
模拟氮沉降对马尾松土壤微生物群落结构及温室气体释放的影响
2019-05-13 02:06宋利豪刘力诚赵泽尧
生态学报 2019年6期
张 雪,梅 莉,宋利豪,刘力诚,赵泽尧
华中农业大学园艺林学学院,湖北省林业信息工程技术研究中心,武汉 430070
土壤呼吸(Soil respiration)是全球温室气体释放的重要来源[1- 2]。即使土壤呼吸有较小比例的波动都会对大气组成及气候变化产生较大的影响[3- 5]。土壤呼吸对土壤环境变化非常敏感,如全球变暖、氮沉降、林分经营及植被破坏等都将影响土壤呼吸及温室气体的释放[6- 7]。森林是陆地生态系统的主体,因此研究影响森林土壤呼吸的因素及其驱动机制对预测全球变化背景下的森林碳循环以及森林的碳汇功能具有重要意义。
大气氮(N)沉降、施用氮肥的经营措施常常导致林地土壤的氮含量增加[8- 9],在此背景下,森林土壤环境、土壤温室气体释放对增氮的响应机制成为生态学关注的热点。以往研究表明,土壤氮增加对土壤温室气体释放的影响受多种因素调控,受立地条件、植被类型等影响而表现出不一致的结果。如对亚热带常绿阔叶林增施氮肥提高了土壤CO2的释放量[10],也有增施氮肥抑制了土壤呼吸[11]以及对土壤呼吸速率影响并不显著的报道[12],氮沉降及增施氮肥对温室气体释放的影响及其驱动机理有待进一步的研究。
根系呼吸不仅是土壤呼吸的重要组成部分,林木细根还可以通过改变土壤微环境而影响森林土壤的生物化学过程,进而成为驱动土壤温室气体(CO2、N2O)释放的重要因素[13]。研究表明,土壤呼吸与细根生物量呈显著正相关[14],在森林土壤氮增加的情况下,根系生物量和根长密度等也将发生变化[15],这种变化又直接或间接改变根际土壤矿质养分及微生物群落结构等,进而影响土壤温室气体的释放。土壤呼吸速率随根系生物量的增加而增加[16- 17],去除根系使土壤呼吸速率大幅降低[18]。根系养分含量的差异也会改变土壤的呼吸速率,如Volder等[19]研究发现,根系氮含量降低会通过减少根系对土壤养分以及水分的吸收作用来降低土壤与根系的呼吸速率,而根系氮含量增加有利于根系的呼吸[20]。然而,在全球氮沉降及林地施肥背景下,根系生物量及氮含量的差异及土壤养分、微生物等理化环境对施肥的响应,以及这种响应如何影响土壤呼吸及温室气体释放尚有待进一步研究。
马尾松(Pinusmassoniana)具有生长快、适应性强等特征,是我国南方最重要的造林树种之一,在区域生态环境建设和林产品供给方面发挥重要作用。本研究以马尾松土壤为对象,通过施氮肥和不同种植密度处理,研究在不同根系生物量条件下,增加土壤氮对马尾松土壤理化性质、微生物群落结构及温室气体释放的影响。旨在阐明氮沉降及施氮肥对温室气体释放的驱动机理,并为马尾松人工林的科学经营提供依据。
1 材料与方法
1.1 材料与处理
2016年10月,将2年生马尾松播种容器苗分别1株和3株移栽入5 L花盆,栽培基质是用河沙∶蛭石∶泥炭土∶黄泥土1∶1∶1∶1比例均匀混合,基质pH值6.85±0.11,全氮2.39±0.09 g/kg、全磷0.56±0.01 g/kg、全钾6.43±0.26 g/kg、有机质64.32±4.33 g/kg。为了研究不同根系生物量对土壤理化性质和土壤呼吸的影响,本实验分单株和3株混合栽植,并分别进行不施肥和施肥,共4个处理。施肥处理:于2017年4—6月分3次施入氮肥(尿素)溶液,共施入180 mg N/盆(等同于林地10 g N/m2);不施肥处理浇同样体积的水,以6月22日最后一次施肥为处理开始时间。每个处理30盆,共计120盆。
1.2 植物和土壤取样与分析
2017年6月18日开始取土壤和植物样品,每周进行一次植物取样,每个处理每次取4盆重复;实验7天和30天后分别取土壤和植物样品,每次每处理取3个重复。先将地上部分茎叶剪下,再将花盆倒扣取出根系植株分根、茎、叶进行保存。植株取出时,将根系附近的土壤轻轻抖落混匀,过2 mm土壤筛,后装于密封袋并放置于4℃冰箱保存,于1个月内测定土壤pH值、速效氮、全氮等理化指标。将取回的植株洗净后按根、茎、叶分别装于信封中,并放在65 ℃烘箱中烘干至恒重,称量获得干重。烘干后的植物粉碎过0.5 mm(40目)筛后进行保存,用于测定根、茎、叶的非结构性碳水化合物(TNC)和全氮含量。
土壤pH值测定采用水浸提-电位法测定;土壤铵态氮与硝态氮含量利用流动分析仪进行测定;植物全氮采用凯式定氮法测定;土壤微生物量碳(Soil microbial biomass carbon,SMBC)、土壤微生物量氮(Soil microbial biomass nitrogen,SMBN)含量采用氯仿熏蒸-氯化钾浸提,用TOC仪进行测定[21];土壤微生物群落结构采用磷脂脂肪酸法(Phospholopid Fatty Acid,PLFA)进行分析。
1.3 土壤呼吸速率及温湿度测定
土壤CO2和N2O释放速率的测定采用静态箱-气象色谱法进行测定。每个处理随机安装5个重复的土壤静态箱。静态箱由聚氯乙烯(PVC)管制成(规格:口径275 mm,高200 mm,底径180 mm),于实验开始前1周按压入土壤5 cm,周围土壤紧密压实防止漏气。于2017年6月18日—2017年7月11日期间每2天取1次,于2017年7月11日—2017年7月18日期间每3天取1次,于2017年7月18日—2017年8月18日期间每15天取1次。
于8:00—9:00 am密封PVC管,分别于密封前、密封培养两小时后取气,每次每处理随机取3个重复,将取出的气体装入真空瓶带回实验室,立即用气相色谱仪(Agilent 7890A)测定CO2、N2O的浓度,并计算土壤单位面积单位时间内的CO2、N2O的释放速率。
土壤CO2、N2O释放速率计算方法:
F=ρ×V/M×Δc/Δt×273/T×α
式中:F为CO2、N2O的释放速率,正值为释放,负值为吸收,ρ为标准状况下气体的密度,CO2、N2O的密度分别为1.978 kg/m3、1.98 kg/m3,V是采样箱体积(m3),M为采样箱底部土壤干重(g),Δc/Δt为在特定时间内的气体浓度变化速率,密闭时间2 h;T为采样点的绝对温度:开氏温度;α分别为N2O换算到N(28/44)、CO2换算到C(12/44)的转换因子;N2O需要乘以1000,得到单位是mg kg-1h-1,CO2直接得到单位是mg kg-1h-1。
1.4 数据处理与分析
应用SPSS19.0软件进行单因素方差分析,当方差分析结果表现为显著性差异时,通过Turkey检验进行不同处理间的多重比较,用Pearson法对土壤呼吸CO2、N2O与各影响因子进行相关性分析,利用逐步回归分析确定影响土壤温室气体释放的主要控制因子,使用Origin 9.0绘图。
2 结果与分析
2.1 植株生物量和氮含量
处理30天后,单株种植的马尾松苗地上、地下部分生物量均小于3株种植的总生物量。不施肥处理下,单株的叶片和根系氮含量均显著大于3株种植。施氮处理对单株种植幼苗各部分生物量无显著影响,但显著降低了3株种植地上部分生物量(表1)。无论是单株还是3株种植,增施氮肥均显著增加了根、茎、叶的氮含量(表1)。
2.2 土壤pH值及速效氮
无论是对照还是施肥在施氮肥处理7天后,单株种植土壤pH值显著降低(P<0.05),3株种植平均土壤pH值也有所降低,但差异不显著(P>0.05);施氮肥处理后,随着时间的延长至30天,各处理土壤pH值差异不显著。单株种植的土壤速效氮含量高于3株,施氮肥显著增加了单株和3株种植的土壤硝态氮含量,而对土壤铵态氮含量影响不显著(表2);随着处理时间延长至30天后,土壤硝态氮含量显著下降,但施肥处理仍显著大于未施肥处理。处理7天后,3株种植的土壤速效氮含量显著低于单株种植速效氮含量;处理30天后,单株和3株对照之间、单株和3株种植施肥之间速效氮差异不显著(表2)。

表1 不同处理马尾松幼苗生物量和氮含量的影响(均值±标准误)(n=3)
顺序调整为:SCK,单株对照; SF,单株施肥;TCK,3 株对照; TF,3 株施肥;字母相同表示在P=0.05(Turkey 检验)水平无显著性差异
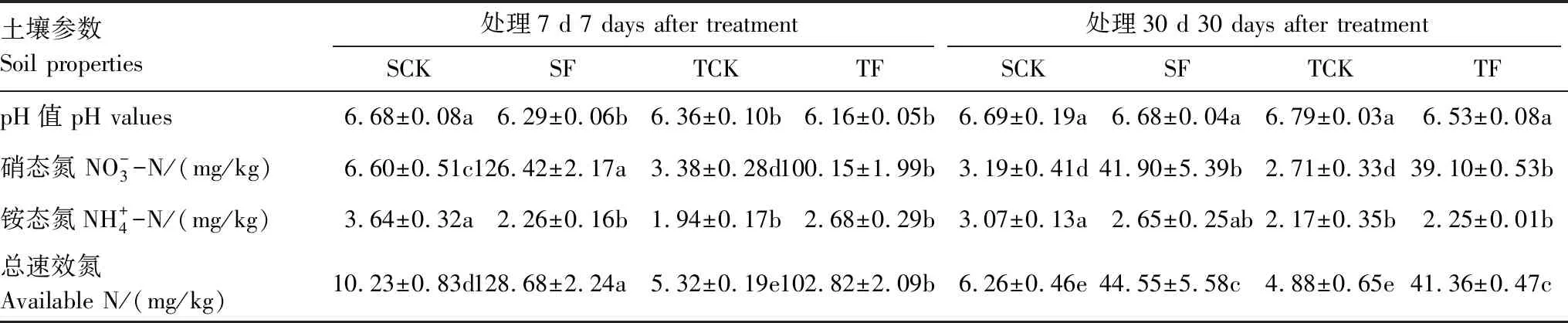
表2 马尾松幼苗土壤pH 值及速效氮含量(均值±标准误)(n=3)
速效氮=硝态氮+铵态氮
2.3 土壤微生物量碳、微生物量氮含量及微生物群落结构
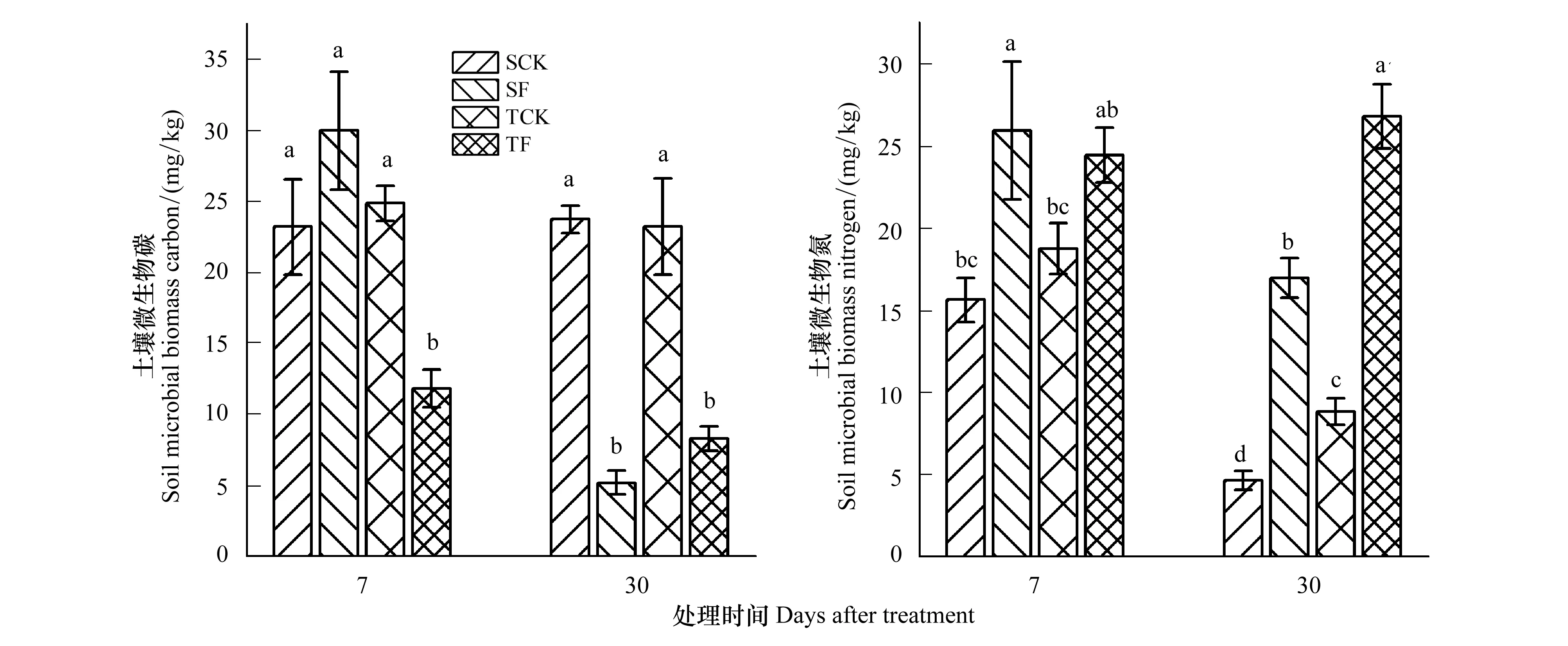
图1 施肥对马尾松幼苗土壤微生物量碳、微生物量氮的影响Fig.1 Effects of N fertilization on soil microbial biomass carbon(SMBC)and soil microbial biomass nitrogen(SMBN)contents of Pinus massoniana seedlings SCK,单株对照; SF,单株施肥;TCK,3株对照; TF,3株施肥。字母相同表示在P=0.05(Turkey 检验)水平无显著性差异,n=3
除单株种植处理7天时SMBC与对照没有显著差异外,施氮肥均显著降低了SMBC含量(图1A)。在施氮肥处理7天和30天时,无论是单株还是3株种植,施肥后SMBN含量均显著增加。处理时间越长,施肥与对照处理的SMBC和SMBN含量差异越大(图1B)。

表3 施氮肥对马尾松幼苗土壤微生物群落结构的影响(平均值±标准误)(n=3)
施氮肥处理7天后,单株和3株种植的土壤微生物群落都有下降趋势,其中3株种植革兰氏阳性菌、革兰氏阴性菌、厌氧菌、放线菌、真菌含量显著降低,而单株种植及3株种植的丛枝菌根真菌差异不显著。在施肥处理30天后,单株与3株种植土壤微生物群落总含量仍然比未施肥低,其中施肥后3株种植的土壤革兰氏阳性菌、革兰氏阴性菌、厌氧菌、放线菌、丛枝菌根真菌、真菌、真核菌含量均显著降低(表3)。3株种植的土壤各种微生物均低于单株种植土壤,在施肥处理30天后,革兰氏阳性菌、革兰氏阴性菌、厌氧菌、放线菌、丛枝菌根真菌显著降低。
2.4 土壤CO2、N2O释放速率
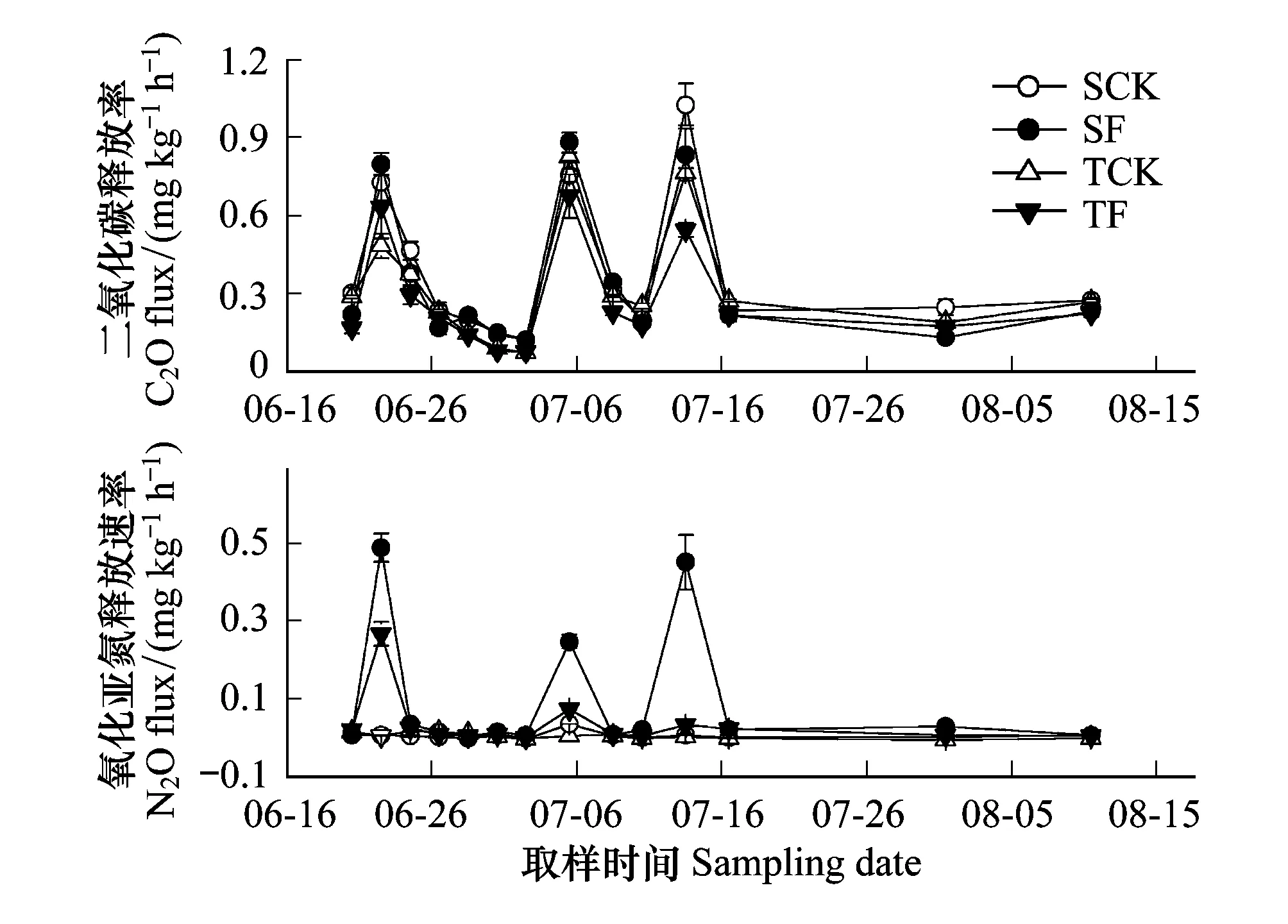
图2 施氮肥对土壤CO2和N2O释放速率的影响(n=3) Fig.2 Effects of nitrogen fertilization on soil CO2 and N2O flux(n=3)
无论是单株还是3株种植,施肥1天后(6月23日)土壤CO2释放速率出现峰值,施氮肥处理的土壤CO2释放速率显著高于未施肥处理。但随处理时间的推移,施肥处理的土壤CO2释放速率与对照差异不显著。处理7天至11天(6月29—7月3日),单株对照和施肥处理CO2释放速率均显著高于3株。而处理3周后(7月14日),施肥处理的土壤CO2释放量低于对照(图2)。施氮肥处理后,显著增加了1株与3株栽植的土壤N2O释放速率,施氮肥处理的N2O气体释放峰值时显著高于对照,随土壤含水量降低,N2O释放量也迅速下降(图2)。无论是单株还是3株种植,施肥处理土壤CO2和N2O释放的峰值出现的时间一致,均出现在浇水后一天。无论是施肥与不施肥处理,其CO2释放速率波动趋势一致,在峰值后有逐渐下降的趋势,在浇水后又急剧回升。但单株和3株施肥处理的N2O释放量在浇水后1天出现峰值,而后迅速降低;未施肥处理N2O释放量对浇水的响应不明显,在监测期内没有显著的波动(图2B)。
2.5 土壤CO2、N2O释放释放的主要影响因素
相关分析表明,土壤CO2释放速率与N2O释放速率相关,土壤CO2释放速率受SMBC、土壤温度、pH值和硝态氮含量等因子的显著影响;土壤N2O释放速率主要影响因子有SMBC、SMBN、土壤湿度、硝态氮及铵态氮含量。土壤CO2释放速率与土壤温度及硝态氮含量呈显著负相关(P<0.05),与土壤SMBC和pH值呈极显著正相关(P<0.01),土壤N2O释放释放速率与SMBN、土壤温度及硝态氮显著正相关,与土壤SMBC、铵态氮及pH值显著负相关(表4)。SMBC与土壤温度显著负相关;而SMBN与土壤pH 值显著负相关,与SMBN、土壤温度、土壤硝态氮含量显著正相关。根系生物量多少不仅显著影响SMBN及土壤湿度,而且对土壤铵态氮含量也有显著影响(表4)。逐步回归分析表明,土壤硝态氮含量是影响CO2(R2=0.334)和N2O (R2=0.924)释放的关键因子。
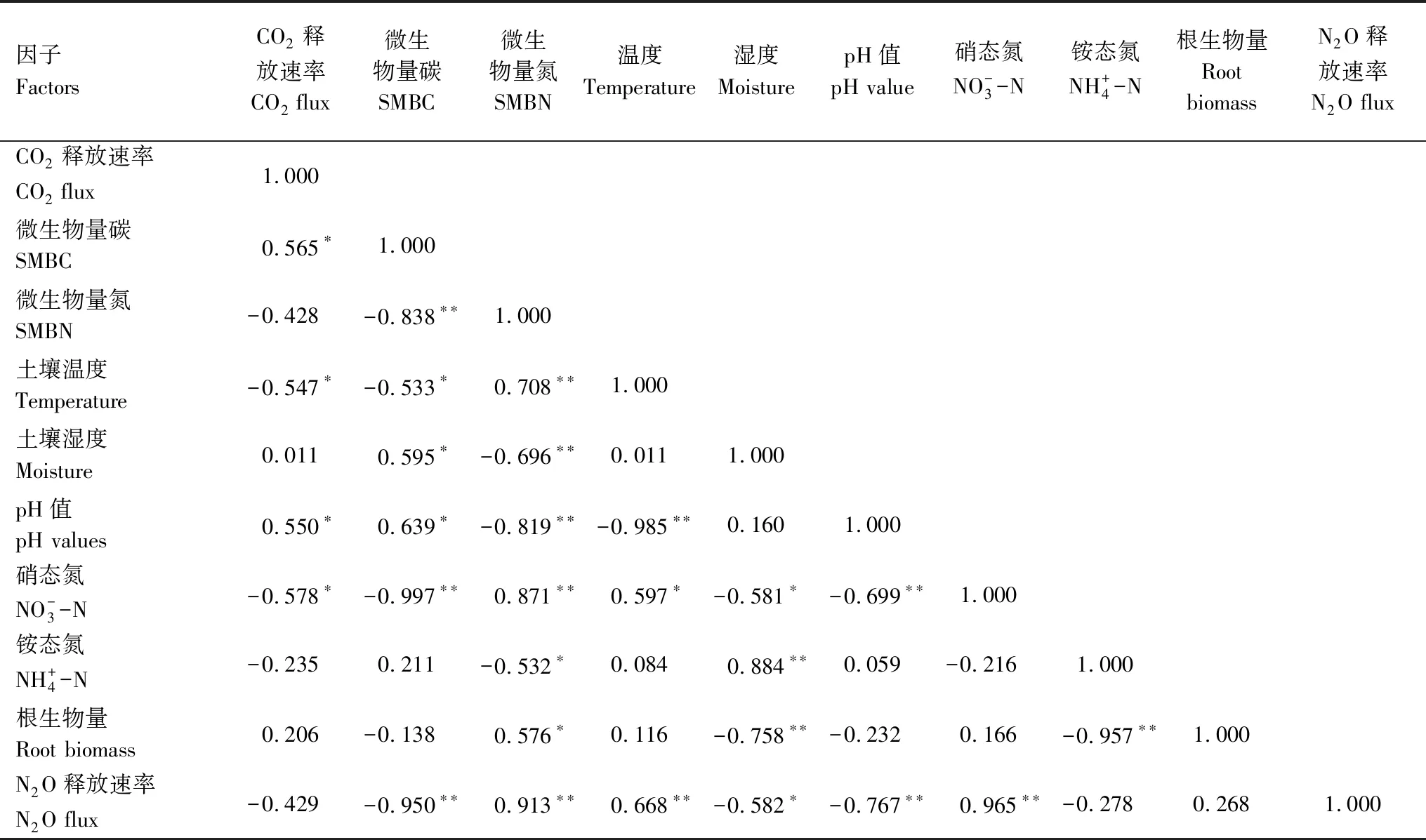
表4 土壤CO2和N2O释放速率与土壤环境因子的相互关系
* 在 0.05 水平(双侧)上显著相关,** 在 0.01 水平(双侧)上显著相关,n=24
3 讨论
3.1 施氮肥对SMBC、SMBN及微生物群落结构的影响
SMBC是土壤碳氮循环途径的关键因子,其含量的变化是预测土壤肥力、土壤微生物活性以及土壤碳氮周转速度的重要指标[22]。本研究模拟氮沉降显著降低了SMBC含量,土壤氮含量显著增加改变了土壤微环境,导致土壤C/N比降低,而较低的C/N有利于土壤有机碳的分解释放,进而导致土壤有机碳的逐渐减少[23];土壤微生物的活性会随着有机碳降低而减弱,也可能是导致SMBC含量降低的原因之一[24]。SMBN的变化是衡量土壤氮素循环周转的关键性因子[25],本研究中施氮肥增加了土壤氮含量和SMBN含量。土壤氮含量的增加提高了土壤微生物氮循环所需的底物,导致微生物活性变强,因此可能是提高SMBN含量的主要原因。
施氮肥处理降低了土壤微生物群落结构总含量,特别是土壤中细菌的含量显著下降,刘桂要[26]对油松(Pinustabulaeformis)林的研究结果也表明,添加氮处理对土壤细菌群落结构组成的影响程度大于真菌。施氮导致单株种植总微生物量下降13.3%,3株种植比单株种植下降更为显著,下降了29.5%。这与Ramirez等[27]对北美洲28个生态系统中的土壤氮沉降使土壤微生物量下降35%的研究结果基本一致。适量的施氮肥可改善土壤理化性质及微生物活性等,但氮施用过量则会造成土壤微生物多样性降低[28- 29]。施氮肥后的土壤速效氮含量在41.36—126.68 mg/kg,超过植物正常需求(10—20 mg/kg),进一步表明过量施用氮肥会导致土壤微生物量下降。洪丕征等[30]模拟氮沉降对红椎(CastanopsishystrixMiq.)人工幼林土壤影响的研究发现,氮沉降导致土壤微生物下降的主要机制是土壤pH值的降低,因为土壤pH值过低会抑制细菌生长[31],这与本研究中土壤pH值与土壤SMBC显著相关的结果也是一致的(表4)。增氮还会通过改变微生物对碳源的利用效率而对土壤微生物量产生影响,一般在高氮处理下,土壤微生物对碳源的利用能力降低[32-33]。氮对土壤微生物群落结构的影响机制是复杂的,需要综合考虑土壤氮的增加量以及氮增加对土壤pH值、土壤C/N、土壤微生物碳源利用效率等的影响等。
3.2 施氮肥对土壤温室气体释放的影响
模拟氮沉降短时间内提高了土壤CO2的释放速率,而后土壤CO2释放与对照相比有降低趋势或无显著差异。这可能是因为在施肥后土壤C/N降低,加快了土壤CO2的释放速率[34]。施肥提高了土壤中有效氮的含量,提高了土壤酶活性,为微生物生命活动提供充足的底物,从而在短时间内达到促进土壤呼吸的作用,但这种促进作用会随着时间的推移而渐减弱甚至消失[35- 36]。也有相关研究证明,长期的施氮肥减少了微生物的呼吸,从而降低CO2的释放速率[35]。Liang等[37]对农田生态系统的研究表明,施肥能够提高土壤中根系的自养呼吸,且自养呼吸的响应快于异养呼吸。施肥后,根系呼吸快速反应而对提高CO2的释放速率有一定贡献,但随时间推移,由于过量施肥处理的土壤微生物群落数量的显著下降,导致土壤异养呼吸显著下降,从而掩盖了根系呼吸的贡献量。
增施氮肥显著提高了土壤N2O释放速率,这与Jassal等[38]对森林土壤施氮肥后的研究结果一致。本研究中施氮肥后土壤硝态氮含量显著增加,而且施肥降低了土壤SMBC含量,增加了土壤SMBN含量,土壤微生物群落总量显著下降,这些因素的响应均与N2O释放速率呈显著相关。相关研究表明,增施氮肥也会通过降低土壤pH值,改变土壤的微生物活性,进而改变土壤的呼吸速率[39]。氮肥施入土壤后,经矿化过程以硝酸盐的形式存在于土壤中,增加了土壤硝化的底物[40];施肥后土壤C/N较低时,也可以降低反硝化微生物对氮素的竞争力,从而减缓土壤的反硝化过程[41-42],施肥后硝化底物的增加及反硝化过程的减缓是N2O释放速率增加的重要原因。
相关性分析表明,与土壤CO2释放速率相关的因子主要有SMBC、土壤温度、土壤pH值及硝态氮含量,逐步回归分析表明土壤硝态氮含量是土壤CO2释放速率的决定性因子。土壤N2O释放速率与SMBC、SMBN、土壤温度、土壤湿度、pH值及土壤硝态氮含量均呈显著相关,逐步回归分析发现土壤N2O释放速率主要取决于硝态氮含量。在以往研究中,土壤温度与土壤湿度是土壤CO2释放速率的主要影响因素[43-44],刘顺[45]对川西亚不同森林类型的研究结果表明,土壤呼吸和总硝化速率与土壤温度显著相关,而与土壤水分相关性不显著。土壤水分是通过影响根和微生物活动、底物的扩散和土壤氧气浓度等而直接和间接的影响土壤碳氮的转化速率[46- 47],当土壤水分不是环境限制因子时,土壤温度通常是影响土壤呼吸的重要影响因子[45]。而本研究中,虽然土壤温度与CO2和N2O的释放呈显著正相关,土壤湿度与N2O的释放显著负相关,但逐步回归分析表明影响CO2和N2O的释放速率的决定性因子是土壤硝态氮含量。这些研究结果说明,在不同地区、不同森林类型及气候条件下,林地土壤环境的复杂性及限制因子的不同,可能是温室气体释放关键因素差异的原因。
3.3 根系生物量与土壤温室气体释放的关系
无论是对照还是施肥处理,3株(根系生物量增加)栽植土壤中的速效氮含量均显著降低(表2),可能是因根系对养分的吸收利用增加而减少了土壤中的氮含量,土壤N2O释放速率因根系增加而显著下降。另外,3株种植根系生物量高,对土壤水分的消耗也较单株高,因而显著降低了土壤水分含量,土壤湿度的降低通过影响土壤温度及SMBN,直接或间接地影响土壤温室气体的释放速率。土壤温室气体释放速率随着随根系生物量的增加而增加[16- 17],在未施肥处理条件下,根系生物量较高的3株处理其土壤微生物群落生物量也较高,也会对土壤温室气体产生一定的影响。
根系呼吸与根系氮浓度具有极大的相关性[48],施肥处理导致根系氮含量显著增加,这在一定程度上可能会增加根系的呼吸速率[20]。但由于过量施肥显著降低了土壤微生物群落结构,土壤微生物呼吸显著下降[35],这在一定程度上掩盖了根系生物量及根系氮浓度对土壤呼吸的贡献量。
猜你喜欢
南方农业(2022年17期)2022-10-21
农业科技通讯(2022年7期)2022-07-19
安徽农学通报(2022年6期)2022-04-07
水土保持研究(2019年6期)2019-10-19
畜牧与饲料科学(2018年3期)2018-05-08
中国水土保持科学(2017年3期)2017-07-05
浙江大学学报(工学版)(2016年2期)2016-06-05
植物营养与肥料学报(2012年1期)2012-10-26
植物营养与肥料学报(2011年2期)2011-10-26
植物营养与肥料学报(2010年3期)2010-11-16
