
外源一氧化氮对镉胁迫酸枣幼苗生理代谢指标的影响
2019-04-16 02:47:54麻云霞李钢铁梁田雨王檬檬童春元田梦妮
西南林业大学学报 2019年2期
麻云霞 李钢铁 梁田雨 王檬檬 邹 苗 童春元 杨 颖 田梦妮
( 内蒙古农业大学沙漠治理学院,内蒙古 呼和浩特 010018)
镉(Cd)是拥有较强毒性的重金属元素和土地污染物之一,最近几年,由于各行业生产水平的高速提升,工业废水、废渣排入农田及不合理施肥制度的使用,导致我国众多地区土壤污染日趋严重,Cd2+产生积聚,其含量超过一般土壤,于植株的生长有负面效应[1-2]。有研究证明逆境下植物细胞超微结构会发生变化,矿质营养元素的吸收速率降低,出现活性氧堆积现象,抑制了植物生长,Cd2+是非还原型重金属,不能参加哈伯-韦斯反应,因此也会影响植物保护酶系统,包括植物呼吸作用酶、光合作用酶及根系脱氢酶和固氮酶等[3-4]。Cd2+在植物生长发育过程中所起的作用呈现双面性,浓度较小时,可促进植物体内某些生理指标的积累,即为学术界认可的施加较小浓度Cd2+对植物能发挥显著的“积极效应”,而浓度较高时则产生负面的胁迫作用,且土壤中Cd2+形态易被植物吸收,从地下部分转运到地上部分,通过食物链等途径传递,最终对动物和人类的健康造成不可估量的危害[5]。
酸枣(Zizyphus jujuba)属鼠李科(Rhamnaceae)枣属(Zizyphus),广布于华北、西北、东北和华东的向阳山坡、荒芜丘陵和平原,从东部的湿润地带到西部的干旱区均有分布。酸枣根系发达,具有抗旱、耐盐碱、耐低温的特性,在水土保持方面有重要的作用,是生态恢复的先锋树种。但目前对该植物的研究主要集中于生物活性物质提取[6]、药用价值开发[7]、组培嫁接[8]、抗旱能力及结构研究[9]等方面。重金属污染研究主要集中在经济作物和农作物上,对园林观赏植物的研究较为少见[10]。酸枣作为枣树的砧木时多在土壤和灌溉条件好的苗圃地,酸枣繁殖过程中幼苗极易受到Cd2+胁迫危害的情况并不多见,但在新建枣园和新疆实生播种建园时确有因Cd2+胁迫出现严重出苗不齐、幼龄树死亡的情况,且Cd2+为环境中存在剧毒的重金属污染物。一氧化氮(NO)作为气体小分子信号物质,在植物体内参与多种生理过程,如种子萌发[11]、调节植物光合作用[12]、侧根生长[13]、植物抗逆性[14]、细胞程序性死亡等[15]。本试验以酸枣为试材,以重金属Cd2+为目标污染物,通过研究不同浓度的硝普钠(SNP)对Cd2+胁迫下酸枣幼苗生长、活性氧代谢、抗氧化酶系,矿质营养元素的摄入及光系统等的影响,筛选出SNP缓解Cd2+胁迫的最佳浓度,初步探讨外施NO应用于酸枣幼苗对Cd胁迫下的调控作用,这将为预防Cd2+造成的植物污染,降低Cd2+在酸枣体内的积累提供依据。
1 材料与方法
1.1 试验材料
本试验在内蒙古农业大学科教示范区(111°73′29″E,40°83′35″N,海拔 1051 m)开展,供试材料为酸枣,种子由巴彦淖尔市林业科学研究所提供。选取大小均匀、圆润光滑的酸枣种子使用0.1%的HgCl2进行10 min杀菌,蒸馏水冲刷3~5遍,放于湿润的纱布上25℃避光催芽,36 h后将出芽的种子播于洗净的蛭石营养钵中,用1/4 Hoagland营养液定时定量灌溉,当幼苗出现3~4片真叶后,选取长势均一、生长较好的幼苗清除完根部残留物,换成完全营养液继续培育,每3 d更换1次营养液。水培期间电动气泵保持持续通气状态,幼苗长出5~6片真叶之后,开始进行Cd2+胁迫试验。
1.2 试验设计
2017年6月15日,对长势恢复较好的幼苗正式开始预试验。将不添加Cd2+作为对照(CK),试验设计梯度为 BT1(25 μmol/L Cd2+);BT2(50 μmol/L Cd2+);BT3(75 μmol/L Cd2+);BT4(100 μmol/L Cd2+);BT5(125 μmol/L Cd2+)。试验 30d 后根据幼苗生物量和相对生长速率(RGR)筛选出50 μmol/L CdCl2为 Cd2+胁迫浓度。
SNP除了产生NO外,还会生成NO2-/NO3-和Fe(CN)52-等。因此,设定 100 μmol/L的 NaNOx和Na3Fe(CN)6处理作为试验相关处理。根据预实验得出Cd2+胁迫浓度及所添加NO供体SNP特性,外源物质对酸枣幼苗在Cd2+胁迫下的生理生态效应试验共设置以下7个处理,试验设计方案如表1所示。

表1 外源物质对镉胁迫酸枣的生理生态效应试验设计方案Table 1 Experimental design scheme of physiological and ecological effects of exogenous materials on Z. jujube under Cd2+ stress
每组处理共设置3个重复,随机分布在大棚中,营养液培养期间其营养液更换频率为每天1次,使用KOH或HCl将pH调整为6.6±0.2。温室里光周期是15 h,白天其最高温度为31.3 ℃,夜间最低温度为14.4 ℃,在试验第20天时,取样进行分析测定。NO供体硝普钠购自德国Merck公司,先用蒸馏水配制100 mmol/L的母液,存于4 ℃低温下,其浓度进行调节后即可用于试验处理。
1.3 测定方法
1.3.1 形态指标和生物量测定
在胁迫试验的第10天(D1,所测总生物量为W1),第 20天(D2,所测总生物量为W2)时,选择每个梯度下的6株酸枣进行形态指标和生物量的测定,使用Li-3000C便携式叶面积仪(Li-Cor,USA)测定幼苗叶片植株叶长及叶宽。用游标卡尺(精度为0.001 cm)测定其基径,用卷尺(精度为0.1 cm)测定主根长度及幼苗株高。用无菌水对幼苗表层土屑进行冲刷,用滤纸将表层残留水分吸干,称取鲜质量,然后放于105 ℃烘箱中烘干杀青10 min,再于65 ℃下烘干,测定干质量,用电子分析天平(精度为0.0001 g)分别称取植物各部分的干质量后并进行累加得出植株总生物量,按公式(1)~(2)计算相对生长速率和根茎比(R/S)。

1.3.2 生理指标测定
指标测定均在处理第20天进行。每个处理选取幼苗嫩叶,放入96%的乙醇磨碎,于25 mL容量瓶中定容,在665、646、470 nm测定吸光值,计算各光合色素含量[16]。净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)测定,使用Ciras-2型便携式光合测定系统(PP Systems,USA)测定,光的强度为 1000 μmol/(m2·s)。
过氧化氢酶(CAT)活性采用紫外吸收法测定,过氧化物酶(POD)活性测定采用愈创木酚法,超氧化物歧化酶(SOD)活性测定采用氮蓝四唑(NBT)光化还原法,抗坏血酸氧化酶(APX)活性利用紫外分光光度法测定。
过氧化氢(H2O2)与丙二醛(MDA)含量测定参照李合生等[17]的方法,过氧根离子自由基产生速率测定参照Elstner 等[18]的方法,脯氨酸(Pro)测定采用酸性茚三酮比色法[17]。
质膜质子泵(H+-ATPase)活性测定参照Wang等[19]的方法;质膜钙泵(Ca2+-ATPase)活性测定参照缪颖等[20]的方法。将幼苗泡于乙二胺四乙酸二钠溶液中(2 mmol/L),除掉表面附着的微量元素,4 h后取出后用蒸馏水冲洗,并分离叶片和根系,杀青完置于80℃下烘干,用原子吸收光谱仪 AA6300 (Shimadzu,Japan)分析 Cd2+和微量元素含量[21]。
1.4 分析方法
数据采用SPSS 21.0 软件统计分析,多重比较(Duncan新复极差法)检验,Microsoft Excel 2003制表作图。显著性水平设定为α=0.05。
2 结果与分析
2.1 不同梯度镉处理酸枣幼苗的生长表现
由表2可知,不同浓度Cd2+处理酸枣幼苗后其生长指标均出现下降,当Cd2+浓度达到50 μmol/L,株高、地上鲜质量、地上干质量、地下鲜质量和地下干质量均显著低于CK(P<0.05),分别下降了25.68%、36.27%、33.33%、64.66%和58.62%。BT1和BT2的R/S均高于CK,BT2下值达到最大,与CK相比差异显著(P<0.05)。BT1与BT2的RGR比CK降低7.31%和37.26%,BT2与CK相比达到显著水平(P<0.05)。BT2的RGR与BT1相比差异显著(P<0.05),为植物RGR下降最快的,对植物生长造成了严重抑制。因此,选用50 μmol/L Cd2+为镉胁迫的处理浓度,即作为研究酸枣幼苗在逆境下对SNP的响应的最佳Cd2+浓度。

表2 不同梯度镉处理酸枣幼苗生长指标的变化Table 2 Effects of different Cd2+ contents on the growth index of Z. jujube seedlings
2.2 外源一氧化氮对镉胁迫酸枣生长的影响
由表3可知,单一Cd2+胁迫下,酸枣幼苗干质量、鲜质量、基径、根长、叶宽、叶长和叶数与CK相比显著下降(P<0.05),分别下降了44.74%,48.90%,6.67%,22.41%、11.92%、7.91%和6.12%。T2、T3与T4可对Cd2+胁迫下酸枣幼苗的抑制效应起到一定的缓解作用,T4的长势较CK出现较好改善,叶数、叶长、叶宽、根长、株高、基径、鲜质量、干质量分别比T1提高了13.96%、6.05%、10.62%、17.60%、5.62%、31.39%和43.24%,显著高于 T1(P<0.05)。T5、T6和 T7下植株生物量和形态指标呈下降趋势,对Cd2+胁迫下酸枣长势的影响越来越小,T5与T1差异均不显著,表明NO消除剂cPTIO的施加消除了SNP对酸枣幼苗Cd2+胁迫下的缓解效应,各指标与单一Cd2+胁迫处理接近;SNP相似物NaNOx、Na3Fe(CN)6处理下的酸枣幼苗生物量和形态指标长与Cd2+和T5并无较大差别,对逆境环境下引起的植株长势变差并无明显作用。因此从Cd2+胁迫下酸枣的生长趋势能够可以看出,NO可显著调节Cd2+胁迫对酸枣的负面效应,25~125 μmol/L SNP施加后较CK均出现了生物量的变化,其生物量积累程度随SNP浓度的不同出现差异,其中100 μmol/L SNP处理效果最好,而NO消除剂cPTIO、SNP类似物NaNOx、Na3Fe(CN)6并不能显著消除Cd2+胁迫对酸枣幼苗生物量和形态指标的负面作用。
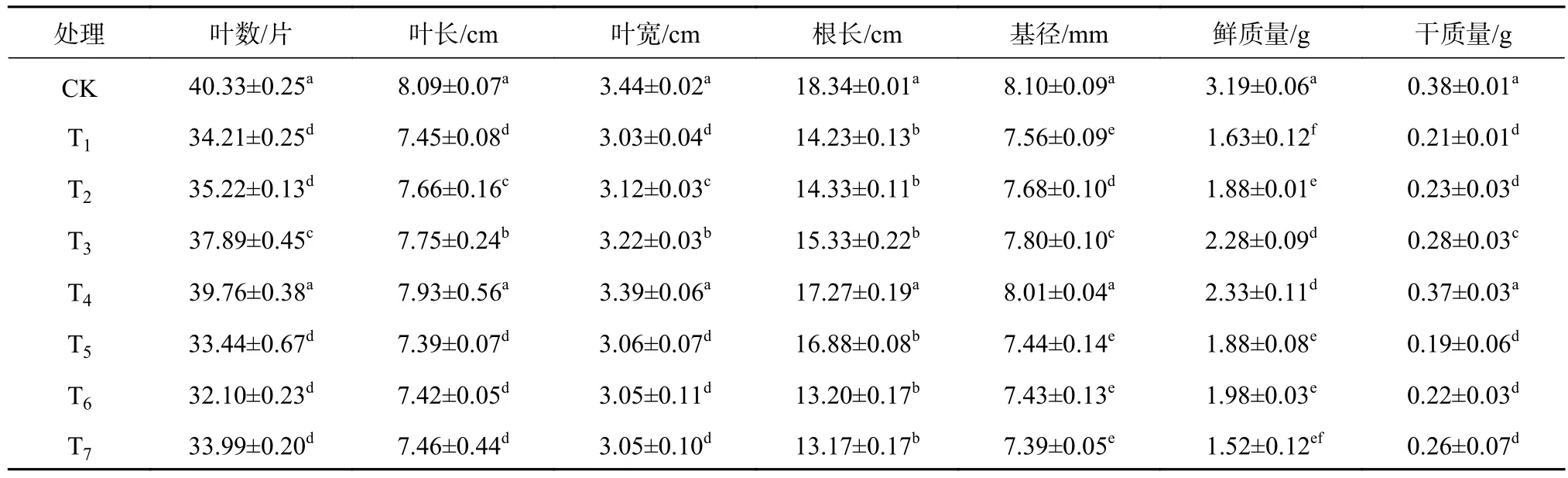
表3 镉胁迫酸枣幼苗生长指标对外源一氧化氮的响应Table 3 Effects of exogenous NO on the growth of Z. jujube seedling under Cd2+ sress
2.3 外源一氧化氮对镉胁迫酸枣光合特性的影响
由表4可知,与CK相比,Cd2+处理使酸枣幼苗叶片的叶绿素a含量、叶绿素b含量、类胡萝卜素含量、叶绿素a/b含量分别下降了49.24%,37.50%、56.67%和18.80%,差异达显著水平(P<0.05)。由于外源NO添加浓度的不同,其植株光合色素含量出现“钟形”变化趋势,其中T4的叶绿素a、叶绿素b的含量和叶绿素a/b的含量达到最大值,分别比T1提高了48.45%、34.52%、21.17%,差异均显著(P<0.05),T3的类胡萝卜素含量最大,且与T4并无显著差异。在添加cPTIO处理后,SNP的缓解效果被消除,在Cd2+处理液中加入 100 μmol/L NaNOx或 100 μmol/L Na3Fe(CN)6,与Cd2+胁迫处理差异不显著。

表4 外源一氧化氮对镉胁迫酸枣叶绿素含量和光合参数的影响Table 4 Effects of exogenous NO on the chlorophyll contents and photosynthetic parameters in Z. jujube under Cd2+ sress
同时,T1使酸枣叶片的Pn、Tr和Gs分别比CK下降了40.23%、52.79%、44.81%,Ci升高了42.51%,差异均显著(P<0.05)。在T2、T3和T4下加入不同浓度的NO大大缓解了Pn、Tr、Gs和Ci的变化趋势幅度,其中Ci处理的效果最明显,T4下的Pn、Tr和Gs分别比T1提高了57.40%、72.58%和62.45%,Ci降低了24.78%,差异均显著(P<0.05)。用 NO消除剂 cPTIO处理可明显抵消SNP对Pn的正面效应,Tr、Gs和Ci则并未出现显著差异,外施100 μmol/L的NaNOx或Na3Fe(CN)6对叶Pn、Tr、Gs影响不显著,对Ci影响显著。
2.4 外源一氧化氮对镉胁迫酸枣幼苗抗氧化酶系统的影响
由图1可知,施加Cd2+后显著降低植株SOD、CAT及叶片内APX含量(P<0.05),与CK相比,Cd2+胁迫下幼苗叶片和根系中SOD活性降低了29.17%和 39.06%,CAT活性降低了47.88%和58.62%,叶片中APX活性下降了51.63%,均显著低于CK(P<0.05),同时Cd2+胁迫下叶片和根系中POD活性与CK相比各自升高了36.11%和93.96%,差异显著(P<0.05)。T2叶片中POD活性略微下降,T3和T4仍继续呈上升趋势,但根系则呈现相反的趋势。T2、T3和T4大大减轻了Cd2+胁迫对SOD、CAT和APX酶活性的负面效应,其中T4和T1比较,幼苗叶片和根系内SOD活性升高了33.10%和57.22%,CAT活性升高了79.07%和91.67%,叶片内POD活性升高了13.27%,根系内POD活性下降了44.39%,叶片和根系内APX活性升高了98.96%和78.66%,差异均显著(P<0.05)。然而 100 μmol/L cPTIO、NaNOx和Na3Fe(CN)6并未显著影响Cd2+胁迫下幼苗叶片和根系内SOD、CAT和APX酶活性及叶片中POD活性,仅显著影响根系中POD含量。
2.5 外源一氧化氮对镉胁迫酸枣幼苗膜脂过氧化的影响
由图2可知,与CK比较,Cd2+胁迫后幼苗叶片和根部内H2O2、MDA、Pro总量及产生速率明显升高,表示Cd2+胁迫致使植物幼苗活性氧出现堆积状况,细胞内部遭受破坏,加深植物受损程度。与单一Cd2+胁迫比较,T3和T4的H2O2、MDA含量和产生速率都产生明显下降,T4幼苗叶片和根系中H2O2比Cd2+处理分别下降了44.93%和59.18%,MDA分别下降了66.67%和60.00%,比Cd2+处理分别下降了39.74%和42.31%,均差异显著(P<0.05)。添加外源SNP能够提高Pro含量,T4与CK相比达到显著水平(P<0.05),幼苗叶片和根系中Pro比Cd2+处理提高46.70%和49.43%,外施100 μmol/L SNP 后,H2O2、MDA 含量和产生速率最接近正常水平,质膜过氧化得到缓解。虽然大剂量SNP的施加使根中H2O2、MDA含量及产生速率有所下降,但并未低于Cd2+胁迫无外源SNP添加的程度,cPTIO的施加消除了NO于Cd2+胁迫后H2O2、MDA、Pro含量及产生速率堆积的负面效应,T5的叶片和根系H2O2、MDA积累及叶片内的产生速率高于T2、T3和T4,与单一Cd2+胁迫处理接近,根系中产生速率低于T2和T3,高于T4;Pro含量低于 T2、T3和 T4,与单一 Cd2+胁迫处理接近,SNP类似物NaNOx、Na3Fe(CN)6与cPTIO相似,对幼苗叶片和根系中H2O2、MDA、Pro含量及产生速率的作用与Cd2+和T5并无较大差别。

图1 外源一氧化氮对镉胁迫酸枣幼苗SOD、POD、CAT和APX活性的影响Fig. 1 Effects of exogenous NO on activity of SOD, POD, CAT and APX of Z. jujube seedlings under Cd2+ stress

图2 外源一氧化氮对镉胁迫酸枣幼苗H2O2、MDA、Pro含量和产生速率的影响Fig. 2 Effects of exogenous NO on H2O2, MDA, Procontents and production rates of Z. jujube seedlings under Cd2+ stress
2.6 外源一氧化氮对镉胁迫酸枣幼苗镉富集量的影响
由图3可知,CK组根内Cd2+总量为0,单一Cd2+处理后,酸枣幼苗叶片和根系内Cd2+总量大幅度升高,无论高低剂量的SNP都可以使Cd2+胁迫下的酸枣幼苗叶片与根系中Cd2+总量出现减小,但整体并未大于Cd2+胁迫无外施SNP处理的Cd2+含量。T4叶片和根系内Cd2+含量比Cd2+处理显著降低了52.24%和 59.90%(P<0.05)。同一处理下,其叶片所含Cd2+总量显著低于根系中的含量(P<0.05),其根系为Cd2+所趋向堆积的部位。加入cPTIO、NaNOx、Na3Fe(CN)6,叶片和根系Cd2+总量均大于T4,但并未达到显著水平,同时cPTIO、NaNOx、Na3Fe(CN)6各处理组下的Cd2+含量总体低于Cd2+处理组。由此可见,单一Cd2胁迫可以导致酸枣幼苗叶片和根系中内源Cd2+聚集,外源NO能明显降低Cd2+胁迫后根系内源Cd2+的堆积。
2.7 外源一氧化氮对镉胁迫酸枣幼苗质膜ATPase活性的影响
由图4可知,Cd2+胁迫后幼苗叶片与根系质膜H+-ATPase和Ca2+-ATPase活性都出现大幅度降低,与CK相比,幼苗叶片和根系质膜H+-ATPase分别显著降低了34.55%和43.82%(P<0.05),Ca2+-ATPase分别显著降低了 30.66%和31.11%(P<0.05)。添加不同剂量的SNP,幼苗叶片和根系质膜H+-ATPase和Ca2+-ATPase活性逐渐升高,T4的H+-ATPase比T1提高了40.64%和 88.97%,Ca2+-ATPase比 T1提高了 30.60%和44.85%,与CK相比达到显著差异(P<0.05),添加100 μmol/L的SNP能使ATPase活性提高到正常水平,而外施 100 μmol/L的 cPTIO、NaNOx、Na3Fe(CN)6下,植物幼苗质膜H+-ATPase、Ca2+-ATPase活性和单一Cd2+胁迫没有明显差别。
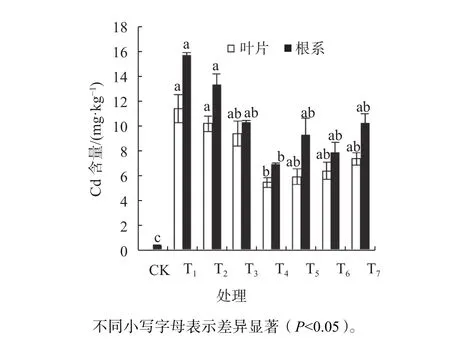
图3 外源一氧化氮对镉胁迫酸枣植株中Cd2+总量的影响Fig. 3 Effects of exogenous NO on Cd2+ contents in Z. jujube under Cd2+ stress
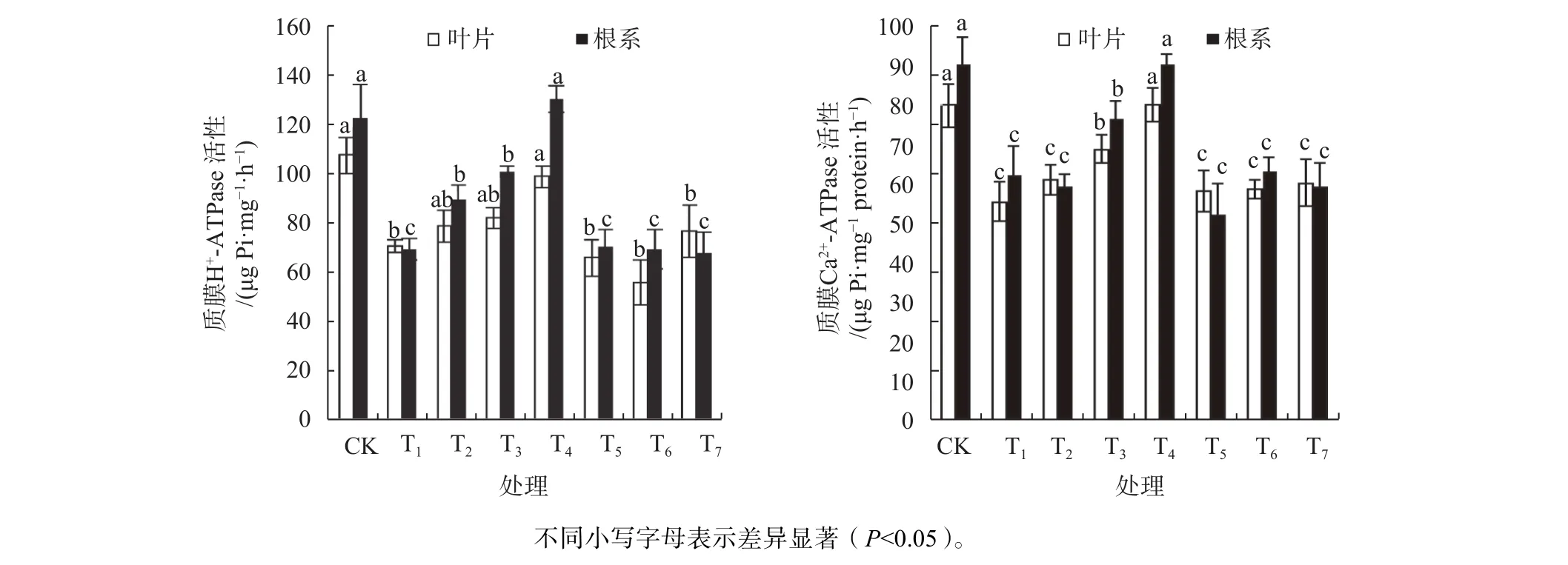
图4 外源一氧化氮对镉胁迫酸枣质膜H+-ATPase与Ca2+-ATPase活性的影响Fig. 4 Effects of exogenous NO on H+-ATPase and Ca2+-ATPase activities in plasma membrane of Z. jujube under Cd2+ stress
2.8 外源一氧化氮对镉胁迫酸枣幼苗矿质营养含量的影响
由表5可知,在酸枣叶片中,Cd2+胁迫降低对钾(K)、钙(Ca)、镁(Mg)、铁(Fe)、铜(Cu)、锌(Zn)的吸收,相比CK分别降低了 63.87%、49.80%、32.66%、50.20%、47.54%、33.51%,差异显著(P<0.05),同时Cd2+胁迫显著提高了对锰(Mn)的吸收(P<0.05),相比CK提高了115.17%;而在酸枣根部内,Cd2+胁迫与 CK相比显著降低了对钾 K、Ca、Mg、Fe、Zn、Mn的摄入(P<0.05),同时显著提高了植株对Cu的吸收(P<0.05)。在酸枣叶片中,加不同浓度的SNP提高了植株叶片对K、Ca、Mg、Fe、Cu、Zn和Mn的摄入,其中在T4下作用最显著,分别相比T1显著提高了141.75%、86.01%、50.50%、94.93%、81.64%、53.18%和134.81%(P<0.05);但在酸枣根系内,加小剂量到大剂量的SNP,植株叶片对Ca、Mg、Fe和Zn的摄入也呈递增趋势,在T4下最大,分别相比T1显著提高了67.84%、33.80%、86.76%和37.05%(P<0.05),K和Cu的摄入量为先减小后增高,于T2下最低,T4比T1提高了8.67%和7.35%,差异并不显著,Mn无明显变化规律,在T3下达到最大。而外 施 100 μmol/L 的 cPTIO、NaNOx、Na3Fe(CN)6下对植株叶片的微量矿质元素无显著影响,cPTIO并没有对植株根系的矿质元素产生较大作用,NaNOx明显降低了植物幼苗根部的Mn含量,Na3Fe(CN)6的加入大大提高了了根系Fe总量,使根系内Cu总量出现下降,这可能与T7引入Fe元素有关。

表5 不同浓度SNP对镉胁迫酸枣矿质营养含量的影响Table 5 Effects of different concentrations of SNP on mineral nutrition in shoots and roots of Z. jujube under Cd2+ stress g/kg
3 结论与讨论
Cd2+是植物生长的非必需元素,Cd2+胁迫可引起植物生理生化方面复杂的变化,能够使内脂氧合酶和还原性辅酶II(NADPH)氧化酶活性提高[22],膜质过氧化和细胞内代谢出现紊乱,最后导致植物长势变差。本试验观察得知,Cd2+中毒的酸枣植株矮小,老叶叶脉间和叶尖失绿,新叶卷曲、老叶出现较多Cd2+中毒的褐色斑点,植物叶性状和基径等均遭受抑制,表明50 μmol/L Cd2+处理使其生理指标值显著减小,使植物的生长状况变差,这进一步验证了前人的结论[23]。试验过程中观测到,植物幼苗根部受到的胁迫伤害较叶片更为严重,其原因为本试验采用的为营养液培养方法,其Cd2+能与植物根部大面积直接接触,根部内Cd2+高于叶片。外源SNP处理后,Cd2+中毒的症状得到明显改善,酸枣生物量明显增加,NO对提高酸枣抗Cd2+胁迫的作用明显,外源NO对Cd2+处理下氧化伤害的缓解作用已在紫花苜蓿(Medicago sathiva)[24]、东南景天(Sedum plumbizincicola)[25]、蚕豆(Vicia faba)等[26]多个物种中得到证明。Cd2+于酸枣幼苗根部存在截留效应,可以和根部中的谷脱甘肽和含硫化合物与其结合构成螯合物发挥解毒效果,根长随着浓度的提高逐渐增长,表示适宜的SNP剂量调节了根系细胞壁内交换位点的转移,使根系生长速度加快,加大了对矿质营养元素的摄入。刘柿良等[27]、Wang等[28]在研究发现了施加NO对植物幼苗的大小浓度-双面现象,这都显示了植物存在“低促高抑”的状况,但在本研究中,植物各生理指标在T4恢复到与CK相近的水平,因此并未设置过多试验组。NO消除剂cPTIO、SNP类似物NaNOx、Na3Fe(CN)6的应用却没有缓解Cd2+诱导的氧化胁迫伤害,这与李海燕等[29]对外源NO对Cd胁迫下玉米(Zea maya)幼苗根的研究中得出cPTIO的加入逆转了SNP保护作用的结论出现了差异,而本研究与此相悖的结果可能与植物物种、器官、发育阶段、胁迫处理的方式、浓度和时间等有关。因此可以判断SNP于Cd2+胁迫下酸枣幼苗生理指标的回升现象为NO导致的,NO很大部分上是通过减少氧化伤害来提高Cd2+耐性的。
汤绍虎等[30]研究表明适宜的SNP能够有效提高黄瓜(Cucunis sativus)的光系统活性、光合速率和产量。光合色素含量是表示植株叶片中光合结构的重要组成部分,其受到胁迫后都会对其含量产生不同程度的改变[31]。Cd2+对植物胁迫作用最明显的特征是植株叶片黄化,Cd2+胁迫后,其叶绿素含量开始出现下降,且叶绿素含量与Cd2+浓度为负相关变化趋势,由此也能够得出,随着Cd2+胁迫浓度的不断加强,叶片的光合速率显著下降,施加NO后又显著增加,从而保证酸枣光合速率出现回升。本研究中,酸枣幼苗于Cd2+胁迫下,Pn大幅度下降,这与叶绿素含量的降低有密切关系;Pn和Gs下降的同时Ci出现上升的趋势。Kopyra等[32]研究表明,施加外源NO能够明显提高Cd2+胁迫下羽扇豆(Lupinus polyphyllus)根尖SOD、CAT、和APX活性,本研究结论与此相似,Cd2+处理显著降低了酸枣SOD、CAT、APX活性,在Cd2+处理下添加外源NO提高了SOD、CAT、APX活性,POD在Cd2+胁迫及施加NO后一直呈上升趋势,这与廖克波[33]对观光木(Michelia odora)在Cd2+胁迫时的变化规律相同。与正常生长的植株相比,Cd2+胁迫致使酸枣幼苗体内MDA和H2O2、含量上升,这与Rodriguez等[34]对豌豆(Pisum sativum)和 Yi等[35]对水稻(Oryza sativa)的研究结果相一致,表明抗氧化物质为逆境环境里限制植株生长发展的关键物质,而100 μmol/LNO显著降低了Cd2+胁迫下叶片细胞MDA和H2O2、的含量,且减小细胞膜透性的上升幅度和细胞膜的变化,缓解了Cd2+胁迫对植株造成的氧化伤害。Pro一方面经由催化鸟氨酸生成,另一方面也可以催化谷氨酸形成,之后会存在降解过程,所以Pro总量取决于这2个过程总的强弱。在此实验结果中显示,Cd2+单一处理后,酸枣内产生大量的Pro,施加外源NO则进一步加速了Pro的这种变化趋势,这和Fan等[36]、Dilek等[37]的试验结果相一致。在NO缓解胁迫损伤的机理,有2种说法被国内外学者认可:其一,NO可直接与活性氧作用,例如可与形成过氧硝酸盐,这种物质对细胞的伤害相对较轻,因而起到了保护细胞的作用;其二,NO可以作为一个信号分子,激活细胞内的抗氧化系统,而这些原因则需在后一步的分子生物学上得到验证。本研究在添加cPTIO后,SNP的效果被显著消除,施用NOx-和Na3Fe(CN)6,并无法显著改变Cd2+胁迫对幼苗抗氧化酶以及MDA和H2O2等总量的大小。植物摄入的Cd+于各部位的总量排列顺序分别为根>茎>叶>籽粒[38]。本研究也表明,酸枣根系中Cd2+的浓度高于叶片,Cd2+于根部内的堆积一方面是因为细胞壁上羧基的交联作用,另一方面则是由于可溶性蛋白中硫基的交互作用,而叶中的Cd2+含量远低于根部可能是因为NO防止幼苗膜质过氧化并维持细胞膜的稳定,Cd2+很难入侵到地上部分的细胞中。这种将Cd2+富集于根系是植物阻止Cd2+胁迫的生存策略。Cd2+处理使酸枣根部内H+-ATPase和Ca2+-ATPase活性显著下降,而NO能够使Cd2+胁迫下酸枣叶片和根部细胞质膜 H+-ATPase和 Ca2+-ATPase活性出现上升,其原因可能是由于施加供体SNP后,细胞膜的结构和稳定性得到恢复,给H+-ATPase和Ca2+-ATPase活性创造了较好的存在和进一步反应的微环境,张鑫荣[39]对巨峰葡萄(Vitis vinifera)的研究更好的验证了以上结论。在幼苗体内,各种离子的含量及种类一直处于一个动态平衡的过程中。然而,Cd2+胁迫打破了这种平衡,Cd2+胁迫降低了酸枣体内K、Ca、Mg、Fe、Cu、Zn的总量,NO提高了酸枣在Cd2+胁迫下对矿质营养元素的摄入。综上所述,Cd2+胁迫会对植物生长产生抑制,而Cd2+胁迫下一定浓度的外源 SNP 处理能够通过增强活性氧清除能力,提高光合色素总量和质膜ATP酶活性,进一步加强叶肉细胞光合性能,提升离子与信号分子跨膜转换,维持矿质营养元素平衡,缓解镉胁迫对细胞质膜的损伤,增强酸枣幼苗对Cd2+毒害的抗性,使 Cd2+对酸枣的毒害得到一定程度的缓解,其中浓度以100 μmol/L缓解胁迫效果最好,而NO消除剂cPTIO、SNP类似物NaNOx、Na3Fe(CN)6并无相同作用。
猜你喜欢
今日农业(2022年13期)2022-09-15 01:18:00
舰船科学技术(2022年11期)2022-07-15 07:51:56
中成药(2021年5期)2021-07-21 08:38:46
河北果树(2020年1期)2020-02-09 12:31:42
西藏农业科技(2019年3期)2019-11-04 00:35:10
现代园艺(2018年3期)2018-02-10 05:18:12
上海农业学报(2017年3期)2017-04-10 12:39:12
中华胃食管反流病电子杂志(2016年4期)2016-11-07 02:38:26
中国医学科学院学报(2015年5期)2015-03-01 04:03:36
中国当代医药(2015年17期)2015-03-01 02:03:39
