
低温超微粉碎对生物酶法制油豆渣蛋白结构影响的拉曼光谱分析
2019-05-05 06:56吴长玲寻崇荣刘宝华王中江江连洲
食品科学 2019年7期
吴长玲,寻崇荣,刘宝华,王中江,滕 飞,江连洲,李 杨*
(东北农业大学食品学院,黑龙江 哈尔滨 150030)
生物酶法制油作为一种新兴的“绿色、环保”提油技术,在提取油脂的同时能高效地回收油料中其他有价值组分,与传统工艺相比,其在能耗、环境保护和安全卫生等方面具有显著优势,且操作条件温和、工序简单[1-4]。现阶段国内外学者针对生物酶法制油技术的研究主要集中在预处理、酶解工艺条件、乳状液破除、油脂释放机制及油脂品质等方面,对于大豆加工过程副产物的研究主要集中在豆腐、豆浆残渣,而针对生物酶法制油豆渣的相关研究不多,对豆渣组分中蛋白结构、作用机理及潜在生物活性更是鲜有研究[5-8]。
针对目前豆渣利用率低且浪费严重的现状,许多学者对大豆加工副产物的深加工技术进行了研究,主要运用超微粉碎及螺杆挤压改性处理技术研究大豆豆渣粒度和加工性质,发现超微粉碎处理使豆渣具有一般颗粒所没有的特殊理化性质,如良好的溶解性、分散性、吸附性、化学反应活性等[9-11]。另外,有研究证实超微粉碎程度会促进大豆和原料中蛋白质的碱提取[12]。同时,Le Gall等[13]研究了粒度对豌豆粉蛋白质提取率的影响,发现随着粉碎程度的降低,面粉的平均粒度增加,而豌豆粉提取的蛋白质量分数从73.6%下降到37.4%。Russin等[14]研究发现分别经3 种不同研磨筛处理的脱脂豆粉,其蛋白提取率不同,说明粒度对蛋白回收有明显影响,这与Le Gall等[13]的研究结果相似。综上所述,超微粉碎对植物蛋白分离、回收及理化性质影响显著,与其他方法相比超微粉碎具有技术简单、成本低和可持续的优势,但超微粉碎的高速剪切作用使得体系温度升高,为避免高温对蛋白构象的影响,本研究采用低温超微粉碎处理生物酶法制油的豆渣。
拉曼光谱是一种基于拉曼散射和瑞利散射效应的光谱学分析技术,广泛应用于蛋白质空间结构的研究中,它可以通过鉴别蛋白质结构特征峰的变化,得到蛋白质分子多肽链骨架构型改变的信息和分子侧链微环境变化的信息[15]。拉曼光谱在不破坏样品的情况下可提供丰富的蛋白质结构信息,故本研究主要采用拉曼光谱深入解析豆渣蛋白结构。
本研究利用低温超微粉碎处理生物酶法制油豆渣,采用拉曼光谱分析豆渣蛋白经不同粉碎程度处理后豆渣蛋白的结构变化,并对其主要的碳链与侧链构象进行研究。旨在通过低温超微粉碎的处理方式高效回收豆渣蛋白,为生物酶法豆渣中蛋白的分离、纯化及回收提供理论指导和实践意义。
1 材料与方法
1.1 材料与试剂
大豆生物酶法制油豆渣由实验室自制;α-淀粉酶(DA4251,酶活力≥10 000 U/g) 合肥泊美生物科技有限责任公司;纤维素酶(酶活力≥10 000 U/g) 上海国药集团化学试剂有限公司;其他试剂均为分析纯。
1.2 仪器与设备
S22-2型恒温磁力搅拌器 上海司乐仪器有限公司;AL204型分析天平 梅特勒-托利多仪器(上海)有限公司;PHS-3C雷磁pH计 上海精科仪器有限公司;SWFJ超微粉碎机 广州市旭朗机械设备有限公司;TDL-408台式离心机 上海安亭科学仪器厂;HH-4数显搅拌水浴锅 常州赛普实验仪器厂;鼓风干燥箱上海一恒科学仪器有限公司;Raman Station 400激光显微拉曼光谱仪 美国PE公司。
1.3 方法
1.3.1 酶法制油豆渣的制备
对Li Yang等[16]的方法进行适当修改,将市售大豆粉碎过60 目筛,取200 g过筛后的粉体,按料液比1∶6加入蒸馏水,待搅拌均匀后放入55 ℃水浴锅内进行酶解,酶解条件为:酶解温度55 ℃、酶解时间2 h、pH 9.0、碱性蛋白酶Protex6L(8 900 U/mL)添加量0.5%(体积分数),边搅拌边酶解,酶解结束后取出并用1 mol/L HCl溶液调节水溶液pH值至7,之后在100 ℃沸水中灭酶5 min,将灭酶后的溶液于4 500 r/min离心20 min,豆渣在平板铺平后置于55 ℃鼓风烘箱,待恒质量后取出研磨即得生物酶法制油豆渣。
1.3.2 低温超微粉碎处理豆渣的制备
由于豆渣中的脂肪氧化后会产生异味并影响提取率[17],因此在低温超微粉碎前按照杨梦曦等[18]的处理方法除去脂肪。然后,采用超微粉碎机在-4 ℃下超微粉碎酶法制油豆渣(水分质量分数≤4%),粉碎时间30 s,出料粒度分别为100、200、300 目,然后烘干至质量恒定,即得所需豆渣样品。
1.3.3 未处理及常温超微粉碎处理豆渣的制备
未处理组即不经超微粉碎处理,其他步骤与1.3.2节一致。常温超微粉碎处理组于常温(25 ℃)下操作,出料粒度为200 目,其他步骤同1.3.2节。
1.3.4 豆渣蛋白的提取
根据Petruccelli等[19]的方法提取渣蛋白。取150 g豆粉,用正己烷以料液比1∶6混合脱脂3 次,得到脱脂豆粕,将脱脂豆粕按1∶10的质量比与水混合,用2 mol/L NaOH溶液调节pH值至8.5,45 ℃搅拌2 h后,将其悬浮液在4 ℃条件下10 000×g离心20 min,取上清液再用2 mol/L HCl溶液调节pH值至4.5。静置后在4 ℃条件下6 000×g离心20 min,得蛋白沉淀后水洗2 次,最后取沉淀分散于水中并用2 mol/L NaOH溶液调节pH值至7.0。再在4 ℃条件下10 000×g离心30 min,除去少量的不溶物,将此蛋白溶液冷冻干燥后研磨即得粉末状大豆分离蛋白。
1.3.5 超微粉碎处理前后豆渣蛋白的拉曼光谱分析
采用Raman Station 400激光显微拉曼光谱仪。激发光波长为785 nm,激光功率为80 mW,扫描范围400~2 000 cm-1,每次扫描时间60 s,积分10 次,4 次扫描进行累加。以苯丙氨酸((1 003±1)cm-1)作为归一化因子,采用ACD Labs V12软件进行谱图基线校正、谱峰归属查找,最终进行豆渣蛋白的拉曼光谱分析。
1.4 数据统计及分析
所有的实验至少进行3 次,利用SPSS Statistics 22软件对数据进行差异显著性分析(方差分析),P<0.05为差异显著。分别采用Origin 8.5、PeakFit 4.12软件进行数据分析、图表处理及图谱分析。
2 结果与分析
2.1 豆渣蛋白主链结构特征拉曼光谱分析
为了有效分析经低温超微粉碎处理生物酶法制油豆渣中蛋白组分结构特征,本研究采用拉曼光谱进行分析研究,结果如图1所示。
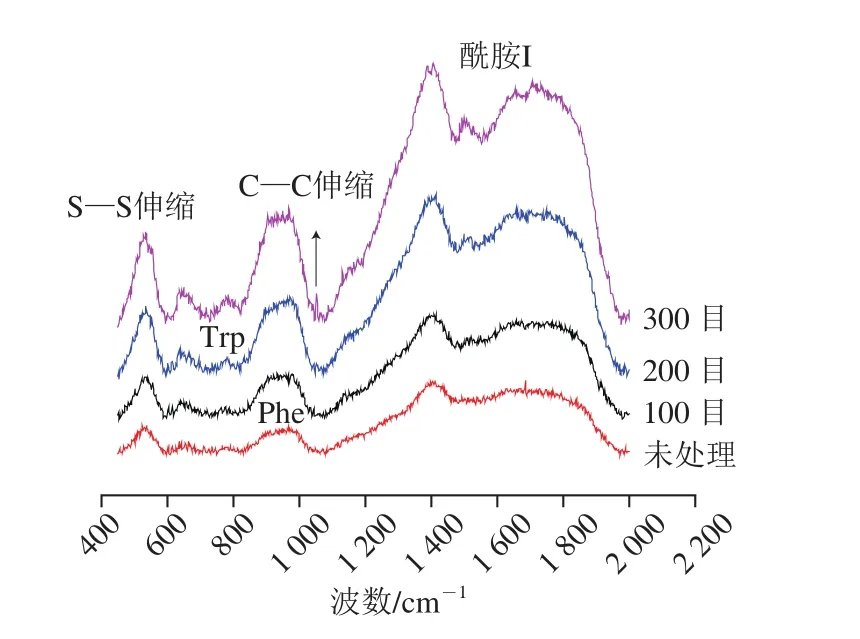
图1 不同粉碎条件下豆渣蛋白拉曼光谱Fig. 1 Raman spectra of soybean meal protein ground to different particle sizes

表1 拉曼光谱峰位归属Table 1 Attribution of Raman spectral peaks
参考拉曼光谱峰位归属(表1),豆渣蛋白酰胺I带拉曼特征峰位置为:α-螺旋结构归属峰位(1 645~1 660 cm-1);β-折叠结构(1 670~1 680 cm-1);β-转角结构(1 680~1 690 cm-1);无规卷曲结构(1 660~1 670 cm-1)。本研究中拉曼图谱二级结构含量的定量计算由PeakFit 4.12软件分析完成,图2所示为豆渣蛋白拉曼谱带在1 600~1 700 cm-1区域内的拟合图谱。
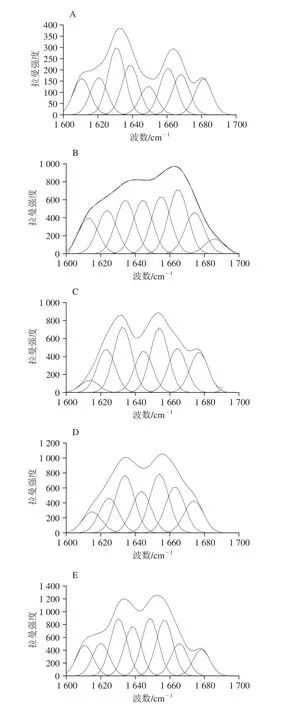
图2 豆渣蛋白拉曼谱带在1 600~1 700 cm-1区域内的拟合图谱Fig. 2 Raman fitting curves of soybean meal protein in the range of 1 600-1 700 cm-1

表2 超微粉碎改性处理豆渣蛋白二级结构组分含量Table 2 Percentages of protein secondary structures in soybean meal subjected to different treatments
将豆渣蛋白的酰胺I带用于研究豆渣蛋白二级结构分析,结果如表2所示。经低温超微粉碎后豆渣蛋白中β-转角及无规卷曲结构含量显著下降(P<0.05),常温超微粉碎后豆渣蛋白无规卷曲结构含量降至12.54%,β-转角结构含量显著增加(P<0.05)。随着低温超微粉碎程度的不断加剧,豆渣蛋白α-螺旋及β-折叠含量分别呈现先增加后下降、先下降后上升的变化趋势。经低温超微粉碎处理与常温处理豆渣蛋白二级结构组分均以α-螺旋、β-折叠为主。另外发现与未处理豆渣蛋白相比,常温超微粉碎处理后豆渣蛋白无规卷曲结构含量较低温超微粉碎下降趋势更为明显。
与未处理条件对比,超微粉碎的高剪切作用使蛋白分子间的相互作用逐渐减弱,其疏水结合及离子结合可能因体积的缩小被切断,蛋白质分子伸展,非极性基团暴露,分子间疏水作用增强,氢键作用逐渐减弱,导致豆渣蛋白α-螺旋与β-折叠结构的加和增加,β-转角及无规卷曲结构含量下降,这与Rahmeh等[20]通过荧光光谱分析分子间偶联程度对G蛋白表面疏水作用影响的研究结果一致。另外,蛋白质二级结构随蛋白质聚合程度而变化,较高程度的蛋白质聚合与稳定的二级结构的积聚高度相关,α-螺旋和β-折叠构象比例高,表明超微粉碎处理后,豆渣蛋白分子间有序结构增加,蛋白质聚合增强。由于粉碎过程中蛋白粒径减小,比表面积大幅增加,豆渣蛋白聚合程度增强,从而导致豆渣蛋白这种结构单元变化的发生,这与Xiong Licheng[21]、Hou[22]和Belton[23]等的研究结果一致。同时,Xiong Licheng[21]和Lee[24]等的研究表明,精磨粉碎处理后蛋白质二级结构β-折叠结构含量的增加和β-转角结构含量的减少,表明形成了更多的聚合和稳定的谷蛋白网络,豆渣蛋白二级结构更趋有序化,印证了本研究中α-螺旋结构含量增加、β-转角及无规卷曲结构含量下降的趋势。
与低温超微粉碎条件相比,常温粉碎在高速剪切作用过程中对豆渣蛋白存在温度效应,处理过程中使部分豆渣蛋白变性,蛋白结构发生解折叠,蛋白分子内疏水性氨基酸暴露,导致分子间作用力下降,故豆渣蛋白β-折叠含量总体增加不明显,β-转角结构增加明显,说明常温粉碎处理下,部分无规卷曲结构转变成β-转角结构。这可能是由于超微粉碎处理温度能先让蛋白解聚,使蛋白分子充分展开,分子内埋游离巯基暴露,而暴露的游离巯基在常温粉碎温度下被氧化形成分子间或分子内二硫键,蛋白分子重新聚合形成新的聚集体造成蛋白三级结构的改变,进而引起二级结构单元含量发生相应变化[25]。
随着低温超微粉碎目数的增加(100~200 目),超微粉碎的这种高剪切作用不断加强,导致豆渣蛋白分子间碰撞作用增强,分子间的相互作用逐渐减弱,氢键作用逐渐减弱,故豆渣蛋白α-螺旋含量与β-折叠含量比值增加,无规卷曲含量呈上升趋势;当粉碎程度达到200 目时,豆渣蛋白分子间碰撞作用停止,粉碎程度继续增加,氢键作用反而增大,分子间作用增强,故出现豆渣蛋白α-螺旋含量与β-折叠含量比值下降现象。
2.2 豆渣蛋白侧链结构特征拉曼光谱分析
已有研究表明蛋白质结构改变能够引起色氨酸残基的暴露,760 cm-1附近区域的拉曼峰强度降低与色氨酸残基由原本“包埋态”转变为“暴露态”有关,在拉曼谱图中表现为色氨酸谱带强度的降低[26-29]。由表3可以看出,与未经超微粉碎处理的豆渣蛋白相比,低温超微粉碎处理引起760 cm-1附近区域的拉曼光谱峰强度降低,表明色氨酸残基趋向于“暴露态”。而较未处理豆渣蛋白,常温超微粉碎处理引起760 cm-1附近区域的拉曼光谱峰强度升高,表明色氨酸残基趋向于“包埋态”。色氨酸残基的暴露是由于超微粉碎处理使蛋白质结构改变,分子结构展开导致的;而常温超微粉碎处理下色氨酸残基的包埋可能与超微粉碎温度导致豆渣蛋白部分变性有关[30]。在本研究中,经低温超微粉碎(200 目)处理的豆渣蛋白在760 cm-1附近区域的色氨酸拉曼光谱峰强度较低,该区域归属于色氨酸残基的伸缩振动,表明此条件下色氨酸残基更趋于“暴露态”,这与上述不同粉碎程度下分子碰撞作用影响有关。
Herrero等[29]研究指出850 cm-1和830 cm-1是酪氨酸残基苯环的呼吸振动和面外弯曲倍频之间的费米共振。如表3所示,I850/I830比值分布于0.99~1.09之间,表明所测豆渣蛋白的酪氨酸残基暴露于溶液的极性微环境下作为中性强度氢键的供体或受体。与未经超微粉碎处理的豆渣蛋白相比,低温超微粉碎处理时酪氨酸费米共振线(I850/I830)比值较小,表明酪氨酸残基微环境在低温超微粉碎过程中得以保持。然而,在常温超微粉碎处理条件下,I850/I830比值降低到0.99,较低温超微粉碎下降更为明显,这表明超微粉碎处理温度会导致酪氨酸残基暴露程度加剧。由此可以推测经超微粉碎处理后豆渣蛋白的酪氨酸应更趋于“暴露态”,且粉碎程度过大会导致酪氨酸“暴露”程度降低。这表明超微粉碎破坏了维持蛋白质三级结构的主要作用力,使包埋于豆渣蛋白分子内部的酪氨酸残基暴露于分子表面,并作为氢键的供体或受体与水相互作用[31]。但随着粉碎程度加大,分子间碰撞作用停止,分子间氢键作用增强,导致疏水性氨基酸酪氨酸残基暴露程度降低。
另外,Ngarize[32]和Ferrer[33]等的研究表明1 450 cm-1是脂肪族氨基酸的拉曼归属谱线,脂肪族氨基酸的拉曼归属峰强度降低与脂肪族氨基酸暴露有关。在本研究中,经超微粉碎作用下的豆渣蛋白在1 450 cm-1处的拉曼归属峰强度总体低于未处理方式,这与豆渣蛋白在超微粉碎过程中的结构重排有关。另发现常温超微粉碎较低温超微粉碎I1450/I1003比值下降更明显,表明常温超微粉碎的高剪切作用会导致部分豆渣蛋白变性,从而影响蛋白质三级结构,与上述2.1节分析结果一致。

表3 不同处理条件下豆渣蛋白侧链基团谱带强度Table 3 Band intensities of side chain groups in soybean meal protein subjected to different treatments
2.3 豆渣蛋白二硫键拉曼光谱分析
de Vuyst[34]和王中江[35]等的研究表明在拉曼谱图中500~550 cm-1范围内是二硫键的特征谱带,二硫键在不同振动模式下所反映出的拉曼光谱位移有所不同,其中,500~515 cm-1处为gauche-gauche-gauche(g-g-g)模式,515~525 cm-1处为gauche-gauche-trans(g-g-t)模式,525~545 cm-1处为trans-gauche-trans(t-g-t)模式。为了探究超温粉碎改性对豆渣中豆渣蛋白二硫键的影响,本研究运用Peak Analyzer软件进行多峰值Guassina拟合处理。
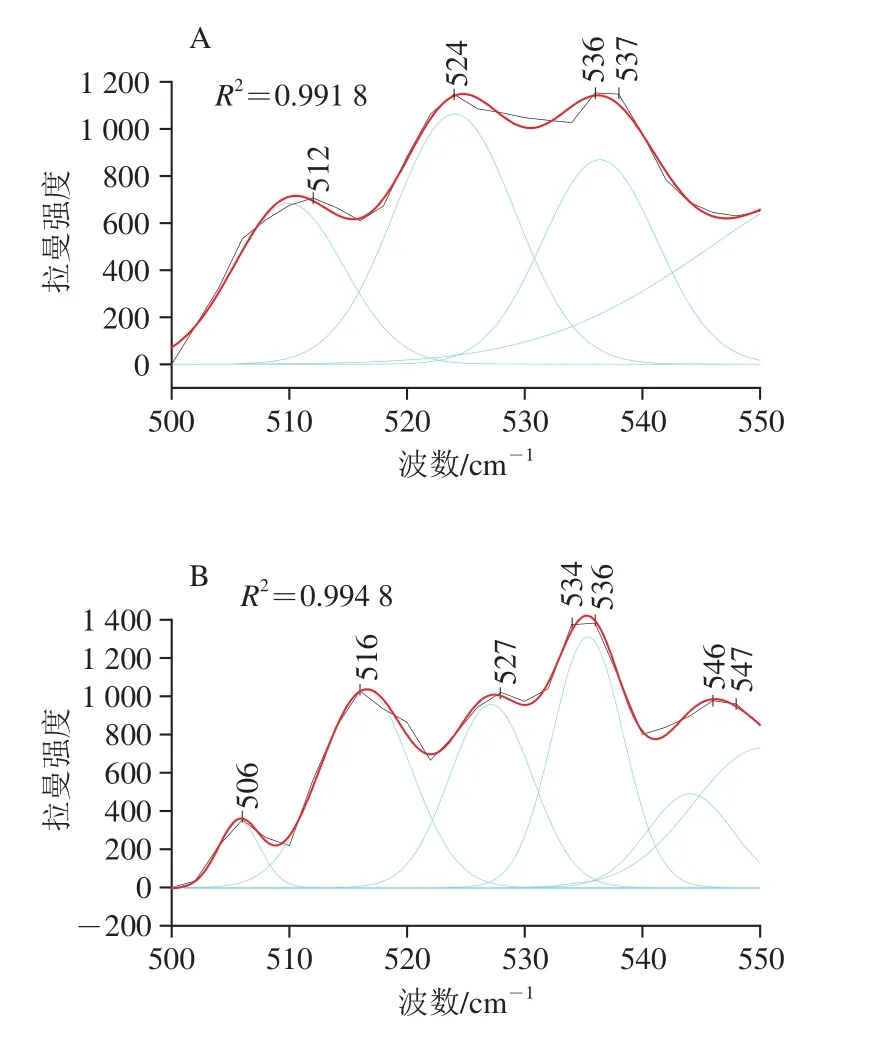
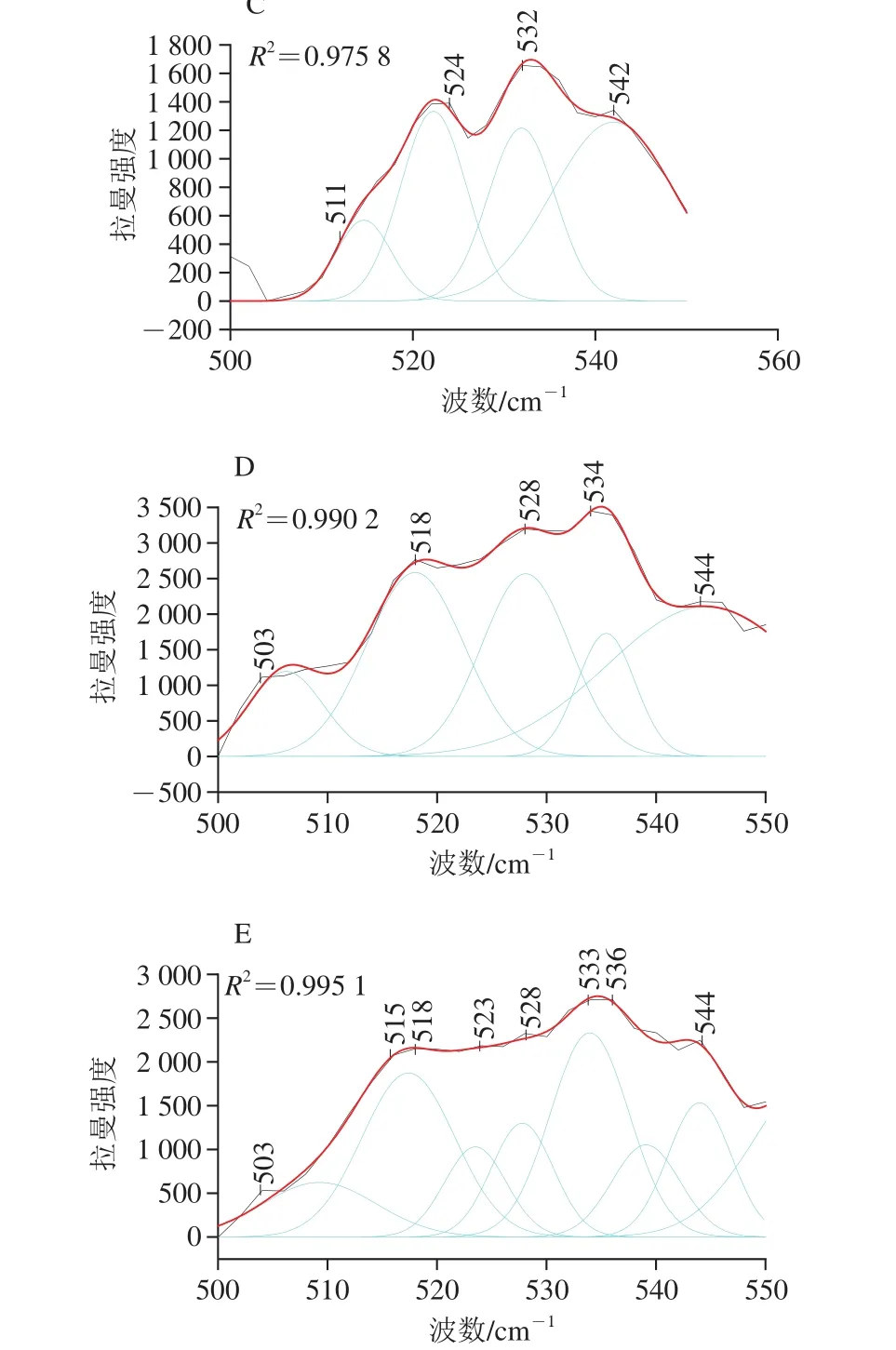
图3 豆渣蛋白拉曼谱带在500~550 cm-1区域内的高斯拟合图Fig. 3 Raman Gaussianves fitting curves of soybean meal protein in the region of 500-550 cm-1
从图3可以看出,拟合谱线重新合成的谱线与实验数据贴近,证明拟合结果是准确的。对拟合的各个高斯峰值进行归属,各处理方式豆渣蛋白的二硫键的振动模式归属情况如表4所示。本研究运用各个峰的高度计算各振动模式的百分比,以及二硫键的数量。结合表4及图3的分析,经超微粉碎处理后g-g-g与g-g-t模式百分比下降显著,t-g-t模式百分比显著增加,且随着物料粉碎程度的加剧,二硫键数量变化较为明显,这可能是由于超微粉碎处理破坏大的蛋白颗粒的同时,使得埋藏在内部的游离巯基暴露,从而被氧化形成二硫键,这与Chan等[36]的研究结果一致。另外,常温超微粉碎过程中剪切力产生的温度效应会引起豆渣蛋白结构解折叠现象,分子间作用力下降,导致分子内二硫键数量下降,故常温超微粉碎处理下二硫键中g-g-g模式含量显著下降。随着粉碎目数的增加,豆渣蛋白t-g-t模式含量分别呈现先下降后增加的变化趋势,说明超微粉碎高速剪切处理将大颗粒蛋白聚集体破坏,形成新的小颗粒的过程中也有部分游离巯基暴露氧化成二硫键,造成蛋白三级结构的改变;同时,t-g-t模式含量的增加表明超微粉碎程度的加深使豆渣蛋白分子间作用力增强。可以得到结论:超微粉碎作用能够显著改变豆渣中豆渣蛋白的二硫键构型,由g-g-g构型向t-g-t构型转变,这种变化表现出一种非线性变化趋势,经超微粉碎处理后t-g-t构型成为豆渣蛋白二硫键的主要构型。共价二硫键和非共价相互作用(如氢键、离子键和疏水键)有助于形成稳定豆渣蛋白结构,这与Domenek等[37]的研究结果相似。
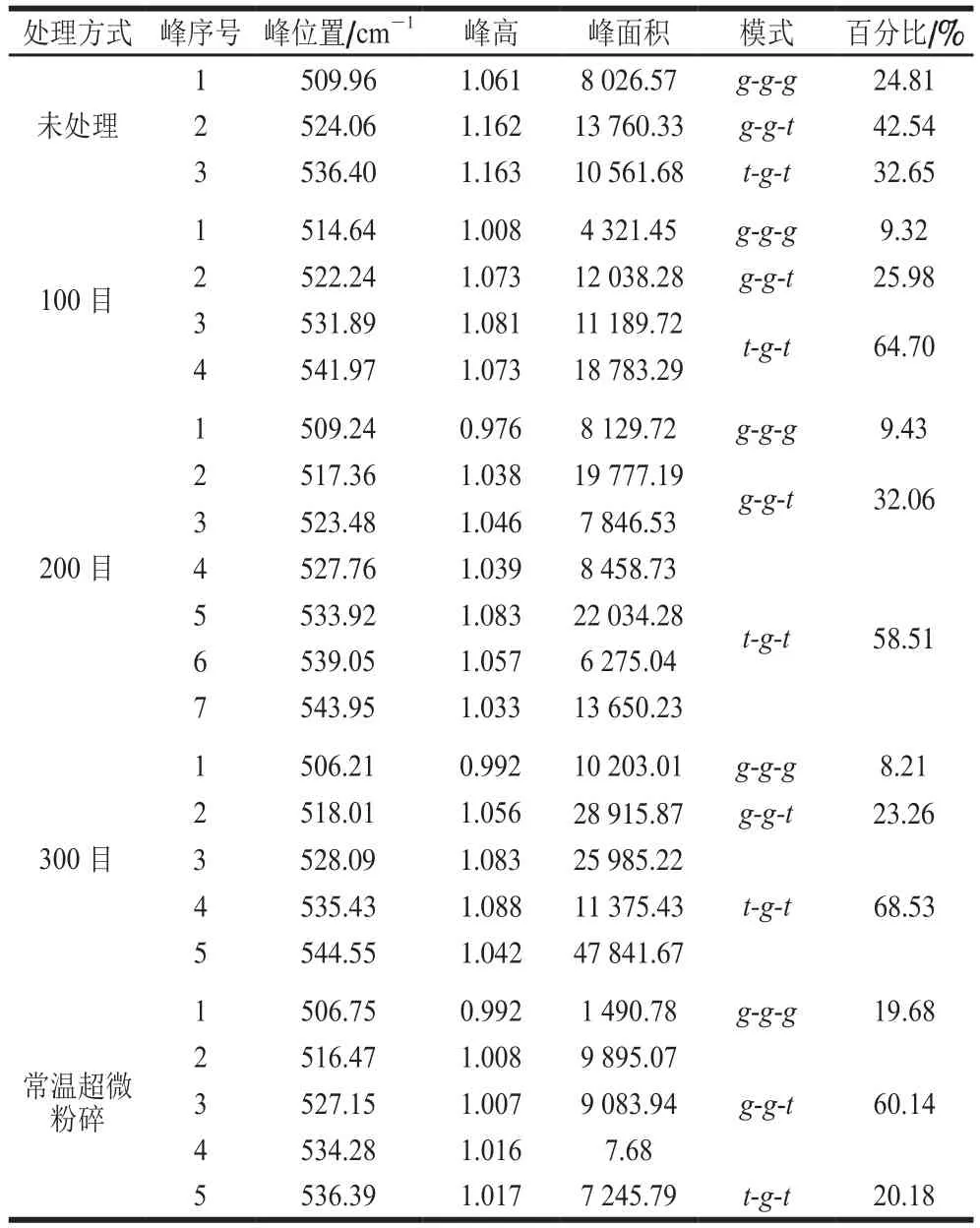
表4 豆渣蛋白在500~550 cm-1区域拟合结果Table 4 Fitting results of soybean meal protein in the region of 500-550 cm-1
3 结 论
生物酶法制油豆渣经低温超微粉碎处理后,豆渣蛋白改性前后二级结构发生显著变化,处理后豆渣蛋白中无序二级结构单元含量明显降低,其中α-螺旋结构含量增加,改性后β-转角结构含量降低至几近为零;通过I760/I1003、I850/I830及I1450/I1003拉曼强度比值发现经超微粉碎改性处理的豆渣中,豆渣蛋白的疏水性基团暴露程度高于未超微粉碎处理方式;超微粉碎处理改变了豆渣中蛋白的二硫键构型,经超微粉碎处理后t-g-t模式含量显著增加,呈现出了一种非线性变化趋势;对比低温、常温超微粉碎发现,在-4 ℃超微粉碎条件下豆渣蛋白二级结构变化更趋明显。
猜你喜欢
食品科学技术学报(2022年5期)2022-10-11
生物化学与生物物理进展(2022年7期)2022-07-25
生物化学与生物物理进展(2022年6期)2022-07-21
中学生数理化(高中版.高考理化)(2021年2期)2021-03-19
粮食科技与经济(2020年2期)2020-05-09
祝您健康·文摘版(2019年12期)2019-12-13
分析化学(2019年4期)2019-05-13
中国饲料(2016年17期)2016-12-01
凤凰生活(2016年3期)2016-03-08
中国老年学杂志(2015年5期)2015-12-31
