
生境破碎化对苦豆子种子害虫种群数量及危害率的影响
2019-04-29 09:28:30张大治张圣昕马志芳吴光胜
生态科学 2019年2期
张大治, 张圣昕, 马志芳, 吴光胜
生境破碎化对苦豆子种子害虫种群数量及危害率的影响
张大治*, 张圣昕, 马志芳, 吴光胜
宁夏大学生命科学学院, 银川 750021
苦豆子是一种典型的防风固沙先锋植物, 其生长分布呈典型的斑块化格局。于2017年6月至9月在宁夏灵武市宁东镇刘家寨地区选取了LJZ2017A(01—08斑块)和LJZ2017B(01—06斑块)两个样地共14个苦豆子植物斑块, 调查了各斑块内苦豆子种子害虫(豆荚螟)的种群数量及其危害率。结果表明: 在LJZ2017A样地斑块面积平均为1457.79 m2, 种子害虫数量平均为0.69只每荚, 密度为0.65只·m-2; 在LJZ2017B样地斑块面积平均为949.87 m2, 种子害虫数量平均1.46只每荚, 密度为1.30只·m-2。种子害虫对豆粒的危害率在LJZ2017B样地为49.91%, 明显高于LJZ2017A样地的24.50%。种子害虫密度与斑块面积的逐步回归分析显示随着斑块面积的增大而呈显著下降趋势(=16.58,= 0.0015,=0.7617), 对豆粒的危害率随着生境破碎化程度增加呈增加趋势(=7.45,=0.0183,=0.6189)。
苦豆子; 生境破碎化; 豆荚螟; 危害率; 斑块面积
0 前言
生境破碎化使生物多样性降低的主要原因之一[1–2]。生境破碎化使斑块数量增加, 斑块面积减少, 生境斑块隔离度增加, 影响物种的繁殖和迁移能力[2–3], 不同的物种对生境破碎化都有其独特的反应, 这种反应在很大程度上取决于物种自身的生物学特性[4]。苦豆子()为多年生豆科草本植物, 主要分布于中国西北的荒漠、半荒漠地区。宁夏苦豆子主要分布在盐池、红寺堡、同心、灵武、中宁、中卫等县区的荒漠半荒漠区域[5], 其野外种群的生长呈现典型的斑块化格局。由于其较高的药用价值和生态功能, 苦豆子资源的合理保护与开发利用越来越引起人们的重视, 宁夏回族自治区已将苦豆子列入重点保护的六大地道药材之一, 纳入国家中药现代化科技产业行动计划中[6]。种子繁殖是苦豆子的一种重要繁殖方式, 近些年来, 由于生态环境的变化、落地种子不易获得适宜生长发育的条件及种子害虫的危害是造成其种群衰落、稀有的重要原因, 是苦豆子种群发展的主要瓶颈。豆荚螟()是苦豆子种子的主要害虫[7], 在苦豆子呈斑块化的分布格局下, 斑块面积与种子害虫的危害率之间存在着怎样的相关性?破碎化的生境斑块格局对苦豆子种子害虫的发生是否存在着影响?对该问题的探讨, 了解害虫发生的规律与危害特点, 以期为该类害虫的防治、合理开发利用苦豆子资源提供理论依据。
1 材料与方法
1.1 研究区域概况
本次实验的采样地设在宁夏灵武市宁东镇南的刘家寨, 该地域属沙漠低山丘陵地貌, 大部分地段属固定半固定沙地, 沙生植物苦豆子种群自然生长, 呈大小不一的典型斑块化分布格局。选择了两个样地(LJZ2017A, LJZ2017B)共14个斑块, 其中LJZ2017A有8个斑块, 编号为LJZ2017A01—LJZ2017A08, 地理坐标为38°05’20N—38°05’28N, 106°36’47—106° 36’50E, 海拔1224 m—1238 m。LJZ2017B样地有6个斑块, 编号为LJZ2017B01—LJZ2017B06, 地理坐标为38°05’7N—38°05’12N, 106°36’31—106°36’34E, 海拔1236 m—1244 m。
1.2 野外调查及室内统计饲养方法
于2017年6月至9月期间, 在苦豆子豆荚螟进入老熟幼虫阶段前进行抽样调查, 每隔10天1次, 共调查3次。对选定的14个斑块按照五点采样法, 在每个斑块内设置5个样点, 每样点再选取5个1 m× 1 m的小样方, 每小样方内取2株苦豆子(用剪刀剪取其整株豆荚), 每样点共10株苦豆子; 小样点间距离根据斑块面积大小不同而有差异, 1 m—5 m不等。每斑块共采集50株苦豆子。将采集到的每株豆荚装入塑料袋, 做好采集记录标签, 供室内统计用。同时测量样地内苦豆子的高度、盖度等, 记录斑块生境情况。
室内对采摘的苦豆子豆荚在体视显微镜下剥检, 统计每枚豆荚中的种子害虫数、豆荚蛀孔数(为采摘后荚内豆荚螟幼虫咬破豆荚荚皮逃逸或转荚所留的孔)、荚内豆粒总数、被害豆粒数等。每个斑块共统计豆荚750枚。将从荚内剥离的苦豆子种子装入饲养袋, 其余豆荚放入饲养盒中, 继续观察记录种子害虫及天敌情况。
1.3 斑块特征指数计算
斑块特征指数计算按照以下公式[8–11]:
(1)斑块密度指数():=/, 式中是某一景观的斑块个数,为景观斑块的面积。值越大表示破碎化程度越高。
(4)种子害虫的平均危害率=受害豆粒数/总豆粒数×100%。
利用EXCEL2007、DPS(v16.05)[12]统计软件对所得数据进行分析处理, 相关性采用Pearson指数。
2 结果与分析
2.1 不同斑块内苦豆子种子害虫密度及其危害率
根据田间调查数据, 两个样地14个斑块面积、斑块中苦豆子种子害虫的种群密度、对豆荚及豆粒的危害率等见表1。
由表1可以看出, LJZ2017A样地斑块面积平均为1457.79 m2, LJZ2017B样地斑块面积平均为949.87 m2, 苦豆子盖度LJZ2017A样地(61.75%)与LJZ2017B样地(65.60%)差异不显著。苦豆子豆荚的每荚平均豆粒数LJZ2017A样地(4.75个每荚)与LJZ2017B样地(4.15个每荚)差异不大, 反应出这两个样地苦豆子的生长情况基本一致。
种子害虫豆荚螟的数量在LJZ2017A样地为0.22—1.38只每荚之间, 平均0.69只每荚, 密度为0.65只·m-2; LJZ2017B样地豆荚螟的数量在0.93—2.11只每荚之间, 平均1.46只每荚, 密度为1.30只·m-2, 约是LJZ2017A样地的2倍(图1)。
种子害虫的危害率在LJZ2017A样地为9.87%—47.12%, 平均为 24.50%; LJZ2017B样地为27.45%—66.42%, 平均为49.91%。危害率LJZ2017B样地明显高于LJZ2017A样地(图2)。
2.2 斑块破碎化与苦豆子种子害虫危害率的关系
2.2.1 苦豆子斑块特征指数
LJZ2017A样地和LJZ2017B样地的斑块特征指数与种子害虫平均危害率情况见表2。LJZ 2017A样地斑块平均面积稍大于LJZ2017B样地, 样地内各斑块最小距离指数(斑块间的中心距离)LJZ2017A样地稍大于LJZ2017B样地。从生境的破碎化程度看, LJZ2017B样地内斑块破碎化程度稍高于LJZ2017A样地, 种子害虫的平均危害率LJZ2017B样地高于LJZ2017A样地, 表明随着斑块破碎化程度的增加, 种子害虫的危害率也呈增加的趋势。
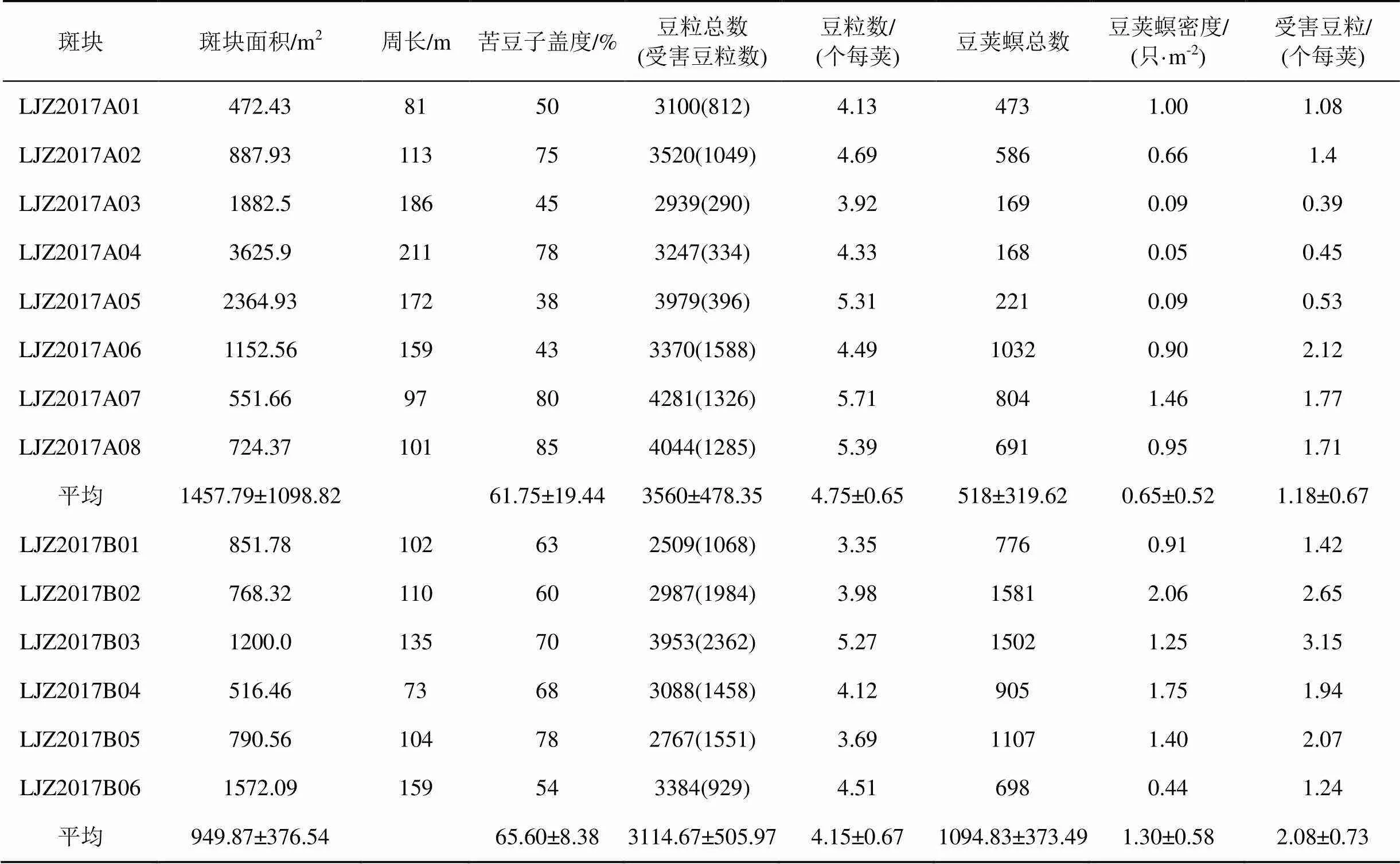
表1 斑块特征指数及豆荚螟危害情况(平均值±标准差)
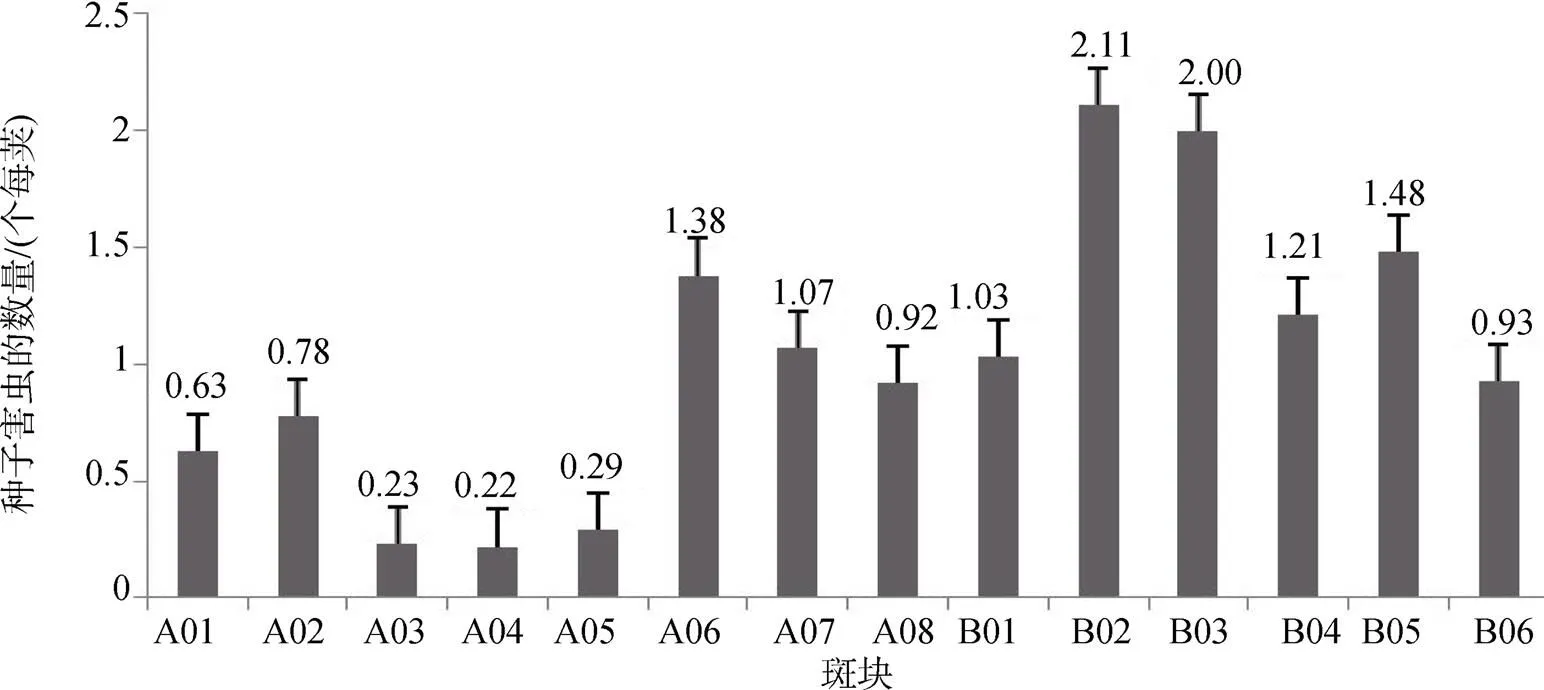
图1 不同斑块中种子害虫的数量
Figure 1 Individuals of seed pest in a pot in different patches
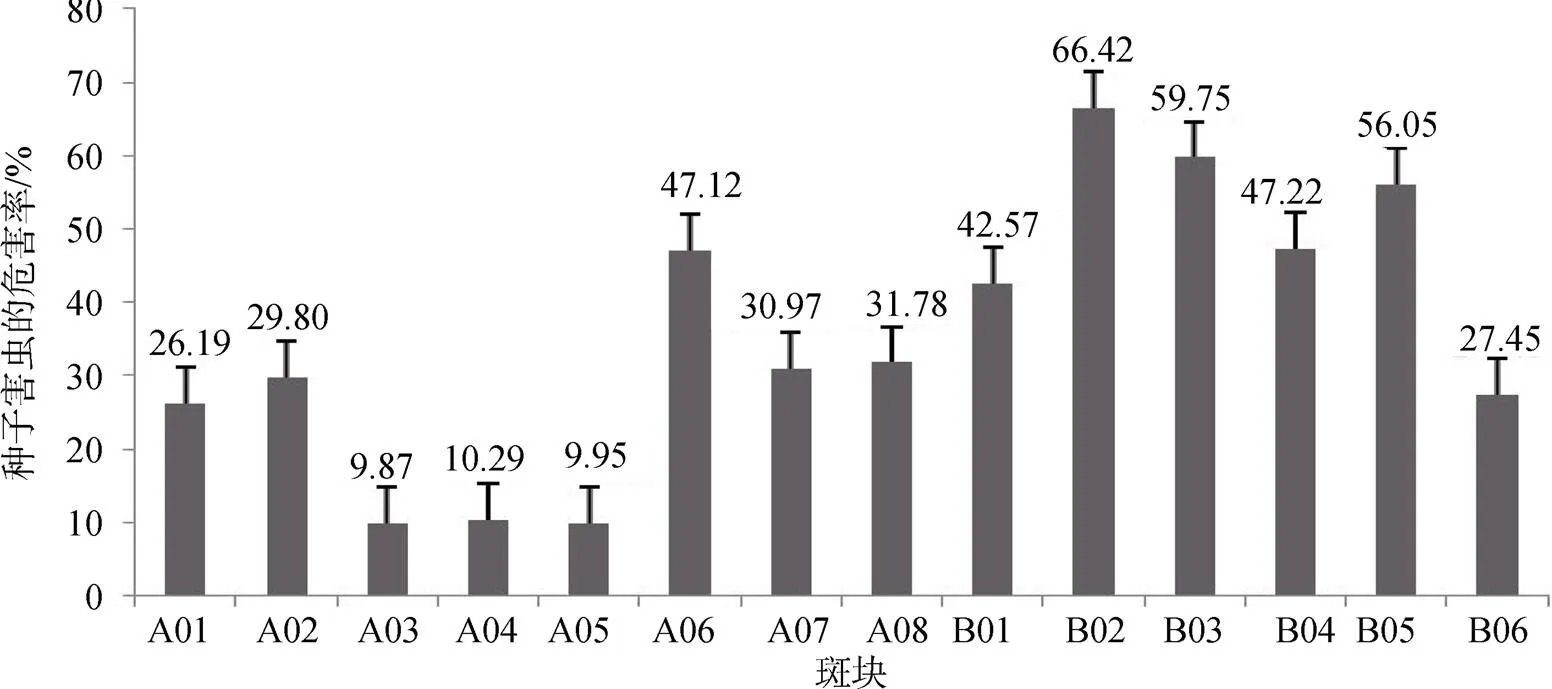
图2 不同斑块中苦豆子种子害虫的危害率
Figure 2 Damage rate of seed pest in different patches
2.2.2 景观斑块面积与豆荚螟密度、危害率的关系
通过对斑块面积与豆荚螟密度进行逐步回归分析, 得到回归方程为:=1.59973-0.0005404(=16.58,=0.0015,=0.7617), 表明斑块内豆荚螟的密度与斑块面积存在显著的相关性, 且为典型的负相关, 即在同等抽样量下, 豆荚螟的数量伴随着斑块面积的增加而数量呈下降趋势。
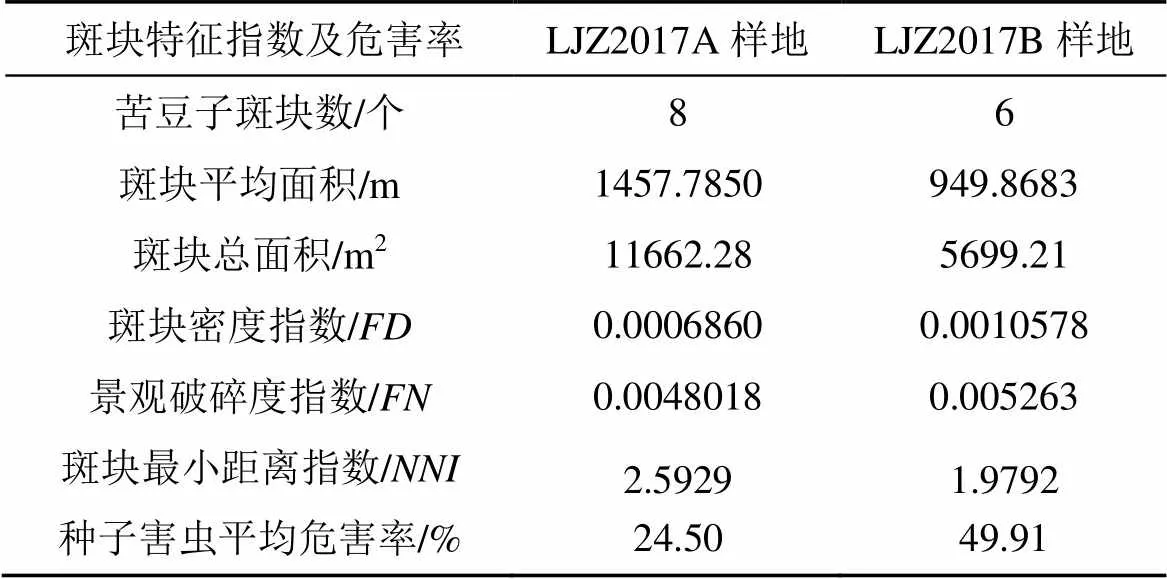
表2 两个样地的景观破碎化指数及种子害虫的危害率
对豆荚螟的危害率与斑块面积进行逐步回归分析, 得到回归方程为:=51.54987-0.01303(=7.45,=0.0183,=0.6189), 表明豆荚螟的危害率与斑块面积存在显著的负相关, 小斑块的苦豆子, 其种子害虫的危害率相对较为严重, 生境的破碎化增加了种子害虫的危害率。
3 讨论
生境面积和破碎化被公认为对种群的丰富度和持久性有重要的影响, 每一个物种对生境破碎化都有其独特的反应, 这种反应在很大程度上取决于物种自身的生物学特性和该物种对生境的依赖程度[13]; 在斑块面积下降时, 景观内资源的互补性可能对某些物种种群密度会产生积极或中立的影响[14]。由于豆荚螟是生境内柠条、苦豆子、沙冬青等豆科植物种子的共有害虫, 幼虫具有转荚习性, 成虫具有一定的迁飞能力。通过本实验反应了苦豆子豆荚螟种群数量的发生与苦豆子斑块的面积、破碎化程度存在着一定的相关关系, 但是这种相关可能还受到其他景观要素的影响, 景观廊道、豆荚螟寄主植物资源的互补性等可能也有一定的影响, 因为在LJZ2017A样地, 斑块周边多为小路或沟渠, 形成典型的景观廊道, 在一定程度上限制了周边环境中豆荚螟的迁入; 而在LJZ2017B样地, 苦豆子斑块周边存在着其他的荒漠沙生植物, 特别是零星散布着的柠条、甘草等其他豆科植物, 且斑块间的距离较近, 有利于周边环境及斑块间豆荚螟的迁移。当然, 在一个局部生境破碎化的景观基质中, 景观内豆荚螟可利用资源的互补性、斑块面积及格局及豆荚螟的扩散能力对其种群的发生依赖程度等方面对还有待于进一步的研究。
本项目试验区由于治沙工程的实施, 沙生植物形成了“生境岛屿”, “生境岛屿”的面积影响着植物的多样性及其分布格局[15], 同样也影响着岛屿内植食性害虫及其天敌的多样性和格局。最小生存面积是评价景观斑块格局与结构对害虫和天敌影响力的重要手段之一, 也是探素利用景观因素达到进行害虫生物防治的重要途径之一。空间变小、连通性降低、空间及集合群落过度破碎化等都不利于猎物—捕食者系统的稳定和持续[16]。本实验获得苦豆子种子害虫的一些寄生性天敌, 这些天敌对豆荚螟种群可能起到了一定控制的作用, 因为通过对斑块面积与种子害虫的危害率、天敌的数量回归分析显示, 苦豆子植物斑块的破碎化程度增加、斑块面积减小, 种子害虫天敌的数量减少了, 而种子害虫的危害率增加了。至于天敌对苦豆子种子害虫的控害率、猎物—捕食者系统的稳定和持续性在多大斑块面积上发挥作用, 需要进一步探讨。
[1] POGSON B. Habitat fragmentation reduces biodiversity[J]. Science, 2015, 347(6228): 1325–1325.
[2] FISCHER J, LINDENMAYER D B. Landscape modification and habitat fragmentation: a synthesis[J]. Global Ecology and Biogeography, 2007, 16: 265–280.
[3] 武晶, 刘志民. 生境破碎化对生物多样性的影响研究综述[J]. 生态学杂志, 2014, 33(7): 1–7.
[4] 杨芳, 贺达汉. 生境破碎化对生物多样性的影响[J]. 生态科学, 2006, 25(6): 564–567.
[5] 尹长安. 干旱荒漠半荒漠地区苦豆子的资源状况及开发利用[J]. 干旱区资源与环境, 1995, 9(2): 48–54.
[6] 史伟, 陈志国. 苦豆子的开发与利用[J]. 草业与畜牧, 2007, 134(1): 57–59.
[7] 郭晶静, 张大治, 杨婷婷, 等. 荒漠风沙区苦豆子种子害虫的空间分布型[J]. 江苏农业科学, 2012, 40(10): 111–113.
[8] 余新晓, 牛健植, 关文彬, 等. 景观生态学[M]. 北京: 高等教育出版社, 2006: 280–317.
[9] 刘惠清, 许嘉巍. 景观生态学[M]. 长春: 东北师范大学出版社, 2008: 270–292.
[10] 邬建国. 景观生态学——格局、过程、尺度与等级(第二版)[M]. 北京: 高等教育出版社, 2007: 1–230.
[11] 王宪礼, 布仁仓, 胡远满, 等. 辽河三角洲湿地的景观破碎化分析[J]. 应用生态学报, 1996,7(3): 299–304.
[12] TANG Qiyi, ZHANG Chuanxi. Data Processing System (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research[J]. Insect Science,2013, 20(2): 254–260.
[13] ANDREN H. Habitat fragmentation, the random sample hypothesis and critical thresholds[J]. Oikos, 1999, 84: 306–308.
[14] SUMMERVILLE KS, CRIST TO. Contrasting effects of habitat quantity and quality on moth communities in fragmented landscapes[J]. Ecography, 2004, 27: 3–12.
[15] 武晶, 余新春, 刘志民, 等. 科尔沁沙地“生境岛屿”面积对植物物种多样性及其分布格局的影响[J]. 生态学杂志, 2017, 36(1): 43–53.
[16] 董红娟, 吴新卫, 洪娇, 等. 空间结构对捕食关系影响研究进展[J]. 应用生态学报 , 2017, 28(2) : 712–720.
Effects of habitat fragmentation on seed pest populations and damage rate of
ZHANG Dazhi*, ZHANG Shengxin, MA Zhifang, WU Guangsheng
School of Life Science, Ningxia University, Yinchuan 750021, China
mainly distributes in the desert regions of Northwest China as a highly adversity-resistant perennial officinal plant. It is a windbreak and fixation sand pioneer plant. A total of 14 patches offrom LJZ2017A (01-08 patches) and LJZ2017B (01-06 patches) were selected.was seed pest of.The damage rate, density and individual ofwere investigated from June to September 2017 in Liujiazhai region of Ningdong Town, Lingwu City, Ningxia. The results showed that in LJZ2017A plot, the average area of patches was 1457.79 m2, the average number of seed pests per pod was 0.69, and the average density was 0.65 individual per square meter. In LJZ2017B plot, the average area of patches was 949.87 m2, the average number of seed pests per pod was 1.46, and the average density was 1.30 individual per square meter. The seed damage rate was 49.91% in LJZ2017B plot, which was significantly higher than 24.50% in LJZ2017A plot. The stepwise regression analysis of seed pest density and patch area showed that the density of seed pests decreased significantly with increasing patch area (= 16.58,= 0.0015,= 0.7617), and the seed damage rate increased with increasing degree of habitat fragmentation (= 7.45,= 0.0183,= 0.6189).
;habitat fragmentation;; damage rate; patch area
10.14108/j.cnki.1008-8873.2019.02.005
Q968
A
1008-8873(2019)02-031-05
2018-03-11;
2018-06-05
国家自然科学基金项目(31560611, 31860619)
张大治(1970—), 男, 宁夏西吉人, 博士, 教授, 主要从事动物生态与动物资源研究, E-mail: zdz313@nxu.edu.cn
张大治
张大治, 张圣昕, 马志芳, 等. 生境破碎化对苦豆子种子害虫种群数量及危害率的影响[J]. 生态科学, 2019, 38(2): 31-35.
ZHANG Dazhi, ZHANG Shengxin, MA Zhifang, et al. Effects of habitat fragmentation on seed pest populations and damage rate of[J]. Ecological Science, 2019, 38(2): 31-35.
猜你喜欢
儿童时代·幸福宝宝(2022年12期)2022-12-09 11:23:50
小学生优秀作文(低年级)(2021年11期)2021-12-06 05:21:56
落叶果树(2021年6期)2021-02-12 01:29:10
作文周刊·小学一年级版(2018年8期)2018-03-15 15:01:57
学生天地(2017年12期)2017-05-17 05:51:16
体育科技(2016年2期)2016-02-28 17:06:07
娃娃画报(2015年11期)2015-12-08 04:39:23
故事作文·低年级(2015年10期)2015-11-27 23:06:44
文理导航·趣味课堂(2015年4期)2015-08-18 19:38:48
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:17
