
艾芬地尔对单眼形觉剥夺小鼠初级视皮层兴奋性递质受体表达的影响△
2019-04-18 01:22:56宋德胜钱晶陈志钧
眼科新进展 2019年4期
宋德胜 钱晶 陈志钧
作者单位:210029 江苏省南京市,南京医科大学附属儿童医院眼科
弱视为视觉发育相关性疾病,是视觉发育期由于单眼斜视、屈光参差、形觉剥夺或双眼高度屈光不正等异常视觉经验,造成单眼或双眼最佳矫正视力低于正常,或双眼视力相差2行以上,临床检查无可见的器质性病变,经恰当治疗后视力可以提高或完全恢复[1]。形觉剥夺为弱视的成因之一,多见于先天性白内障、角膜混浊、完全性上睑下垂、医源性眼睑缝合或遮盖等情况。由于形觉刺激不足,剥夺了黄斑形成清晰物像的机会而形成弱视[1],而且视觉发育关键期单侧的形觉剥夺会导致视觉环路众多位置发生可塑性变化。视觉可塑性也已成为大脑可塑性的重要组成部分,视觉可塑性的研究不仅能为弱视的治疗提供光明的前景,更能丰富大脑神经可塑性的内容。目前关于视觉可塑性的研究包括长时程增强(long-term potentiation,LTP)和长时程抑制(long-term depression,LTD),稳态突触缩放(homeostasis synaptic scaling),脉冲时间依赖的突触可塑性(spike timing dependent plasticity,STDP)[2-5],且发生机制均与N-甲基-D-天门冬氨酸(N-methy-D-aspartate,NMDA)的生化代谢相关,NMDA受体是哺乳动物中枢神经系统兴奋性神经递质谷氨酸的3种离子型受体之一。NMDA受体在视觉转导过程、神经细胞生长及突触可塑性变化中发挥重要作用,并与视觉发育有密切的关系,它在弱视形成中的作用及其作用机制一直是眼科医生及视觉科学家们研究的热点。NMDA受体第二亚单位 (NMDA-NR2)是NMDA受体复合物的调节亚单位,通过多种方式调节受体复合物的功能活性,不同NR2的参与可赋予通道复合物不同的电生理学和药理学特性[6]。而NR2中NR2A亚基与NR2B亚基又因其本身的功能特性及其与突触可塑性与再可塑性的密切关系成为目前神经科学研究的一大热点,有学者发现NR2A亚基与NR2B亚基发育性表达与视觉发育关键期的开启与结束有关[7-9],并且外界视觉经验对两种亚基的转换至关重要,出生后早期,NR2B亚基占优势,发育过程中,逐渐被NR2A亚基取代[10]。Guo等[11]研究结果发现,暗饲养能延长两种亚基转换的时间,使功能性NR2B亚基型NMDA受体在突触部位增加,其为暗饲养小鼠STDP整合时间窗拓宽(可塑性增强)的内在机制,并且研究发现NR2B亚基特异性受体阻断剂艾芬地尔能使暗饲养对小鼠视皮层NR2B亚基/NR2A亚基相互转换时间造成的影响恢复正常。
单眼形觉剥夺可导致弱视,对视觉可塑性的影响较暗饲养更为严重,而关于单眼形觉剥夺后视皮层NR2A亚基和NR2B亚基表达及NR2B亚基阻断剂对其表达影响的研究未检索到相关文献,这就限制了此种形式可塑性研究的发展。本研究以单眼形觉剥夺小鼠为实验模型,从蛋白和mRNA两个途径观察双侧视皮层NR2A亚基和NR2B亚基的表达并探索NR2B亚基受体阻断剂艾芬地尔对两种亚基表达的影响。
1 材料与方法
1.1材料出生后3~4周C57BL/6J小鼠(野生型)15只,雌雄不限,无特定病原体动物,12 h12 h昼夜循环饲养(光亮时间在0700至1900)。饲养温度为23 ℃。实验动物由中国医学科学院生物医学工程研究所提供。实验动物的使用遵循国家科学技术委员会颁布的《实验动物管理条例》。
1.2方法
1.2.1实验动物分组将健康小鼠随机分3组,每组5只,分组均以出生后24 d为基准:正常对照组、单眼形觉剥夺组和联合组。单眼形觉剥夺组和联合组右眼褥式缝合6 d,建立单眼形觉剥夺模型。
1.2.2实验模型制备单眼形觉剥夺模型制备:单眼形觉剥夺组和联合组小鼠均选择右眼进行干预,于出生后24 d,实验小鼠称质量后,七氟烷麻醉,剪除右眼睑缘周围毛发,碘伏消毒上下眼睑局部皮肤,距上下睑缘各1.0 mm处,剪去皮肤和睑板组织,用6-0丝线间断缝合实验眼3~4针。术后每天涂抗生素眼膏及检查创口情况,避免感染造成的缝线脱落。这两组小鼠均共同在自然光线环境下饲养,饲养至生后30 d。联合组小鼠称质量后,于造模后第1、3、5天腹腔注射艾芬地尔,给药剂量为10 mg·kg-1。
1.2.3qRT-PCR法检测视皮层NR2A亚基和NR2B亚基mRNA的表达NR2A上游引物:5’-ACGTGACAGAACGCGAACTT-3’,下游引物:5’-TCAGTGCGGTTCATCAATAACG-3’;NR2B上游引物:5’-GCCATGAACGAGACTGACCC-3’,下游引物:5’-GCTTCCTGGTCCGTGTCATC-3’;GAPDH上游引物:5’-AGGTCGGTGTGAACGGATTTG-3’,下游引物:5’-GGGGTCGTTGATGGCAACA-3’;引物序列来自PrimerBank,由北京赛百盛基因技术公司合成。采用Trizol法提取组织mRNA,紫外分光光度计测量RNA纯度并计算RNA浓度,参照SYBR Green试剂盒说明书将RNA逆转录合成cDNA,以cDNA为模板进行扩增。扩增条件:95 ℃预变性15 s;95 ℃变性 15 s,57 ℃退火30 s,74 ℃延伸30 s,共40个循环。以GAPDH为内参照,采用2-ΔΔCT法计算NR2A mRNA和NR2B mRNA的相对表达量。
1.2.4Westernblot法检测视皮层NR2A亚基和NR2B亚基蛋白的表达采用细胞膜蛋白与细胞浆蛋白抽提试剂盒提取带有NR2A亚基和NR2B亚基的膜蛋白,BCA法测定蛋白质量浓度,取新管做好标记,加入适量膜蛋白,相对应蛋白中加入对应的5倍上样缓冲液,涡旋振荡后短暂离心。用96 ℃金属浴加热10 min后迅速离心,快速置于冰上,-20 ℃保存待用。聚丙烯酰胺凝胶电泳,采用湿转法转印至PVDF膜;5 g·L-1脱脂奶粉中封闭1 h,分别加入相应一抗,摇床4 ℃孵育过夜;TBST缓冲溶液漂洗5次,每次10 min,加入缓冲溶液稀释的二抗室温下摇床缓慢孵育1 h,TBST缓冲溶液漂洗5次,每次10 min,ECL化学发光法显色,X线显影。使用Image Pro-Plus软件测量各蛋白条带的吸光度(absorbance,A)值。目的蛋白的相对灰度=A目的蛋白/A相应内参。
2 结果
2.1qRT-PCR法检测结果
2.1.1各组内左右侧视皮层NR2AmRNA、NR2BmRNA相对表达量比较正常对照组左右侧视皮层NR2A mRNA、NR2B mRNA相对表达量差异无统计学意义(P=0.454、0.484)。单眼形觉剥夺组与联合组左侧视皮层,即剥夺眼对侧视皮层,NR2A mRNA相对表达量较右侧视皮层减少(均为P=0.000);单眼形觉剥夺组左侧视皮层NR2B mRNA表达较右侧视皮层增多(P=0.000),联合组左右侧视皮层NR2B mRNA相对表达量差异无统计学意义(P=0.431;见表1和表2)。
2.1.2各组间左侧视皮层NR2AmRNA、NR2BmRNA相对表达量比较与正常对照组相比,单眼形觉剥夺组左侧视皮层NR2A mRNA相对表达量明显减少(F=117.332,P=0.000),NR2B mRNA相对表达量明显增加(F=55.506,P=0.000);与单眼形觉剥夺组相比,联合组左侧视皮层NR2A mRNA相对表达量差异无统计学意义(F=10.661,P=0.061),NR2B mRNA相对表达量明显减少(F=173.044,P=0.000;见表1和表2)。
2.1.3各组间右侧视皮层NR2AmRNA、NR2BmRNA相对表达量比较与正常对照组相比,单眼形觉剥夺组右侧视皮层NR2A mRNA相对表达量明显增加(F=8.591,P=0.026),NR2B mRNA相对表达量明显减少(F=515.238,P=0.000);与单眼形觉剥夺组相比,联合组右侧视皮层NR2A mRNA相对表达量差异无统计学意义(F=0.072,P=0.798)、右侧视皮层NR2B mRNA相对表达量增加(F=37.308,P=0.001;见表1和表2)。
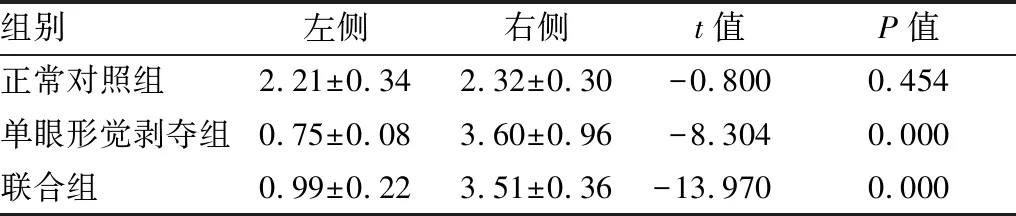
表1 各组小鼠视皮层NR2A mRNA相对表达量
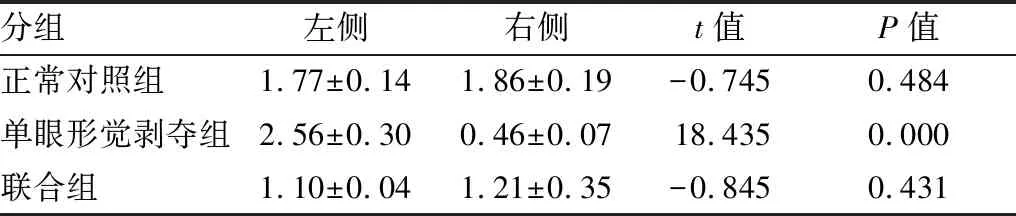
表2 各组小鼠视皮层NR2B mRNA相对表达量
2.2Westernblot法检测视皮层NR2A亚基和NR2B亚基的表达在相对分子质量220 000与 105 000 之间可见目的条带NR2A亚基、NR2B亚基。在相对分子质量71 000与50 000之间可见内参条带β-tubulin。
2.2.1各组内左右侧视皮层NR2A蛋白、NR2B蛋白相对表达量比较正常对照组左右侧视皮层NR2A蛋白、NR2B蛋白相对表达量差异无统计学意义(P=0.890、0.606),单眼形觉剥夺组与联合组左侧视皮层,即剥夺眼对侧视皮层NR2A蛋白相对表达量均较右侧视皮层减少(P=0.014、0.013);单眼形觉剥夺组左侧视皮层NR2B蛋白表达较右侧视皮层增多(P=0.023),联合组左右侧视皮层NR2B蛋白相对表达量差异无统计学意义(P=0.234;见表3和表4)。
2.2.2各组间左侧视皮层NR2A蛋白、NR2B蛋白相对表达量比较与正常对照组相比,单眼形觉剥夺组左侧视皮层NR2A蛋白相对表达量明显减少(F=42.829,P=0.003),NR2B蛋白相对表达量明显增加(F=15.027,P=0.018);与单眼形觉剥夺组相比,联合组左侧视皮层NR2A蛋白相对表达量差异无统计学意义(F=2.117,P=0.219),NR2B蛋白相对表达量减少(F=15.984,P=0.016;见表3和表4)。
2.2.3各组间右侧视皮层NR2A蛋白、NR2B蛋白相对表达量比较与正常对照组相比,单眼形觉剥夺组右侧视皮层NR2A蛋白相对表达量增加(F=8.514,P=0.043),NR2B蛋白相对表达量减少(F=8.514,P=0.043);与单眼形觉剥夺组相比,联合组右侧视皮层NR2A蛋白相对表达量差异无统计学意义(F=3.603,P=0.131),右侧视皮层NR2B蛋白相对表达量增加(F=10.816,P=0.025;见表3和表4)。

表3 各组小鼠视皮层NR2A 蛋白相对表达量
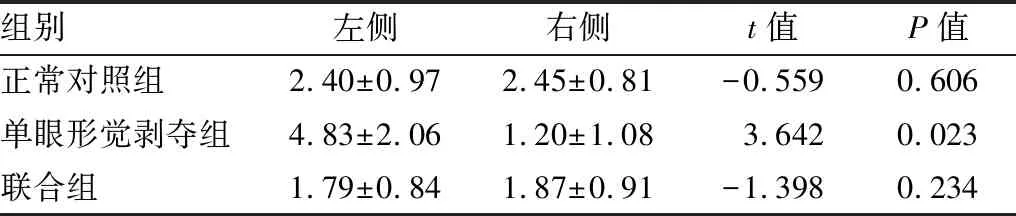
表4 各组小鼠视皮层NR2B 蛋白相对表达量
3 讨论
单眼形觉剥夺是异常视觉经验的重要形式之一。众多研究表明,视觉发育关键期单侧的形觉剥夺会导致视觉环路中众多位置发生可塑性变化。鼠类90%~95%的视纤维在视交叉中交叉到对侧[12],因此这种影响具有半脑选择性,也就是主要影响被剥夺眼的对侧半脑,故单眼形觉剥夺对被剥夺眼对侧视皮层所产生的影响远大于剥夺眼同侧视皮层。小鼠出生后3~5周为视觉发育关键期[13],在此期间进行单眼形觉剥夺已成为一个成熟的弱视模型,自从20世纪60年代创建此模型以来,就成为了视觉发育及可塑性研究的典型模型。
本研究从分子遗传学及蛋白组学水平发现:(1)关键期内单眼形觉剥夺6 d剥夺眼对侧视皮层中NR2A mRNA及蛋白表达减少,NR2B mRNA及蛋白表达增多;(2)剥夺眼同侧视皮层中,NR2A mRNA及蛋白表达增加,NR2B mRNA及蛋白表达减少;(3)NR2B受体阻断剂艾芬地尔能显著降低NR2B受体的表达,并阻断单眼形觉剥夺对NR2B表达的影响,但对NR2A表达无影响。
NR2A和NR2B型NMDA受体在视皮层中占绝大部分,出生后早期,NR2B亚基占优势,发育过程中,逐渐被NR2A亚基取代[14-16]。外界视觉经验对两种亚基的转换至关重要,异常的视觉刺激能使NR2A/NR2B比值朝着错误的方向发展,如暗饲养能延长两种亚基转换的时间[14,17-18],视觉经验通过不同的调控水平调节NR2A/NR2B比值。有一种观点认为,NR2A亚基的发育性表达增加是视觉经验在转录水平起作用的结果,而NR2B亚基则是在翻译水平缓慢下降[19],同时MIb-2泛素化也可增强NR2B亚基降解[20]。剥夺双眼视觉经验后,NR2A mRNA合成减慢,NR2B翻译抑制消除,NR2B降解减弱。因此,视觉剥夺可降低NR2A水平,同时增加NR2B水平,进而NR2A/NR2B值出现明显下降[21]。本研究显示,关键期内单眼形觉剥夺6 d剥夺,眼对侧视皮层中NR2A mRNA及蛋白表达减少,NR2B mRNA及蛋白表达增多,NR2A/NR2B比值降低,转录与翻译结果一致,证明视觉经验可能在转录水平抑制了NR2B的翻译,而剥夺眼同侧视皮层中NR2A mRNA及蛋白表达增加,NR2B mRNA及蛋白表达减少,NR2A/NR2B比值升高,表明剥夺眼同侧视皮层NR2A/NR2B比值不仅会竞争性增加,而且比正常情况下更加明显(F=117.332,P=0.000),单眼形觉剥夺可能会导致剥夺眼同侧视皮层可塑性关键期缩短。尚需电生理学方法来证实此假设。
艾芬地尔作为选择性作用于NMDA受体NR2B亚型的阻滞剂受到越来越多的关注,其在治疗神经病理性疼痛、药物依赖、脑卒中、帕金森病中具有很大的潜力[22-24]。随着NR2A亚基与NR2B亚基研究的深入及其在突触可塑性中发挥的重要作用,艾芬地尔作为特异的NR2B亚基阻断剂在视觉可塑性方面的应用越来越广泛。艾芬地尔能使暗饲养对小鼠视皮层NR2B亚基/NR2A亚基相互转换时间造成的影响恢复正常,阻断NR2B亚基介导的兴奋性突触后电流,并且这种阻断作用明显较单眼形觉剥夺组增强,艾芬地尔能使暗饲养拓宽的脉冲时间依赖的突触可塑性时间窗恢复正常,进而解释了STDP整合时间窗拓展的机制[11]。但是关于NR2B亚基受体阻断剂对受体亚基具体表达的影响至今仍未见报道。我们的研究表明,艾芬地尔能显著降低剥夺眼对侧视皮层NR2B mRNA和蛋白的表达量,并且降低作用要比单眼形觉剥夺组明显,而剥夺眼同侧视皮层NR2B mRNA和蛋白反而有所增高,但差异并无统计学意义,可能是艾芬地尔使剥夺眼对侧视皮层NR2B亚基表达减少,消除两侧视皮层之间的竞争抑制的结果。而艾芬地尔对NR2A亚基的表达并没有影响,也说明NMDA受体NR2A亚基和NR2B亚基的总量并不是不变的,两者的表达可能有不同的调节路径。
另外,本实验的缺陷在于:(1)没有对两种亚基的表达进行连续性的检测,进而绘制发育关键期NR2A亚基和NR2B亚基表达曲线;(2)视皮层定位采用小鼠脑解剖图谱,并参照特定的解剖标志,未能在高倍显微镜下通过神经细胞结构等确定视皮层边界,取材难免存在误差。
猜你喜欢
中华养生保健(2022年7期)2022-04-13 13:03:18
昆明医科大学学报(2022年1期)2022-02-28 07:43:34
昆明医科大学学报(2021年8期)2021-08-13 08:59:20
昆明医科大学学报(2021年6期)2021-07-31 07:40:08
世界最新医学信息文摘(2020年68期)2020-12-25 11:55:27
医学与法学(2020年3期)2020-09-18 09:21:48
吉林大学学报(医学版)(2015年4期)2015-12-17 07:48:13
西南医科大学学报(2015年1期)2015-08-22 13:01:48
中国社区医师(2015年12期)2015-01-27 11:19:31
下一代英才(2013年5期)2013-04-29 19:47:37
