
退耕还湖后不同植被群落湿地土壤剖面磷素形态分布特征①
2019-04-08 09:09包先明
土壤 2019年1期
包先明,崔 宏
退耕还湖后不同植被群落湿地土壤剖面磷素形态分布特征①
包先明1,崔 宏2,3
(1 淮北师范大学生命科学学院,安徽淮北 235000;2 临泉第二中学,安徽临泉 236400;3安徽自然灾害过程与防控研究省级重点实验室,安徽芜湖 241000)
对菜子湖退耕还湖区不同植被群落(苔草、芦苇和酸模)下湿地剖面土壤进行分层采样,分析了土壤剖面有机磷(orP)、无机磷(inP)及无机磷组分铝磷(Al-P)、铁磷(Fe-P)、闭蓄态磷(O-P)和钙磷(Ca-P)的含量,探讨了植被群落对退化湿地生态恢复后土壤剖面磷素形态分布特征的影响。结果表明:研究区土壤剖面磷素以无机磷为主(301.94 ~ 645.17 mg/kg)占全磷含量的52.59% ~ 84.64%;除酸模群落下0 ~ 6 cm土层外,土壤剖面有机磷含量范围为78.40 ~ 254.27 mg/kg。土壤剖面无机磷组分以钙磷和铁磷为主,分别占无机磷总量的29.67% ~ 67.58% 和21.90% ~ 52.29%。除酸模群落下0 ~ 6 cm土层外,3种植被群落下土壤无机磷含量均随着剖面深度增加而增加,而有机磷含量则随着剖面深度增加先急剧降低后逐渐升高;土壤铝磷和铁磷含量总体上均随着剖面深度增加而降低;除酸模群落下0 ~ 6 cm土层土壤闭蓄态磷外,其他土壤剖面闭蓄态磷和钙磷含量总体均随着剖面深度增加而升高。研究区3种植被群落下土壤铝磷、铁磷和有机磷存在表聚现象,酸模群落下土壤剖面表现最为显著。研究区植物对土壤磷素吸收利用是磷素形态转化和表聚的驱动力,而钙磷是各形态磷表聚的主要来源。
植被群落;湿地土壤;土壤剖面;磷素形态
湿地是位于陆地和水生生态系统之间的过渡带,具有独特的水文、土壤、植被与生物特征,是地球表层各种生源要素的源、汇和转化的场所[1]。磷是植物生长必需元素之一,决定着湿地结构、功能和生产力的重要元素,也是湿地生态系统主要的限制性因子[2-3]。磷在湿地土壤中以有机磷和无机磷的形态存在[3]。有研究表明闽江口鳝鱼滩湿地芦苇和短叶茳芏沼泽土壤以无机磷为主要组分,其中无机磷主要贡献者为铁磷和闭蓄态磷[4];三峡水库淹没消落区土壤中的磷也以无机磷为主,有机磷为辅,而主导土壤全磷含量的是无机磷中的钙磷,其次为有机磷[5];而美国阿卡迪亚公园湿地土壤磷以铝磷为主,可占全磷62% ~ 66%[6];菜子湖退耕还湖区湿地土壤磷素以无机磷为主,其中铁磷和钙磷占无机磷主导地位[7];三江平原湿地小叶章草甸土壤全磷以有机磷为主,无机磷为辅[8]。因此,不同研究区湿地土壤磷组分各不一致,受研究区成土母质、植被、水文条件及利用方式等因素影响[8-12]。
湿地土壤磷形态变化深受植物生长节律影响[8],水分、热量等环境条件也是磷形态转化的重要驱动因子[8-11]。湿地植被自然演替过程中不同植物生物量积累和营养物质循环过程的变化导致沉积物中磷形态的差异[12]。植物还可通过与微生物协同机制作用下,对土壤磷素地球化学过程产生影响[13]。而不同植物吸收土壤磷素的最佳时期存在差异,对土壤有效磷吸收也存在差异,且吸收有效磷的来源也存在差异[14-15],进而影响了湿地土壤磷素形态含量及分布。胶州湾盐沼不同植被群落下土壤磷含量存在差异,互花米草土壤全磷含量最高,而芦苇湿地土壤则最低,但碱蓬土壤有效磷含量最高,0 ~ 20 cm土壤全磷含量最高,随着深度增加而降低[16];宋晓琳等[17]研究表明,在不同覆被条件下双台子河口湿地土壤全磷含量在0 ~ 30 cm深度内变化不大,30 cm以下土壤全磷含量升高。因此,湿地植物可通过协同微生物群落活性、吸收根际分泌物和凋落物等影响湿地土壤磷地球化学过程[3,8,13],进而影响湿地土壤磷素组分特征、磷素有效性及全磷含量。
综上所述,有关湿地土壤磷素形态研究主要集中在不同类型湿地土壤磷形态组分含量及特征、水分条件和植被状况对湿地土壤磷形态组分特征的影响上,从土壤剖面视角去研究植物如何影响湿地土壤磷形态特征的研究较少。1998年长江特大洪水后,退耕还湖成为我国长江中下游湿地生态恢复的重要举措。退耕还湖后湿地生态系统功能恢复成为湿地科学研究的重要领域。以土壤剖面为视角不仅有利于研究湿地生态恢复下植被对湿地土壤磷形态的影响,也有利于研究植被类型对不同磷形态垂向迁移的影响,可进一步丰富植被影响湿地土壤磷素生物地球化学过程的认识。本文选取安徽省安庆市菜子湖退耕25 a圩区苔草、芦苇和酸模植被下湿地土壤为研究对象,对土壤剖面有机磷和无机磷组分进行分析比较,探究植被群落对磷素组分在土壤剖面上分布特征的影响,可为退化湿地生态恢复中选择有利于土壤磷素养分保持的植被类型提供参考。
1 材料与方法
1.1 研究区概况
安庆菜子湖湿地是安徽省沿江湿地自然保护区的重要组成部分,地理位置117°01′ ~ 117°09′ E,30°43′ ~ 30°58′ N,总面积17 300 hm2。菜子湖流域属亚热带季风气候区,气候温和,雨量丰沛,多年平均气温16.5℃,多年平均降雨量1 389.1 mm,多年平均蒸发量1 611.4 mm。20世纪50年代开始大规模的围湖造田,20世纪80年代开始退田还湖,许多退耕还湖区因地制宜地选择了恢复为自然湿地或进行水产养殖,旨在寻求湿地生态调蓄功能与经济收益间的平衡[18]。
1.2 土样采集与处理
采样区位于菜子湖金神镇幸福圩(30°51.028′ ~ 30°51.043′ N,116°00.970′ ~ 117°00.998′ E),该圩建于1958年,于1986年破圩,恢复为自然水域,后进行水产养殖。依据典型性和代表性原则,选取苔草、芦苇和酸模3种不同植被群落湿地(图1)作为研究样地。苔草样地苔草高度为80 ~ 100 cm,有倒伏,盖度约100%;芦苇样地芦苇新株高度200 cm左右,立枯死株高度330 cm左右,盖度约100%;酸模样地植株高度约100 cm,盖度约90%[18]。每种类型植被下湿地采3个混合土样,每一个混合土样由3个点混合而成。采样时,发现各个样点土壤剖面均在6 cm处发生明显的颜色、紧实度和根系多少的转变,所以本研究按0 ~ 6、6 ~ 13、13 ~ 25、25 ~ 40、40 ~ 55 cm 采集5个层次土壤样品,共采集45个混合土样[18]。土壤样品经自然风干,挑去根、虫体、石砾,研磨,过20目塑料筛及分取部分再研磨过100目塑料筛,装袋备用。采样区不同植被群落下土壤理化性质见文献[18]。
(SM、LW和TC分别为酸模、芦苇和苔草样地)
1.3 分析方法
无机磷(inP)分级采用文献[19]连续浸提分级方法,分为铝磷(Al-P)、铁磷(Fe-P)、钙磷(Ca-P)和闭蓄态磷(O-P);有机磷(orP)含量测定采用烧灼法[20]。
1.4 数据处理
利用SPSS(17.0)及Excel2003进行数据处理、制图及分析。
2 结果与分析
2.1 不同植被群落下湿地土壤剖面无机磷和有机磷分布特征
土壤中的全磷可分为无机磷和有机磷两大部分,其中无机磷是磷素的一个极其重要的组成部分,是植物所需磷的主要来源,其含量占全磷的60% ~ 80%[3,20-21]。研究区3种植被群落湿地土壤无机磷含量在301.94 ~ 645.17 mg/kg(图2),占全磷含量的52.59% ~ 84.64%(表1)。除酸模植被下0 ~ 6 cm土层外,3种植被群落下土壤无机磷含量均有随着土壤剖面深度增加而增加的趋势,相同土壤剖面深度无机磷含量存在显著差异;但0 ~ 6 cm土层无机磷占全磷比例显著低于6 ~ 55 cm各个层次。同时总体上,酸模样地各个深度土壤剖面无机磷含量显著高于其他2种植被样地(表1)。土壤有机磷是土壤磷素的重要组成部分,对土壤肥力和植物营养有着重要影响,我国大部分土壤有机磷占全磷的20% ~ 40%[20]。东北三江平原小叶章草甸土壤有机磷总量高于无机磷[8],而研究区湿地土壤有机磷含量除酸模植被下0 ~ 6 cm土层为581.67 mg/kg外,含量范围在78.40 ~ 254.27 mg/kg,占全磷比例为15.36% ~ 47.41%(表1)。不同植被对有机磷的影响主要表现在0 ~ 6 cm土层,酸模群落下该层有机磷含量高于苔草群落下,而苔草群落下土壤有机磷含量高于芦苇群落下,6 cm以下各个深度土壤剖面有机磷含量无显著差异,但均存在随着土壤剖面深度增加先急剧下降再逐渐升高的趋势。有机磷占全磷比例也存在系统趋势(表1)。
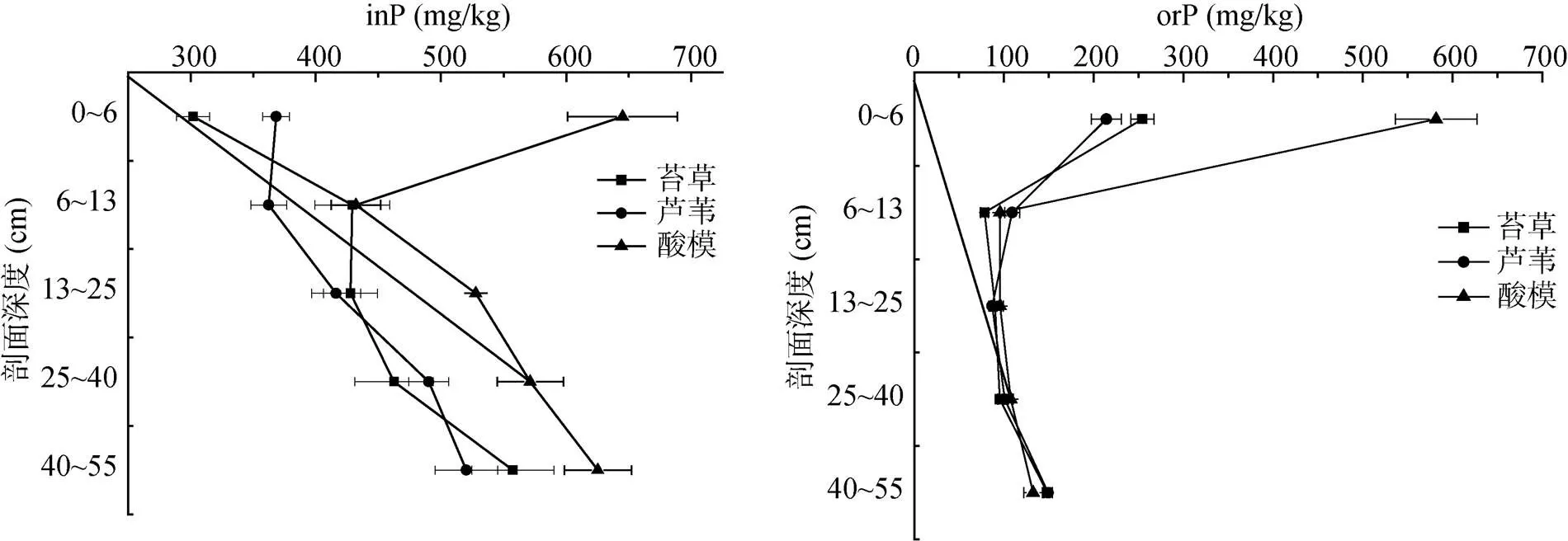
图2 不同植被群落湿地土壤剖面土壤无机磷(inP)和有机磷(orP)含量的变化
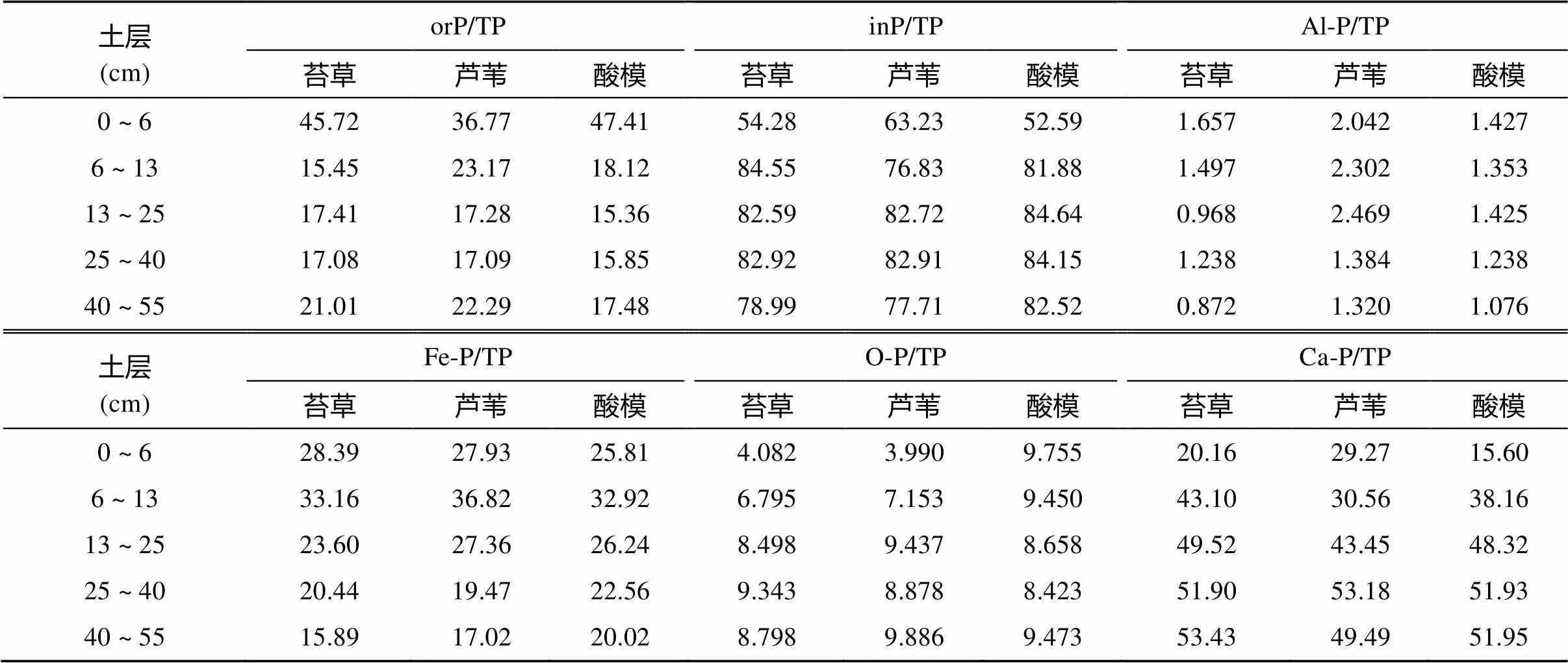
表1 不同植被类型湿地土壤剖面中不同形态磷占全磷的比例(%)
注:表中TP表示全磷,inP表示无机磷,orP表示有机磷,Al-P表示铝磷,Fe-P表示铁磷,Ca-P表示钙磷,O-P表示闭蓄态磷;下同。
2.2 不同植被群落下湿地土壤剖面各形态无机磷分布特征
土壤中磷素的形态决定磷素有效性,恰当的磷素分级方法能较好地评价土壤有效磷库的大小和磷素的供应状况[3,21]。研究区3种植被群落下湿地土壤剖面无机磷形态分布见图3,各形态无机磷占全磷量的百分比见表1。研究区湿地土壤所有剖面铝磷含量范围5.01 ~ 17.50 mg/kg,占无机磷总量的1.11% ~ 3.23%,占全磷量的0.872% ~ 2.49%;铁磷含量范围为112.15 ~ 316.60 mg/kg,占无机磷总量的21.90% ~ 52.29%,占全磷量的15.89% ~ 36.82%;闭蓄态磷含量范围为22.71 ~ 119.69 mg/kg,占无机磷总量的6.31% ~ 18.56%,占全磷量的3.990% ~ 9.886%;钙磷含量范围为112.11 ~ 393.68 mg/kg,占无机磷总量的29.67% ~ 67.58%,占全磷量的15.60% ~ 53.43%。所有无机磷组分中铝磷含量最低,所占比例最小,而钙磷和铁磷占无机磷主导地位,是土壤无机磷主要组分。有研究表明向海湿地[22]]和三峡库区消落带湿地[23]土壤无机磷以Ca-P为主,而闽江口鳝鱼滩湿地芦苇和短叶茳芏沼泽土壤以无机磷主要以铁磷和闭蓄态磷为主[4]。本文结果与同地区不同退耕年限下菜子湖湿地土壤无机磷组分以铁磷和钙磷为主导地位[3]一致。
研究区不同植被下湿地土壤铝磷存在差异,芦苇群落和酸模群落下土壤相对应剖面层铝磷含量高于苔草。除酸模植被下0 ~ 6 cm土层土壤铝磷含量显著高于其他群落外,芦苇植被下0 ~ 25 cm土层土壤剖面铝磷的含量高于其他植被群落土壤剖面相对应层。总体上3种植被群落下土壤铝磷均存在随着土壤剖面深度增加,铝磷含量先降低后升高现象,占全磷比例总体上也存在逐渐降低趋势(表1)。酸模群落下湿地土壤铁磷含量最高,而苔草与芦苇植被下基本一致,但3种植被群落下土壤剖面铁磷含量均存随着剖面深度增加而降低,其中占全磷比例有先升高后逐渐降低的趋势(表1)。酸模植被下0 ~ 13 cm土层土壤闭蓄态磷显著高于芦苇和苔草植被,而3种植被下13 ~ 55 cm土层土壤闭蓄态磷差异不显著,总体上除了酸模0 ~ 6 cm土层外,3种植被群落下土壤蓄态磷有着随着剖面深度增加而升高的趋势。3种植被群落下湿地剖面相同深度土壤钙磷存在显著差异,但总体上均存在随着土壤剖面深度增加,钙磷含量增加的现象,占全磷比例也存在相同趋势。4种无机磷组分差异在3种植被群落下土壤剖面0 ~ 25 cm差异性表现更为显著,而25 ~ 55 cm差异相对小些。
2.3 研究区湿地土壤剖面磷素形态间的相关性分析
湿地土壤磷素生物地球化学过程中,各个土壤磷形态组分间存在相互转化过程,但这些转化又深受土壤环境影响[3,20-21,23],因此,湿地土壤各个组分间及其与土壤理化性质存在一定相关性。研究区土壤铝磷含量与土壤全磷、有效磷、有机磷、无机磷、铁磷、闭蓄态磷及有机质含量均存在极显著正相关关系(表2);铁磷含量与土壤有机磷、有效磷及有机质含量存在极显著正相关关系,与全磷、无机磷、闭蓄态磷含量存在显著正相关关系;闭蓄态磷含量与土壤全磷、无机磷、有效磷含量及土壤pH存在极显著正相关关系,与有机磷含量存在显著正相关关系;钙磷含量与土壤黏粒含量、pH存在极显著正相关关系,与有机磷含量、容重存在显著正相关关系,与有机质含量存在显著负相关关系;无机磷含量与全磷含量、pH存在极显著正相关关系,与有机磷、有效磷含量存在显著正相关关系;有机磷含量与全磷、有效磷、有机质、黏粒含量存在极显著正相关关系,而与容重存在极显著负相关关系。
3 讨论
土壤无机磷是湿地植物所需磷的主要来源,一般在酸性土壤无机磷中铝磷和铁磷是植物有效磷源,闭蓄态磷是潜在有效磷源,而钙磷利用率很低[3,21]。而有些研究表明酸性土壤环境下,钙磷也是植物吸收有效磷的重要来源之一[12,23-25]。在研究区样地4种无机磷形态中,土壤钙磷含量最高,占土壤无机磷比重最大,与无机磷含量呈显著正相关关系(表2),同时3种植被下土壤剖面变化趋势与无机磷基本一致。因此,钙磷主导研究区无机磷变化,而铁磷含量较高,占有次等地位。前人研究表明湿地利用方式、植被群落及生长期、干湿交替、季节变化、土壤母质和季节性洪水等均影响湿地土壤磷素组分及含量[3-5,7-9,12-16]。本研究区处于一个退耕圩区,圩区建于1958年,一直种植水稻,1986年破圩后形成自然湿地,随后进行渔业养殖至今[18]。3种植被群落样区相邻,除了植被群落不同外,其他水文条件、土壤母质、利用方式均一致。因此,植被群落是影响湿地土壤磷素形态及其在土壤剖面上分布差异的主要因素。
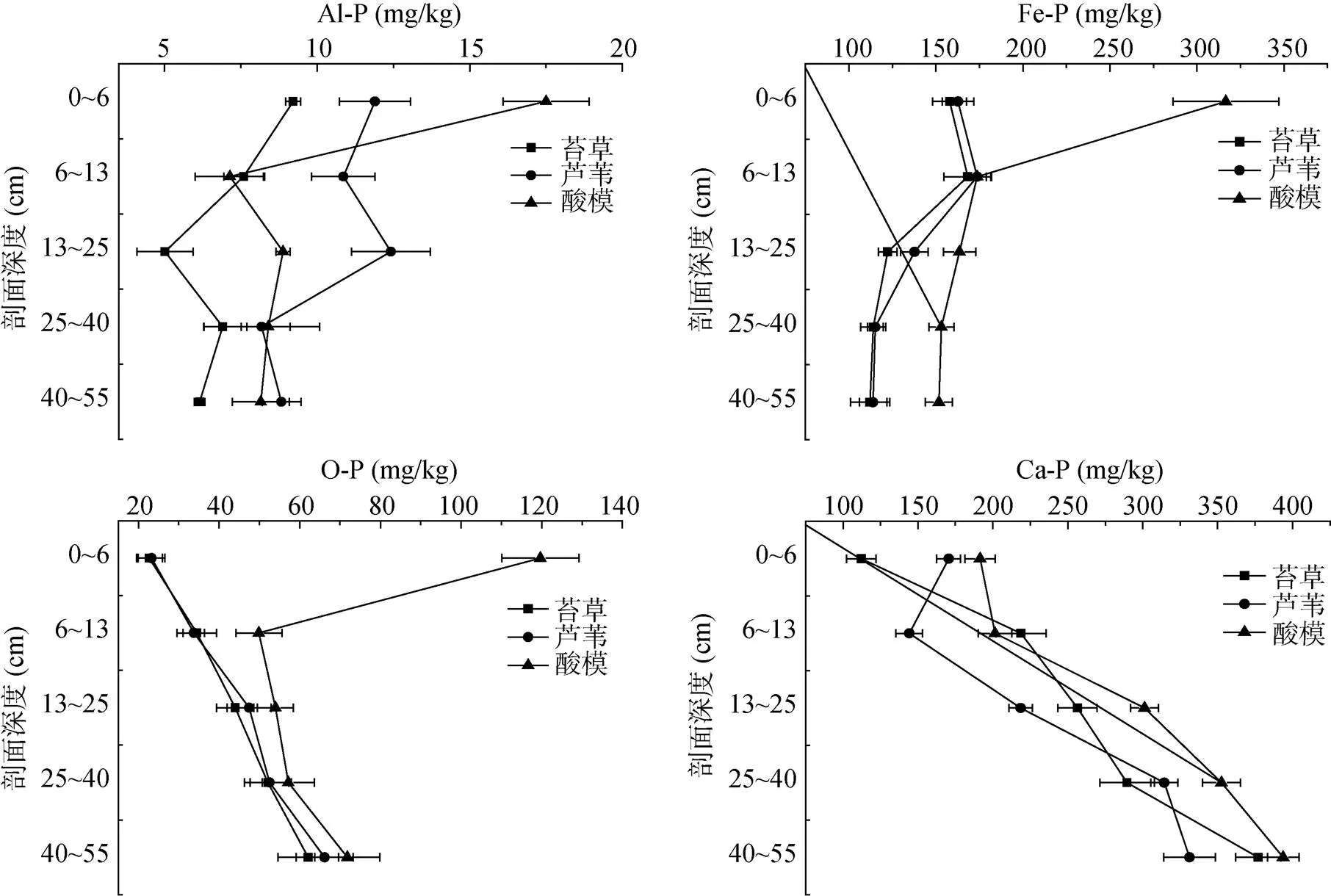
图3 不同植被群落湿地土壤剖面Al-P、Fe-P、O-P和Ca-P分布特征
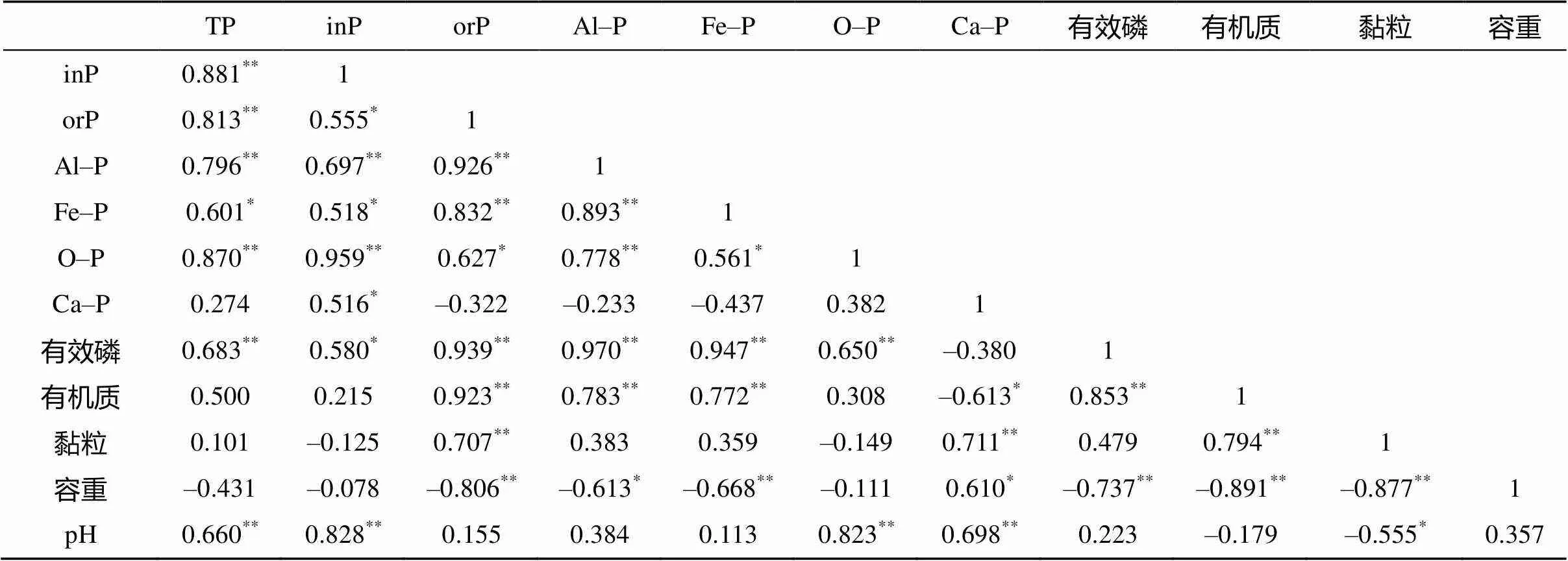
表2 研究区湿地土壤磷素各形态组分间及与土壤理化性状间的相关系数
注:=15;*表示在<0.05水平显著相关;**表示在<0.01水平极显著相关。
湿地土壤无机磷形态变化可受植物生长节律影响[8,13],不同湿地植物吸收土壤有效磷来源的磷形态和吸收最佳时期存在差异[14-15]。植被根系通过分泌物影响调控磷循环相关微生物群落及活性,促进土壤磷素转化[13],同时根系分泌的有机酸可活化土壤中的铝磷、铁磷和钙磷,其中低分子有机酸对铝磷和铁磷活化效率最高[23-24],且可将有效性低的钙磷形态大量转化为其他形态无机磷以供自身利用[10,24-25],而有机酸对闭蓄态磷活化最小[24]。同时,湿地水文条件也是磷形态转化的重要驱动因子[3,8],影响湿地土壤无机磷形态分布[9]。淹水条件促进湿地土壤铁磷中铁还原成氧化亚铁、晶形磷酸铁转化为无定形态,使磷被活化释放,为植物吸收利用[3,8],也可促使植被较难利用的土壤闭蓄态磷因氧化铁胶膜的还原而活化,在落干氧化时转为铁磷[26],进而促进磷素生物有效性提高。研究区苔草和酸模群落绝大多数根系分布在0 ~ 13 cm土层,13 ~ 55 cm土层根系较少,而芦苇根系在25 cm以下分布也较少,越接近表层土壤3种植被根系越多,磷素活化率也越高,有效磷含量也越高[18]。因此,退耕后淹水水文条件和3种植物通过根系分泌物不仅促进了铁磷与铝磷活化,提高表层土壤磷素生物有效性[18],本研究表明研究区铁磷、铝磷含量与有效磷含量均存在极显著正相关关系(表2);同时也促进了表层土壤剖面钙磷和闭蓄态磷占全磷比例下降,而铁磷和铝磷占全磷比例上升(表1),表明研究区表层土壤钙磷和闭蓄态磷向铁磷和铝磷转化。
植物通过对有效磷的吸收,还可促使磷素养分向表层迁移和积累[27-28]。众多研究表明,土壤磷素养分迁移、转化的主要存在形态均以有效性较低的钙磷、闭蓄态磷、铁磷为主[27],其中磷迁移和转化与土壤钙磷活性增加密切相关[12,25,27-29]。因此,研究区3种植被群落下均表现出表层土壤铝磷和铁磷相对富集,土壤钙磷相对消耗的现象,且土壤剖面钙磷与铁磷和铝磷存有不显著的负相关关系(表2)。不同植物根系通过分泌不同类型和数量的有机酸[30],而不同有机酸对不同无机磷形态活化存在差异[24]。同时不同植物群落下土壤剖面根系分布存在差异,研究区芦苇相对于苔草和酸模分布更深的根系[18],芦苇样地0 ~ 25 cm剖面土壤钙磷占全磷比例下降显著,土壤铁磷和铝磷占全磷比例上升显著;而钙磷、铁磷和铝磷相同变化趋势出现在苔草和酸模0 ~ 13 cm土壤剖面上。综上,植物根系是土壤剖面磷形态转化驱动力,而土壤钙磷活化是研究区其他磷形态表聚的重要磷源。
土壤有机磷的多少取决于土壤有机质数量及有机质分解速率[3,23],湿地土壤有机磷的磷素有效性较低,需经微生物分解矿化提高有效性[3,20]。研究区湿地土壤有机磷含量与土壤有机质含量呈极显著正相关(表2)。3种植被通过对无机磷吸收、植被凋落物及根系残体在表层土壤分解,积累土壤有机磷。湿地有机磷积累条件是微生物对有机磷的分解速率小于有机磷的积累速率[8],而淹水水文条件降低了微生物分解速率。同时研究区土壤有机磷含量与土壤黏粒含量呈极显著正相关(表2),表明黏粒也是影响土壤有机磷的重要因素。黏粒具有较大的比表面积,表面上易附着生物碎屑等有机物质。此外,黏土胶体可以保护有机质免受微生物的分解,进而促进对有机磷的保护。因此,苔草和酸模群落下0 ~ 6 cm土层和芦苇群落下0 ~ 13 cm土层由于凋落物和根系残体等富集,促使土壤有机磷所占全磷比例显著增加,而易被植物吸收利用的无机磷占全磷比例显著下降(表1)。不同湿地植被根系生物量和凋落物量及它们分解速率也存在差异[31-32]。酸模群落下土壤有机磷表聚最为显著,这可能是因为酸模群落凋落物中含有较高的多种维生素及氨基酸,但粗纤维含量较少,有利于微生物对凋落物的分解和转化,促进有机质较快积累,而苔草和芦苇干物质中所含粗纤维较高,导致枯落物分解速率较慢有关[18]。
研究区3种植被群落下均存在土壤全磷含量0 ~ 6 cm土层较高,除酸模群落样地外,显著低于40 ~ 45 cm土层,而6 ~ 13 cm土层急剧下降,13 ~ 55 cm土层土壤全磷含量随着深度增加而逐渐升高[18],表明退耕还湖后,湿地生态恢复过程中土壤磷素存在流失趋势。总体上,3种植被群落下土壤剖面的不同形态磷分布特征基本一致,除了含量较低的铝磷外,土壤铁磷、钙磷、闭蓄态磷、有机磷、无机磷和全磷均在相同深度剖面上酸模群落样地大于或等于芦苇和苔草群落样地,表明在当前湿地利用背景下,酸模群落相对于芦苇和苔草群落更有利于研究区湿地土壤磷素养分保持。
4 结论
1) 研究区湿地土壤磷素以无机磷为主,有机磷为辅;无机磷形态以钙磷为主,占无机磷总量的29.67% ~ 67.58%,且3种植被群落下土壤剖面分布特征与无机磷基本一致,主导着无机磷剖面分布特征。
2) 3种植被群落下湿地土壤铝磷、铁磷和有机磷都存在表聚现象,但不同植被群落表聚程度存在差异,酸模群落下湿地土壤这些磷素组分表现最为明显,且存在闭蓄态磷和无机磷表层富集。土壤钙磷在淹水和植被作用下活化,是提供表聚各种形态磷源的主要贡献者。研究区植物对土壤磷吸收利用是这些形态磷素转化和表聚的驱动力。
3) 相对于芦苇和苔草植被,研究区酸模植被更有利于退耕还湖后湿地生态恢复的土壤磷素养分保持。
[1] Mitsch W J, Gosslink J G. Wetlands[M]. New York: John Wiley and Sons. 2000
[2] 张仲胜, 吕宪国, 薛振山, 等. 中国湿地土壤碳氮磷生态化学计量学特征研究[J]. 土壤学报, 2016, 53(5): 1160– 1169
[3] 王国平. 湿地磷的生物地球化学特征[J]. 水土保持学报, 2004, 18(4): 193–199
[4] 章文龙, 曾从盛, 仝川, 等. 闽江口芦苇和短叶茳芏沼泽土壤磷分级特征比较[J]. 湿地科学, 2014, 12(6): 683– 689
[5] 张彬, 方芳, 陈猷鹏, 等. 三峡水库消落区土壤理化特征及磷赋存形态研究[J]. 环境科学学报, 2012, 32(3): 713–720
[6] SanClementsa M D, Fernandez I J, Norton S A. Soil and sediment phosphorus fractions in a forested watershed at Acadia National Park, ME, USA[J]. Forest Ecology and Management, 2009, 258: 2318–2325
[7] 刘文静, 张平究, 董国政, 等. 不同退耕年限下菜子湖湿地土壤磷素组分特征变化[J]. 生态学报, 2014, 34(10): 2654–2662
[8] 秦胜金, 刘景双, 王国平, 等. 三江平原湿地土壤磷形态转化动态[J]. 生态学报, 2007, 27(9): 3844–3851
[9] 陈峰峰, 李秋华, 焦树林, 等. 百花湖入库河流麦西河河口消落带土壤磷形态及其分布特征研究[J]. 长江流域资源与环境, 2013, 22(4): 486–492
[10] 卢同平, 张文翔, 武梦娟, 等. 干湿度梯度及植物生活型对土壤氮磷空间特征的影响[J]. 土壤, 2017, 49(2): 364–370
[11] 卢同平, 张文翔, 牛洁, 等. 典型自然带土壤氮磷化学计量空间分异特征及其驱动因素研究[J]. 土壤学报, 2017, 54(3): 682–692
[12] 梁威, 邵学新, 吴明, 等. 杭州湾滨海湿地不同植被类型沉积物磷形态变化特征[J]. 生态学报, 2012, 32(16): 5025–5033
[13] 孙波, 廖红, 苏彦华, 等. 土壤-根系-微生物系统中影响氮磷利用的一些关键协同机制的研究进展[J]. 土壤, 2015, 47(2): 210–219
[14] 章文龙, 曾从盛, 陈晓艳, 等. 闽江河河口湿地土壤有效磷时空分布与来源[J]. 生态学杂志, 2015, 34(1): 168– 174
[15] 刘长娥, 杨永兴, 杨杨. 九段沙中沙湿地植物磷元素的分布积累与动态[J]. 同济大学学报, 2008, 36(11): 1537– 1541
[16] 杜云鸿, 徐振, 谢文霞, 等. 胶州湾盐沼不同植被群落下土壤磷的分布特征[J]. 湿地科学, 2016, 14(3): 415– 420
[17] 宋晓琳, 吕宪国, 陈志科. 不同覆被条件下双台子河口湿地土壤主要营养元素含量[J]. 生态学杂志, 2010, 29(11): 2117–2121
[18] 崔宏, 张平究, 孔令柱, 等. 退耕还湖后不同植被群落下湿地土壤有机质及磷素含量变化[J]. 水土保持通报, 2013, 33(6): 94–98
[19] Chang S, Jackson M L. Fractionation of soil phosphorus[J]. Soil Science, 1957, 84(2): 133–144
[20] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 1999
[21] 屈凡柱, 于君宝, 陈小兵, 等. 湿地土壤磷分级方法研究[J]. 土壤通报, 2012, 43(1): 243–248
[22] 白军红, 余国营, 张玉霞. 向海湿地土壤中无机磷酸盐的存在形态研究[J]. 水土保持学报, 2001, 15(1): 98–101
[23] 向万胜, 黄敏, 李学垣. 土壤磷素的化学组分及其植物有效性[J]. 植物营养与肥料学, 2004, 10(6): 663–670
[24] 孔涛,伏虹旭,吕刚,等. 低分子量有机酸对滨海盐碱土壤磷的活化作用[J].环境化学,2016, 35( 7): 1526–1531
[25] 刘晶晶, 李敏, 曲博, 等. 湿地挺水植物根系土壤中的磷形态变化与分析[J]. 中国环境科学, 2013, 33(11): 2040– 2046
[26] 马利民, 张明, 滕衍行. 三峡库区消落区周期性干湿交替环境对土壤磷释放的影响[J]. 环境科学, 2008, 29(4): 1035–1039
[27] 余海英, 李廷轩, 张树金, 等. 温室栽培条件下土壤无机磷组分的累积、迁移特征[J]. 中国农业科学, 2011, 44(5): 956–962
[28] 吴艳宏, 周俊, 邴海健. 贡嘎山冷杉林草海子小流域土壤及湖泊沉积物中磷的形态及迁移特征[J]. 地球环境学报, 2013, 4(1): 1208–1214
[29] 应维南, 孙家干. 海积母质水稻土发育过程中磷的迁移[J]. 土壤通报, 1987, 18(5): 221–222
[30] 陈凯, 马敬, 曹一平, 等. 磷亏缺下不同植物根系有机酸的分泌[J]. 中国农业大学学报, 1999, 4(3): 58–62
[31] 郭绪虎, 田昆, 肖德荣, 等. 滇西北高原纳帕海湿地湖滨带优势植物凋落物分解特征研究[J]. 生态科学, 2013, 1999, 32(2): 200–205
[32] 赵建刚, 杨琼, 陈章和, 等. 几种湿地植物根系生物量研究[J]. 中国环境科学, 2003, 23(3): 290–294
Distribution of Phosphorus Fractions in Wetland Soil Profiles Under Different Vegetation Communities After Returning Farmland to Lake
BAO Xianming1, CUI Hong2,3
(1 College of Life Science, Huaibei Normal University, Huaibei, Anhui 235000, China;Linquan No.2 High School, Linquan, Anhui 236400, China; 3 Anhui Key Laboratory of Natural Disasters Process and Prevention, Wuhu, Anhui 241000, China)
Soils from the wetlands under different vegetation communities (,,Linn) were sampled to investigate the distribution of organic P, inorganic P and inorganic P forms in soil profiles. The results showed that inorganic P contents were ranged from 301.94 to 645.17 mg/kg, accounting for 52.59%–84.64% of total P in all soil profiles, while organic P contents were ranged from 78.40 to 254.27 mg/kg except for 0–6 cm layers underLinn. Ca-P and Fe-P dominated inorganic P in all soil profiles, accounting for 29.67%–67.58% and 21.9%–52.29% of inorganic P, respectively. Except for 0–6 cm layers underLinn, inorganic P contents decreased while organic P decreased seriously first and then increased with depth of soil profiles under all vegetation types. Except for O-P in 0–6 cm layers underLinn, Fe-P and Al-P contents decreased while Ca-P and O-P contents increased with depth of soil profiles. Al-P, Fe-P and organic P accumulated in surface layers under all vegetation types, especially underLinn. Plant uptake drives the form transformation and surface accumulation of phosphorus, while Ca-P is the main source of surface accumulated phosphorus.
Vegetation community; Wetland soil; Soil profile; Phosphorus form
国家自然科学基金项目(41001369)和国家水体污染控制与治理科技重大专项(2012ZX07103-005)资助。
包先明(1978—),男,安徽青阳人,博士,副教授,主要从事湿地沉积物营养元素地球化学循环研究。E-mail: xmb-7896@sohu.com
S153
A
10.13758/j.cnki.tr.2019.01.004
猜你喜欢
新农业(2022年17期)2022-09-15
河北科技师范学院学报(2022年2期)2022-08-26
中国农学通报(2022年21期)2022-08-22
农业工程学报(2022年5期)2022-06-22
食品安全导刊(2021年21期)2021-08-30
食品安全导刊(2021年20期)2021-08-30
石材(2020年9期)2021-01-07
农药科学与管理(2019年7期)2019-11-29
建材发展导向(2019年10期)2019-08-24
中学生数理化(高中版.高考理化)(2019年6期)2019-06-22
