
β-氨基丁酸对吊竹梅的气孔运动影响机制初探
2019-05-13 08:24:44刘阿萍关思静胡本祥颜永刚
陕西农业科学 2019年4期
高 静,王 楠,刘阿萍,陈 莹,关思静,胡本祥,颜永刚,张 岗
(1.陕西中医药大学 药学院,陕西 西安 712046;2.陕西中医药大学 陕西省中药资源产业化协同创新中心,陕西 咸阳 712000)
吊竹梅(TradescantiazebrinaBosse)是园林重要观赏植物,同时亦可作药用,有清热凉血、解毒、利尿的功能[1],现在大多采用扦插繁殖,但在扦插繁殖中易发生蒸腾失水,导致植物萎蔫。气孔是蒸腾失水的重要通道,湿度、温度、CO2浓度、光照、微生物等许多因素都可影响气孔运动,NO、活性氧(ROS)和Ca2+等信号分子参与气孔运动的调节[2]。
β-氨基丁酸(BABA)是一种极具潜力的广谱性植物化学诱抗剂,不仅可以增强植物的抗病性,还可以提高植物抵抗干旱、高温、高盐和重金属等非生物胁迫的能力[3]。最新的研究显示BABA可增强酵母诱导的气孔关闭现象[4]。笔者研究探索BABA对吊竹梅气孔运动的影响,为后续BABA作为气孔抑制剂适度提高水分利用率,促进扦插苗的成活率提供一定参考价值。
1 材料与方法
1.1 实验材料
将吊竹梅种植在培养土(营养土、蛭石、珍珠岩1:1:1混合配置)中,温室培养,适当浇水,保持土壤湿润即可[5]。
1.2 实验试剂
BABA、KCl、2-(N-吗啉代)乙磺酸(MES)、L-抗坏血酸(AsA)、Ca2+螯合剂(EGTA)、Ca2+通道抑制剂(LaCl3)、还原型谷胱甘肽(GSH)、细胞壁过氧化物酶抑制剂(SHAM)、NO清除剂(c-PTIO)和NO合成酶抑制剂(L-NAME)均购自上海源叶生物技术有限公司。
1.3 实验方法
1.3.1 撕取表皮条 取长势良好的吊竹梅植株,并取顶端的第二片完全展开的叶片,用镊子轻轻撕取下表皮,放入MES-KCl(MES 10 mM,KCl 50 mM,CaCl2100μM pH 6.15)缓冲液中。
1.3.2 气孔试验 主要有:
(1)诱导气孔关闭:用镊子轻轻撕取待测叶片的下表皮条,将其转移至MES-KCl缓冲液中,并在光(400μmol·m-2·s-1)下处理 2 h,使其气孔完全打开。然后将部分表皮条转入不同浓度(10 μM、100 μM、250 μM)BABA处理液中,并继续在光下照射。1 h 后于显微镜下观察,并拍照记录。
(2)抑制剂处理的表皮条实验:撕取待测植株的表皮条,将其转移至将MES-KCl缓冲液中,并在光(400μmoll·m-2·s-1)下处理 2 h。然后将部分表皮条转入含有信号抑制剂AsA、EGTA、LaCl3、GSH、SHAM、c-PTIO、NaN3和L-NAME的MES-KCl缓冲液中,并在光下继续照射0.5 h。再将抑制剂处理过的表皮条移入BABA溶液中光照1 h 后于显微镜下观察,拍照记录。
1.4 气孔开度测量
每个实验处理至少选取120个气孔进行测销量。用Image-Pro Plus 6.0(Media Cybernetics, Silver Springs, MD)测定线段的方法测量气孔开度。
1.5 数据统计
使用 Spss 20.0 软件进行数据分析,所有数据图均由 Origin 8.0 完成。
2 结果
2.1 BABA诱导吊竹梅下表皮气孔关闭
由图1看出在经过充分的光照后,吊竹梅的下表皮气孔完全打开,对照组的气孔开度达到了10.42μm。在用BABA处理1 h后,气孔开度明显下降,10 μM、100 μM、250 μM分别使气孔开度与对照相比分别下降了26.3%、35.3%和30.5%。100 μM BABA处理过的吊竹梅气孔开度最小,随着BABA浓度增加到250 μM,减小的趋势减弱。
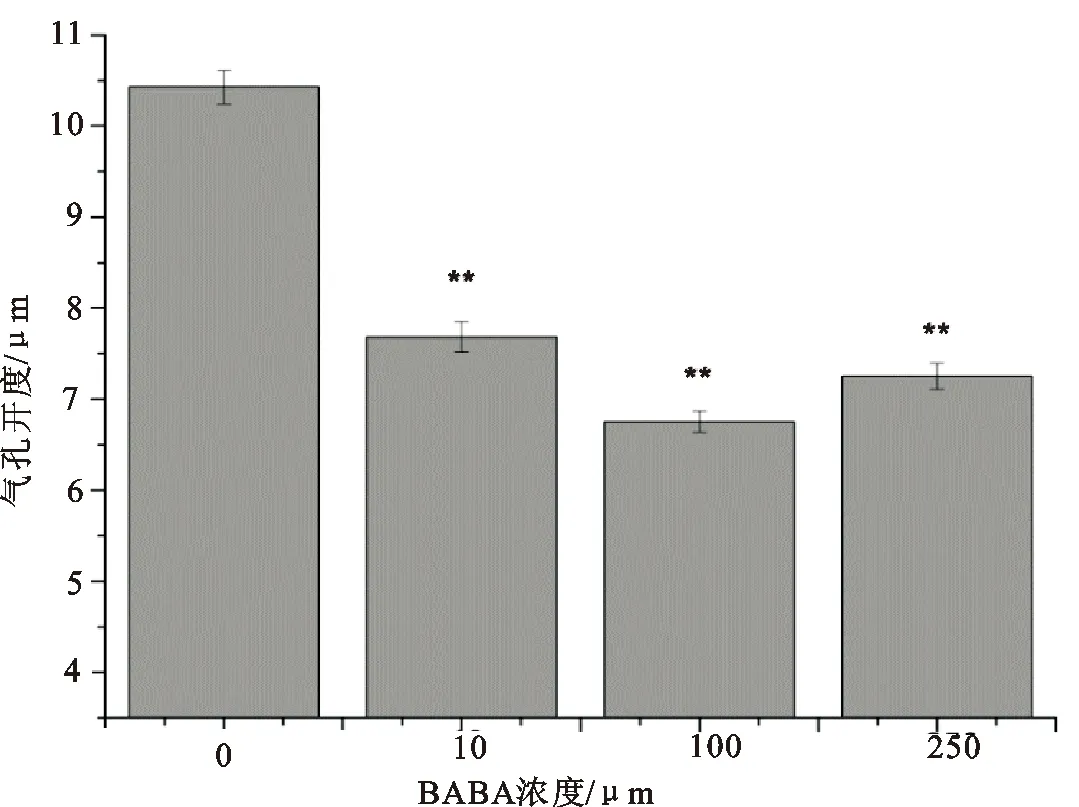
图1 不同浓度BABA诱导的气孔关闭
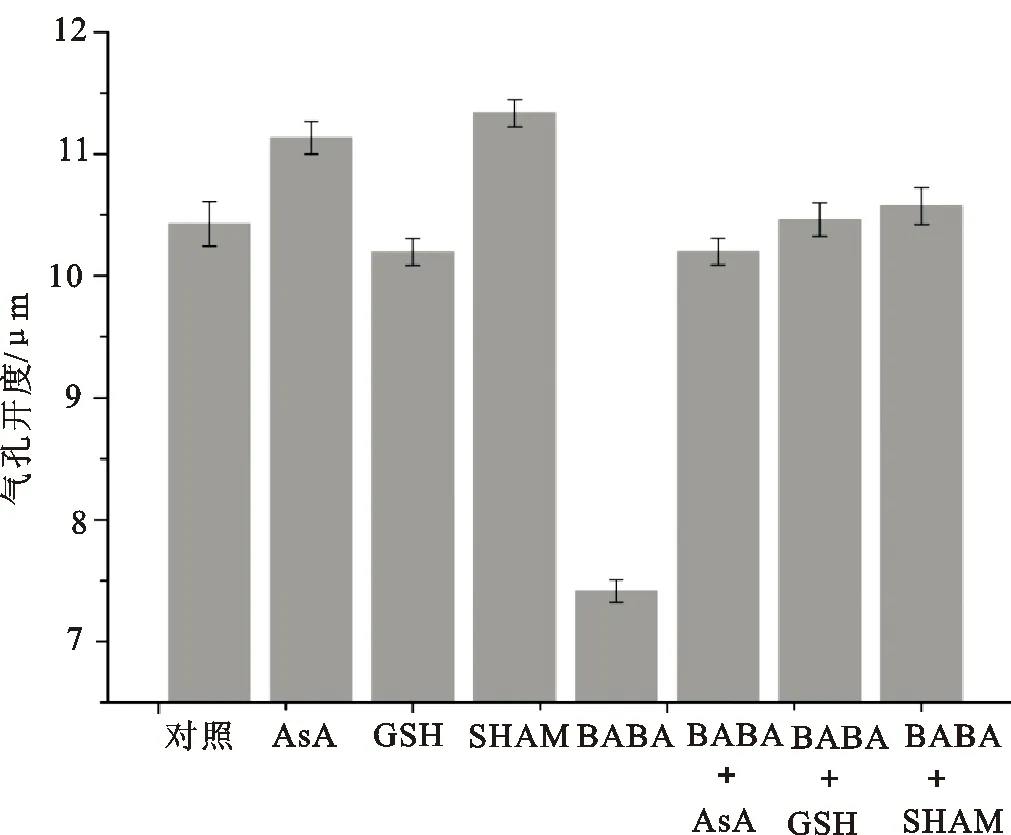
图2 活性氧参与BABA诱导的气孔关闭
2.2 ROS在BABA对吊竹梅叶片气孔开度影响中的作用
如图2所示,AsA、GSH和SHAM都逆转了BABA诱导的气孔关闭,经过AsA、GSH和SHAM处理后,BABA处理的表皮条气孔开度分别恢复到对照组的97.8%、94.0%和103.7%。
2.3 NO和Ca2+在BABA对吊竹梅叶片气孔开度影响中的作用
由图3和4所示,经过NO 清除剂(c-PTIO)、NR抑制剂(NaN3)和NOS抑制剂(L-NAME)预处理后,BABA导致的气孔关闭分别恢复到对照的99.5%、107.4%和106.3%。此外,经过钙离子螯合剂(EGTA)和钙离子拮抗剂(LaCl3)处理后,BABA处理的表皮条气孔开度分别恢复到对照组的100.3%和105.7%。

图3 NO参与BABA诱导的气孔关闭
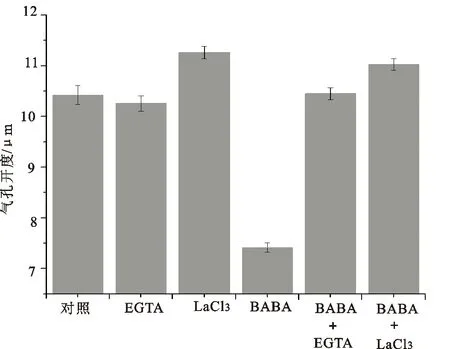
图4 Ca2+参与BABA诱导的气孔关闭
3 讨论
气孔是植物体上控制CO2的吸收和水分的散失的重要通道,其开闭运动受到诸多因素影响。许多信号物质在气孔开闭信号传递中发挥重要作用,包括产生H2O2的积累、肌动蛋白的重组、[Ca2+〗cyt的增加、pH值的改变等[2-4]。本论文的研究表明BABA可以诱导吊竹梅表皮气孔关闭。
笔者实验中的过AsA、SHAM和GSH完全抑制了BABA诱导的气孔关闭,这说明ROS参与了BABA诱导的气孔关闭,其中SHAM是细胞壁中的过氧化物酶抑制剂[6]。先前的研究表明,ROS (H2O2、O2-等)在气孔运动中发挥着重要作用。在笔者实验中,BABA诱导的气孔关闭被SHAM有效逆转,这说明细胞壁过氧化物酶可能参与了BABA诱发的吊竹梅保卫细胞ROS爆发过程。对SHAM敏感的过氧化物酶也是SA、UV-A和几丁质诱导的气孔关闭中H2O2的重要来源[3-7]。在保卫细胞中,ROS的生成与许多机制有关。因此,可以推测BABA诱导H2O2可能主要由细胞壁中的过氧化物酶产生。
现已证明,NO可以和ROS相互作用,通过调节细胞质膜Cl-通道、K+通道、Ca2+和MAPK的活性等控制气孔运动[7],本研究中,NOS抑制剂(L-NAME)、NO清除剂(c-PTIO)、NR抑制剂(NaN3)、钙离子螯合剂(EGTA)、钙离子拮抗剂(LaCl3)均扭转了BABA诱导的气孔关闭,推测NO和Ca2+介导了BABA诱导的气孔关闭。
4 结论
BABA可以诱导吊竹梅表皮气孔关闭,并且ROS、NO和Ca2+参与其信号传递过程,细胞壁中的过氧化物酶可能是H2O2产生的重要来源。
猜你喜欢
黑龙江大学自然科学学报(2022年4期)2022-11-17 08:07:52
河北科技师范学院学报(2022年2期)2022-08-26 08:55:46
干旱地区农业研究(2022年1期)2022-01-28 04:51:30
装备制造技术(2019年12期)2019-12-25 03:06:52
科技创新与品牌(2019年12期)2019-02-06 04:03:29
浙江工业大学学报(2017年5期)2018-01-22 02:03:42
广东农业科学(2017年5期)2017-08-29 10:37:54
中国塑料(2016年3期)2016-06-15 20:30:00
中国铸造装备与技术(2015年5期)2015-12-10 10:23:33
华东理工大学学报(自然科学版)(2015年4期)2015-12-01 04:00:36
