅度达51.5%,达到显著性水平(P<0.05)。年际变化方面,对照样地内3年NEE分别为4.2,2.3和3.1 μmol m-2s-1;增温样地内NEE分别为2.7,0.9,1.2 μmol m-2s-1,其中2013年和2014年NEE变化不显著(P>0.05),却显著低于2012年(P<0.05)。
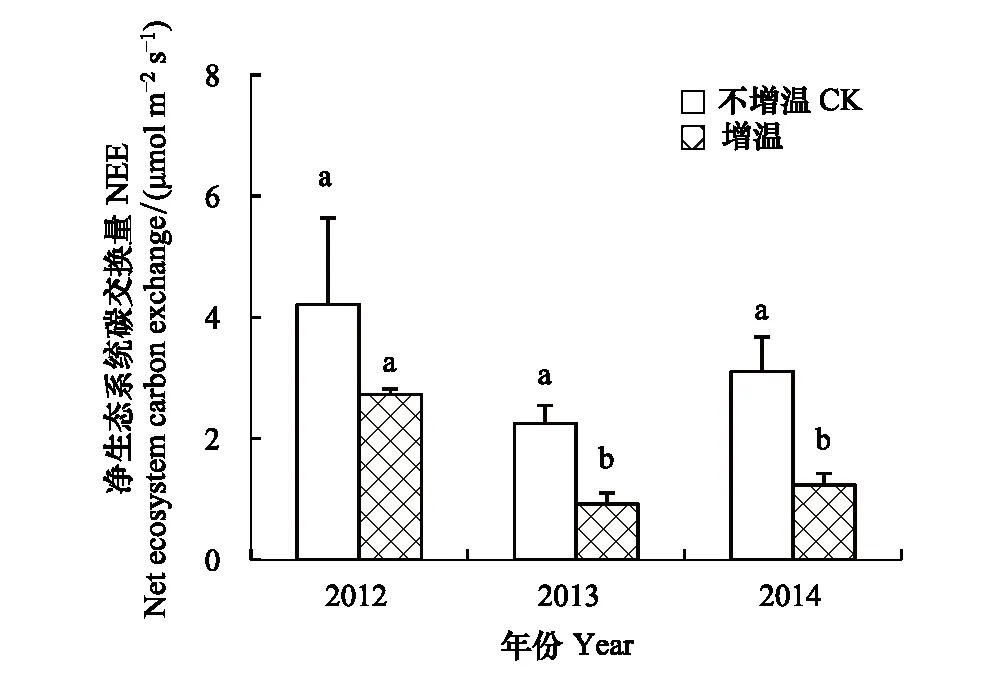
图5 增温对净生态系统碳交换量的影响Fig.5 The net ecosystem exchange under warming condition
2.6 青藏高原高寒草原碳交换与非生物因子和生物因子之间的关系
生长季内,青藏高原高寒草原GEP、ER、SR、NEE和地上生物量与土壤温度(ST,Soil temperature)、土壤水分(SW,Soil moisture)之间没有显著的相关关系(P>0.05,表1);与ST和SW不同,空气温度(AT,Air temperature)与GEP、ER、NEE和地上生物量之间有显著的负相关关系(P<0.05),其中,AT与GEP和NEE呈极显著的负相关关系(P<0.01); AT也与SR呈负相关关系,但没有达到显著水平(P>0.05)。
青藏高原高寒草原生态系统碳交换与地上生物量之间具有显著的线性相关关系(P<0.05)。地上生物量与GEP、ER和NEE之间呈极显著的正相关关系(P<0.01),它分别能解释38.6%、36.8%和35.8%的GEP、ER和NEE的变化;地上生物量与SR呈正相关,却没有达到显著性水平(P>0.05),它能解释7.3%的SR变化(图6)。
3 讨论
草地生态系统碳交换包括碳输出和输入两个过程,碳输出主要以生态系统呼吸为主,碳输入主要是指植物光合作用,在生态系统尺度上称为GEP[15]。植物光合作用的强弱决定了GEP的高低,而光合作用又受到环境温度的影响,适宜的温度范围内,增温有利于植物光合作用增强,而当环境温度超过植物的最适温度时,高温会破坏植物细胞原生质和光合膜结构[16],从而降低光合作用。此外,水分条件亦是限制植物光合作用的主要因素[17]。Wertin等[18]在北美通过对干旱群落3年的增温实验结果表明,温度升高显著降低了植物的光合作用。郭亚奇等[19]在藏北高寒草地的研究发现,增温使紫花针茅的光合速率下降,气孔导度降低,而水分增加可以促进紫花针茅的光合速率升高。也有研究认为由于在水分胁迫条件下植物光合作用受到影响,叶片气孔导度下降[20- 21],CO2进入细胞内的阻力增加,抑制了植物的光合作用[17],从而导致GEP在不同水分条件的生态系统中差异性较大。本研究结果表明,模拟增温显著降低了GEP(P<0.05),其原因可能是增温导致土壤含水率下降,对植物形成了干旱胁迫,从而降低了植物生产力,进而引起GEP下降。

表1 高寒草原碳通量与非生物因子之间的关系
*、**分别表示在95%、99%的置信水平上具有差异显著性;总生态系统生产力:GEP,Gross ecosystem productivity; 生态系统呼吸:ER,Ecosystem respiration; 土壤呼吸:SR,Soil respiration; 净生态系统生产力:NEE,Net ecosystem carbon exchange; 地上生物量:AGB,Above-ground biomass
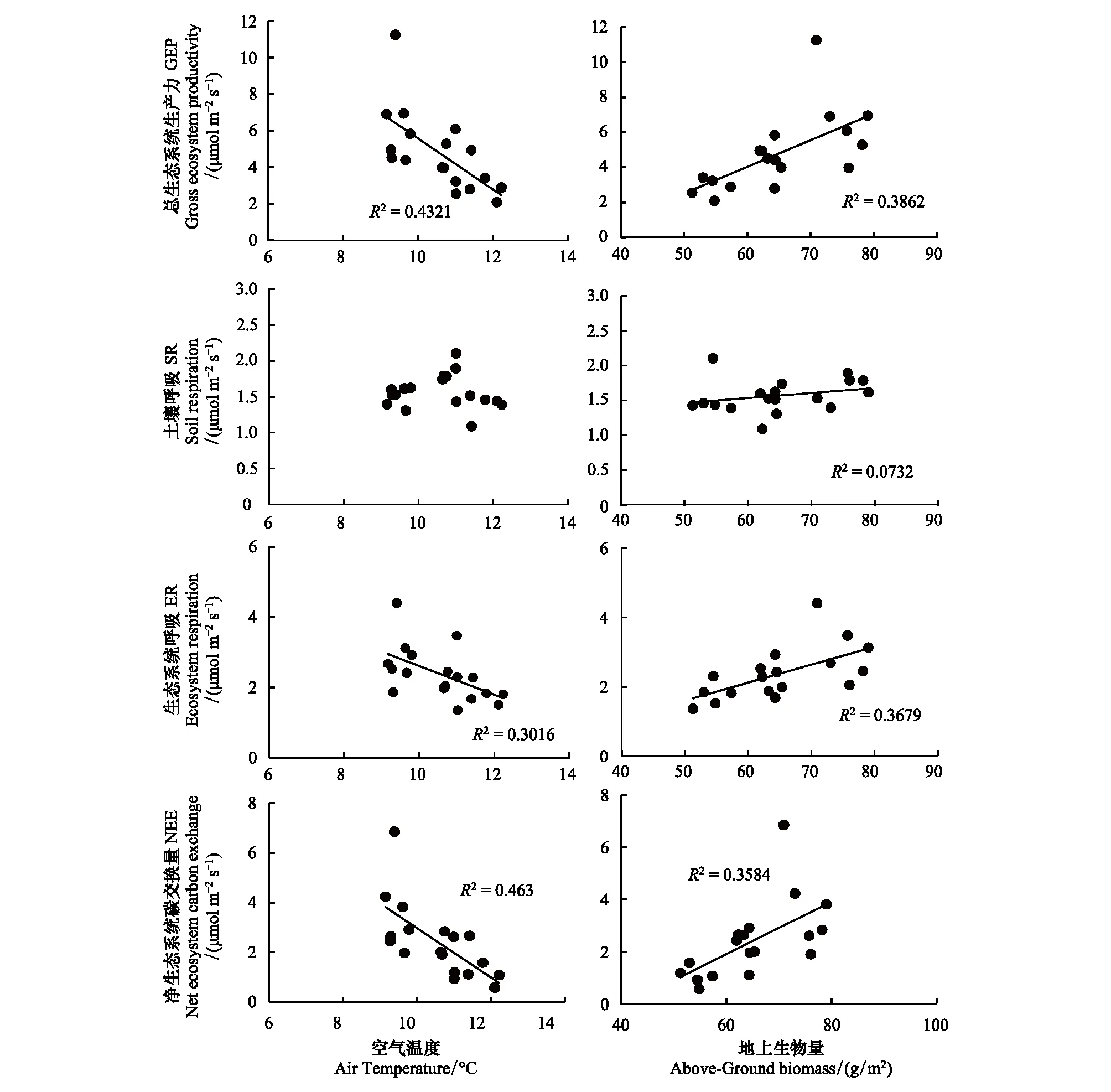
图6 总生态系统生产力、土壤呼吸、生态系统呼吸和净生态系统碳交换量与空气温度和地上生物量的关系Fig.6 Relationship between gross ecosystem productivity,soil respiration,ecosystem respiration and net ecosystem carbon exchange and air temperature and above-ground biomass
ER是碳交换过程中的重要环节,ER的强弱不仅关系着生态系统植物生产力的高低,而且对全球气候变化起着重要的反馈作用。一项在青藏高原的研究[22]结果表明,生长季前期,增温显著降低了ER,而在生长季后期,增温对其影响并不显著,这与生长季不同时期水分条件差异有关,既土壤温度和水分共同调节ER。Lin等[11]的研究结果显示增温对青藏高原高寒草甸生长季平均ER无显著影响,但显著促进生长季初期和末期ER,土壤温度与ER呈正相关,其解释了63%—83%的ER变化。本研究得出增温条件下,3年ER均有不同程度的降低,ER与空气温度呈显著的负相关关系(P<0.05),这与Fu的研究相似[23],其发现增温不仅降低了青藏高原高寒草甸ER,而且降低了ER的温度敏感性。SR是ER的主要组成部分,包括土壤中有机质分解、根系呼吸、及土壤动物和微生物呼吸,SR的强弱决定着土壤与大气之间碳交换能力的高低,而SR/ER能间接地反映出SR对生态系统碳输出的贡献。Lin等[11]的研究发现增温显著增加SR,土壤温度与SR呈正相关。本研究中,SR对增温的响应存在年际差异;2012年和2014年增温促进了SR,但2013年增温显著的抑制了SR(P<0.05),其原因可能是2013年土壤水分较低,水分胁迫抑制了SR。本研究中,增温并未显著改变SR/ER,这可能是ER的敏感度高于SR,抵消了SR的变化[11]。总体来看,增温对SR的影响相对较小,而对ER的影响较大,这主要取决于增温导致高寒草原地上生物量降低[24]。
生态系统呼吸与生态系统初级生产力之间的平衡称为生态系统净交换(NEE)。NEE是评价生态系统碳汇功能的重要指标。土壤温度和土壤水分是生态系统碳收支过程的主要控制因素[2,25]。Peng等[26]在青藏高原高寒草甸的研究中得出增温导致GEP的增加大于ER的减少,表现为NEE增加。但在本研究中,增温条件下高寒草原GEP和ER均有所下降,最终导致NEE降低,其主要原因是增温引起的干旱胁迫和地上生物量降低。Ganjurjav等[13]认为,虽然增温导致高寒草甸生长季平均NEE显著增加,但在雨季来临之前,增温显著降低高寒草甸NEE,说明增温条件下土壤水分是决定NEE的关键因素。Xia等[14]的研究结果表明,在干旱年份,温带草原NEE表现出对草地生物量的正向依赖性。因此,可以推测出全球气候变暖对青藏高原高寒草原碳汇功能具有消极的影响。
4 结论
通过模拟增温对青藏高原高寒草原碳交换的观测与分析,结果表明,增温对高寒草原碳交换各组分的影响存在年际差异,但总体上对碳交换存在负面影响。增温显著降低了高寒草原地上生物量、GEP、ER和NEE,而SR对增温的响应随着年份的变化而不同。GEP、ER、SR和NEE与土壤温度和土壤水分无显著相关,而GEP、ER和NEE与空气温度呈显著的负相关关系。NEE降低主要是土壤干旱胁迫和地上生物量降低引起的。因此,在未来气候变暖的环境下,青藏高原高寒草原的碳汇功能会有一定程度的减弱。
猜你喜欢
今日农业(2022年3期)2022-11-16
军事文摘(2021年20期)2021-11-10
空间科学学报(2021年2期)2021-07-21
科学大众(2020年10期)2020-07-24
水土保持研究(2019年6期)2019-10-19
中国水土保持科学(2017年3期)2017-07-05
浙江大学学报(工学版)(2016年2期)2016-06-05
中国学术期刊文摘(2016年1期)2016-02-13
中国学术期刊文摘(2016年8期)2016-02-13
应用海洋学学报(2015年4期)2015-11-24
