
不同低温环境对烟苗理化性的影响
2019-04-12 03:59:00殷雪艳
陕西农业科学 2019年2期
周 然,杨 柳,殷雪艳,刘 彪,赵 爽
(四川省烟草公司 凉山州公司,四川 西昌 615000)
引言
烟草作为一种喜温植物[1],在高温或低温对烟草进行胁迫时,促使烟草理化性发生改变[2-4],如幼苗承受低温胁迫的程度具有一定的限度,即使在短时间的低温胁迫条件,不影响烟草的生存与繁殖,但对烟叶的产量和质量造成严重的影响[5],如叶片数量降低、叶片展开程度降低,氮类化合物和碳类化合物比例失调[6]等,其中游离型脯氨酸(PRO)含量在氮类化合物占有重要的地位,尤其在低温胁迫时,通过自身的变化,调节植物体的内在环境,进而减少低温对植物造成的影响[7];丙二醛(MDA)含量在碳类化合物占有重要的地位,它属于过氧化物被分解的产物,能够较为灵敏的响应低温胁迫,其含量的变化幅度具有体现植物对低温的耐受性[8];导电率具有反映植物细胞在低温胁迫下的破损程度,以及细胞体内总体物质成分的变化[9]。因此,作者通过研究烟草幼苗在不同低温胁迫的条件下,以及相同低温胁迫下的不同品种的导电率、PRO含量和MDA含量,以期该理化性指标反映烟草品种的自身特性,以及作为烟草幼苗在低温胁迫下的特性指示,为指导烟农的合理种植提供参照。
1 材料与方法
1.1 试验材料
供试的烟草品种为云烟85和K326,开展试验地为会理县四川中烟基地单元。
1.2 试验设计
筛选籽粒饱满,大小较为一致的云烟85和K326两个品种,并于基地单元中的温室大棚内采用漂浮法进行育苗,其光照要求为自然光,待烟株幼苗生长至6片真叶时,每个品种选择大小一致的有品种200株,并将其分别分为4组,其中云烟85品种分为A1、A2、A3和A4组,K326品种分为B1、B2、B3和B4组。各组烟株幼苗均已栽至培养基成分相同,并等量的花盆中,于25℃,自然光照的条件下培养48h。待烟株幼苗返苗后,将A1和B1组作为对照组,即烟株幼苗放置于25℃的人工智能气候箱中进行低温处理;A2和B2组烟株幼苗放置于4℃的人工智能气候箱中进行低温处理;A3和B3组烟株幼苗放置于6℃条件下进行低温处理;A4和B4组烟株幼苗放置于10℃条件下进行低温处理。对烟株幼苗进行低温处理24h后,取5株烟草幼苗的第4片真叶,并混合,测定其导电率、PRO和MDA含量。
1.3 测定项目及方法
导电率性测定参照尹航运用电导仪测定烟草的相对导电率[10];PRO含量的测定,利用含量磺基水杨酸、茚三酮和甲苯试剂,采用分光光度法测定其含量[11],MDA含量的测定,运用硫代巴比妥酸法测定其含量[12]。
2 结果与分析
2.1 不同低温处理对导电率的影响
图1显示,烟草幼苗通过低温处理后,云烟85和K326 叶片内导电率随着温度的降低,而呈现升高的趋势,其中在4 ℃低温胁迫的条件下,导电率达到最大值,云烟85和K326的导电率分别为27.7%和34.6%。除对照组外,低温胁迫致使两品种之间的导电率存在显著性差异(p<0.05),即在相同的处理组之间,云烟85小于K326叶片内导电率。对于同一个品种不同的的低温胁迫,云烟85叶片的导电率在10 ℃条件下,上升幅度较小(9.4%),与对照组无显著性差异,但与4和6 ℃条件下的叶片导电率存在显著性差异,此外,对照组同样与4 ℃和6 ℃条件下的叶片导电率存在显著性差异,而与4 ℃和6 ℃的低温胁迫处理组的增幅分别为75.4%和31.2%,且两组之间存在显著性差异;K326各处理之间的叶片导电率,其显著性差异规律与云烟85相同,其在4 ℃、6 ℃和10 ℃低温胁迫下,导电率的增幅分别为89.1%、40.9%和12.8%。
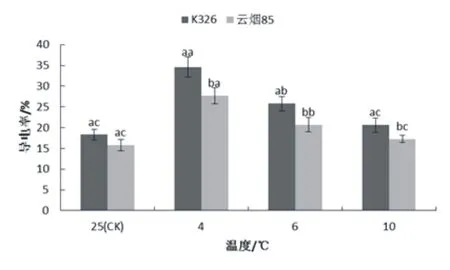
图1 低温影响导电率的变化

图2 低温影响PRO含量的变化
2.2 不同低温处理对PRO含量的影响
图2显示,烟草幼苗经低温处理后,云烟85和K326的PRO含量均呈现较为相似的变化规律,即PRO含量升高,且低温胁迫的温度越低,其含量越高,但云烟85的PRO含量始终大于K326,且两品种间存在显著性差异(p<0.05)。由于烟草幼苗的品种不同,各处理间PRO含量增幅有所差异,云烟85幼苗在4、6和10 ℃低温胁迫的条件下,其PRO含量均与对照组存在显著性差异,且各低温胁迫组之间存在显著性差异,其增幅分别为195.3%、142.9%和83.6%;K326幼苗在低温的胁迫下,其各组之间PRO含量的显著性差异,以及与对照组之间的显著性差异的规律,与云烟85相同,K326幼苗在4、6和10 ℃低温胁迫下,其叶片中PRO含量的增幅分别为151.7%、112.6%和65.8%。云烟85和K326烟草幼苗在响应低温胁迫时,叶中PRO含量和增幅程度有所差异,表明两者防御低温胁迫的代谢途径可能有所差异。
2.3 不同低温处理对MDA含量的影响
云烟85和K326烟草幼苗经低温胁迫后,两者的酶活性均大于对照组,但随着温度的降低,呈现下降的趋势,即低温胁迫温度为4 ℃时,烟草幼苗的MDA含量最低,其次是低温胁迫温度为6 ℃时,最后是低温胁迫温度为10 ℃的烟草幼苗。图3表明,云烟85和K326烟草幼苗在相同的温度处理间,云烟85小于K326,但对照组之间不存在显著性差异,而其余组之间的两个品种的MDA含量存在显著性差异(p<0.05)。在相同的品种,不同的处理组之间,云烟85的低温胁迫组的MDA含量均与对照组存在显著性差异,而4和6 ℃低温胁迫组的MDA含量无显著性差异,但均与10 ℃低温处理组存在显著性差异,各低温胁迫组MDA含量的增幅分别为41.2%、78.4%和133.8%;K326烟草幼苗在4、6和10 ℃低温胁迫下,其MDA含量均与对照组存在显著性差异,且各胁迫组之间同时存在显著性差异,其MDA含量的增幅分别为53.4%、95.7%和163.2%。综上所述,低温胁迫在一定程度上影响到烟草幼苗叶片内MDA含量,使其含量升高,此外,表明该物质可作为烟草幼苗响应低温胁迫的特性指标。

图3 低温影响MDA含量的变化
3 结论与讨论
云烟85和K326烟草幼苗受到低温胁迫时,导电率随温度降低,而呈升高的趋势,与尹航等研究的结果较为一致[10],其原因可能为低温造成细胞膜的通透性改变,即通透性增加,破损程度增加;PRO含量随温度降低,呈逐渐升高的趋势,其研究结果与晋艳等研究较为一致[1],呈现该结果的原因为脯氨酸自身组分的积累,改善细胞的渗透性等,进而增加植物对低温胁迫的抗性;MDA含量随温度降低,逐渐降低,但均大于对照组,与尹航等研究有所差异[10],其原因可能为烟草品种,且尹航等研究的不同品种,具有不同的变化规律。肖玉洁[13]等认为耐低温型品种的植物,其导电率和MDA含量低,且PRO含量高,而低温敏感型品种于此相反,因此,云烟85可能属于耐低温型,K326可能属于低温敏感型。
猜你喜欢
奥秘(创新大赛)(2023年3期)2023-05-06 01:48:20
农业技术与装备(2020年10期)2020-11-03 11:25:44
浙江中西医结合杂志(2017年2期)2017-01-12 18:23:59
当代化工研究(2016年9期)2016-03-20 16:22:08
河北书画研究(2016年6期)2016-02-03 02:57:44
大连工业大学学报(2015年4期)2015-12-11 04:06:50
智能建筑电气技术(2015年5期)2015-12-10 05:52:23
诗歌月刊(2014年3期)2015-04-14 17:50:51
传奇故事(破茧成蝶)(2015年8期)2015-02-28 09:29:41
发明与创新(2015年30期)2015-02-27 10:39:51
