
基质浓度对分体式厌氧氨氧化工艺固碳能力的影响
2019-04-04 03:03:34马雪研项邦东张文杰
安全与环境工程 2019年2期
马雪研,项邦东,张文杰
( 1.桂林理工大学广西环境污染控制理论与技术重点实验室,广西 桂林 541004;2.桂林理工大学广西岩溶地区水污染控制与用水安全保障协同创新中心,广西 桂林 541004 )
传统污水处理厂所采用的生物脱氮处理工艺不仅耗能高,需要添加有机碳源,还会产生较多的CO2气体,造成温室效应,并且很难用于处理低碳氮比的废水[1]。近年来,厌氧氨氧化工艺由于其成本效益高、无需外加碳源等优点[2],已经逐渐应用于处理高氨氮废水[3-4]。

(1)
厌氧氨氧化工艺的关键微生物——亚硝化细菌和厌氧氨氧化细菌均为化能自养型微生物,具有一定的固碳能力[10],但厌氧氨氧化菌会受到许多因素的影响使其活性降低,如温度、溶解氧(DO)、pH值、基质浓度、水力停留时间、亚硝酸盐抑制作用等[11-12]。Chouten等[13]通过同位素示踪法和酶活法推断其固碳方式为Calvin cycle或acetyl CoA途径。据日本Hitachi Plant Technologies Ltd研究组的报道,厌氧氨氧化工艺的固碳能力不仅受到细菌新陈代谢的影响,还与生化反应的过程等有关[14]。因此,针对厌氧氨氧化工艺的固碳潜力和机理进行研究,对促进微生物增殖的同时兼顾温室气体的减排,开发高固碳能力的厌氧氨氧化工艺具有重要的现实意义。
氨与亚硝酸盐作为厌氧氨氧化反应的底物,若超过一定的浓度范围,就会成为厌氧氨氧化细菌的抑制剂[15]。本试验通过改变反应器进水的基质浓度,研究其对短程硝化与厌氧氨氧化耦合工艺短期运行、固碳能力的影响以及在最佳固碳效果时的参数值,可为分体式厌氧氨氧化工艺的实际应用提供理论依据。
1 材料与方法
1.1 试验装置
本试验采用的分体式厌氧氨氧化试验装置如图1所示。试验所有反应器装置均由有机玻璃加工而成,其中R1短程硝化反应器为本研究中的前置工艺,调节pH值在7.55~7.60之间,使用温度控制系统将反应器温度控制在(30±1)℃之间,为亚硝化菌提供最适宜的生长条件;R2为沉淀池,用于回流和沉淀,R1出水溢流至内圆柱后,污泥通过重力沉降回流,上清液从外圆柱上的小孔流出;R3为中转池,向其中添加亚硫酸钠浓溶液起脱氧作用;R4为上流式折流板厌氧氨氧化反应器,由主体反应器和调节槽构成,主体反应器外侧用黑布包裹,防止光线对厌氧氨氧化菌代谢产生不利影响[16]。试验通过控制反应器内温度在(30±1)℃之间,保证厌氧氨氧化细菌的活性,温度通过设置在反应器外部的保温夹层控制,由温控系统为反应器持续提供保温循环水;循环水从反应器底部进入夹层,再从反应器顶部返回温控系统。
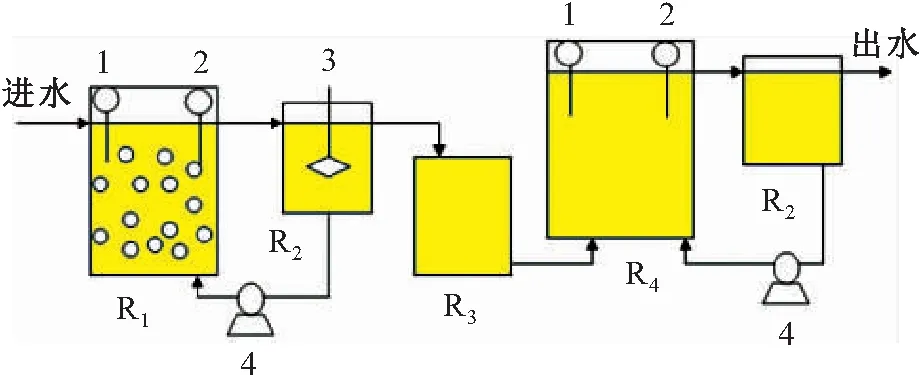
图1 分体式厌氧氨氧化试验装置原理图Fig.1 Schematic of experimental flow chartR1.短程硝化反应器;R2.沉淀池;R3.中转池;R4.厌氧氨氧化反应器;1.pH在线监测仪;2.溶解氧在线监测仪;3.搅拌器;4.污泥回流泵
1.2 接种污泥和试验用水
本试验中R1、R4反应器中污泥取自实验室驯化成功的短程硝化污泥和厌氧氨氧化污泥[17-18],接种污泥数量分别为1 L、1.5 L,接种污泥形态如图2和图3所示。

图2 短程硝化污泥Fig.2 Partial nitrification sludge

图3 厌氧氨氧化污泥Fig.3 Anammox sludge

阶段NH4HCO3KH2PO4NaHCO3CaCl2·2H2OMgSO4·7H2O微生物促进剂(瑞成环保提供)第一阶段85501 0001001000.5第二阶段14550按需配制1001000.5第三阶段18050按需配制1001000.5第四阶段24050按需配制1001000.5
注:人工模拟废水中除微生物促进剂的单位为mL/L外,其余组分的单位均为mg/L。
1.3 分析项目和方法
1.3.1 常规水质指标分析
试验中,确保出水水质稳定后开始采集水样。采集的水样经过0.22 μm滤膜后置于取样瓶中,取样后立即检测或者放置于4℃冰箱保存后统一检测。水样中常规水质指标及其测定方法见表2。

表2 常规水质分析项目和测定方法
*注:德国耶拿总有机碳分析仪(multi N/C UV3100)的工作原理是:首先测试样品的总碳,其次测试样品的总无机碳,最后得到样品的总有机碳,该过程中包括无机碳(IC)这一指标,因此选用该种测定方法。

(2)
固碳量=(进水IC-出水IC-单位时间内产生的CO2密度)÷单位时间进入反应器中的氮素量
(3)
式中:IC为水样中以各种形式存在的无机碳;CO2密度统一采用0℃、101 kPa条件下的数值,其值约为1.977 g/L。
1.3.2 CO2测试分析
本试验采用上海精科GC-112A型气相色谱仪对收集的气体成分进行分析,该仪器使用浙江大学智达信息工程有限公司研发的N(VI)2000色谱数据工作站。CO2标准气体由上海神开气体有限公司提供,标准气体CO2浓度为4.92%,试验操作条件见图4。
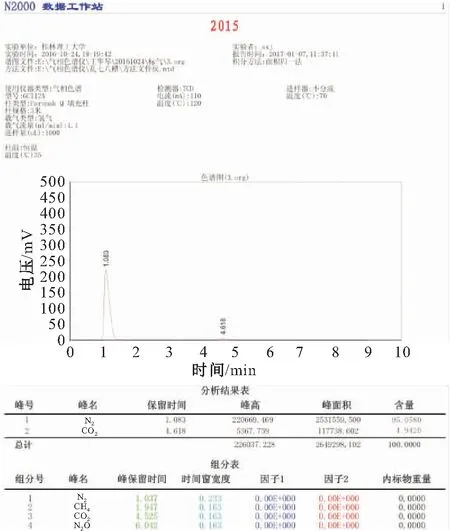
图4 CO2标准气体色谱图及分析结果Fig.4 Standard gas chromatography of CO2 and analysis results
1.4 微生物种群多样性分析
本试验中所采用的污泥来自于基质试验中4个阶段前后两部分的污泥,一共8个样品,采样离心后统一保存于-20℃冰箱中。采用PCR-DGGE分子生物学技术来研究反应器中微生物群落结构的变化,并借此确认分体式厌氧氨氧化工艺中起关键作用的功能微生物。此项试验一共分为三个步骤,分别为核酸提取、PCR扩增和DGGE技术,统一在微生物实验室中完成,测序则委托上海生物工程有限公司完成。
2 结果与分析
2.1 短程硝化阶段
2.1.1 短程硝化阶段的脱氮性能分析
分体式厌氧氨氧化工艺中短程硝化阶段进出水氮素浓度的变化,见图5。反应器中接种已经驯化成功的亚硝化污泥,利用控制DO浓度的方法来控制亚硝化过程的稳定运行,4个阶段的曝气量分别为0.10 L/min、0.18~0.20 L/min、0.38~0.40 L/min和0.50 L/min。
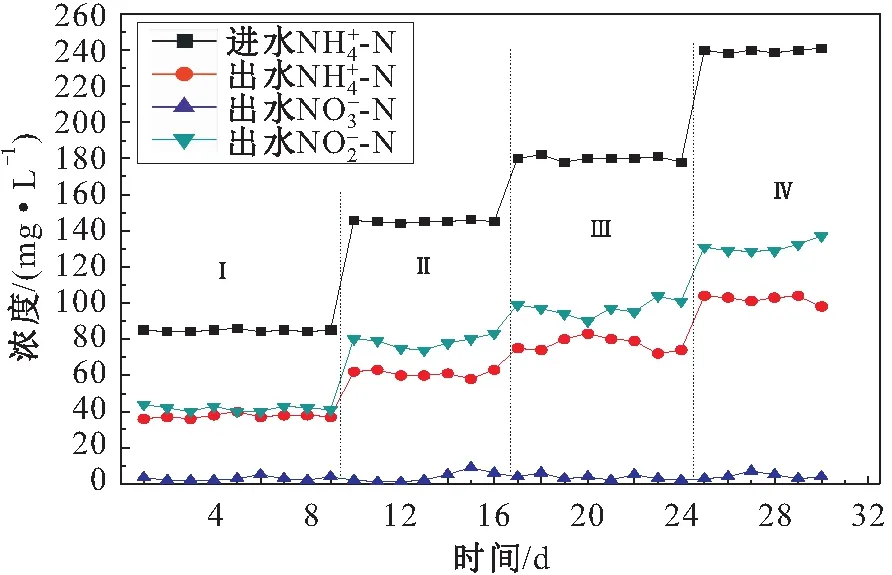
图5 短程硝化阶段进出水氮素浓度浓度的变化Fig.5 Nitrogen relationship between influent and effluent in partial nitrification stage
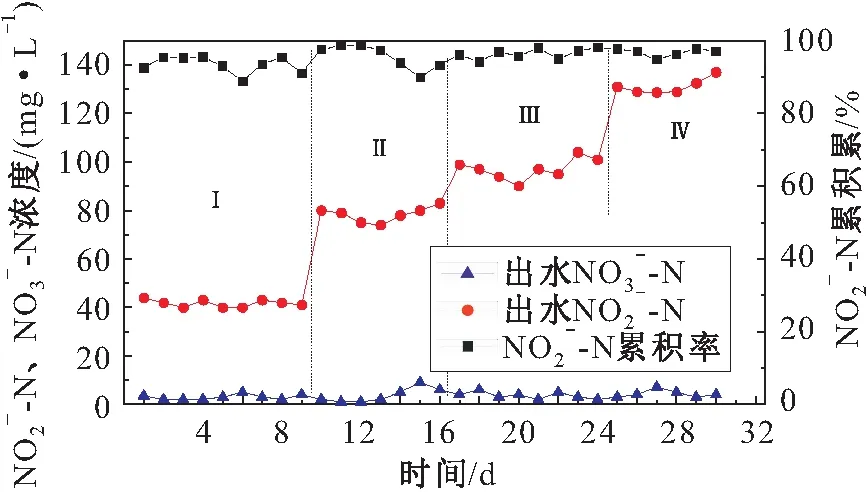
图6 短程硝化阶段N-N的累积率Fig.6 N-N accumulation rate in partial nitrification stage
2.1.2 短程硝化阶段的固碳能力分析
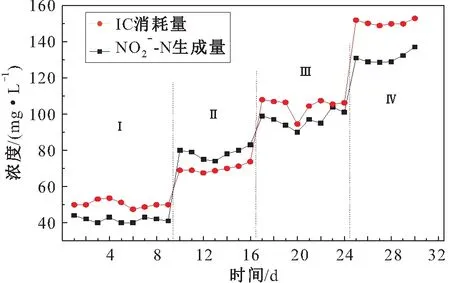
图7 短程硝化阶段IC消耗量与N-N生成量的 关系Fig.7 Relationship between IC consumption and N-N production in partial nitrification stage
y=0.847x+7.074 (R2=0.937,Sig.=0.00)
其中,R2=0.937说明方程拟合度很高,在Anova中符合F校验;Sig.<0.05,说明显著性良好,表明两者之间呈正相关性。
短程硝化反应式如下:
ΔG=-275 kJ/mol
(4)
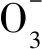
图8 短程硝化阶段CO2释放量与N-N生成 量的关系Fig.8 Relationship between CO2 emission and N-N production in partial nitrification stage
y=1.694x-11.884 (R2=0.976,Sig.=0.00)
其中,R2=0.976说明方程拟合度很高,在Anova中符合F校验;Sig.<0.05,说明显著性良好,表明两者之间呈正相关性。
2.2 厌氧氨氧化阶段
2.2.1 厌氧氨氧化阶段的脱氮性能分析
分体式厌氧氨氧化工艺中厌氧氨氧化阶段进出水氮素浓度的变化,见图9。

图9 厌氧氨氧化阶段进出水氮素的关系Fig.9 Nitrogen relationship between influent and effluent in Anammox stage
为了证实上述假设,本试验在第四阶段后另增设一组试验,即在进水中增加NaHCO3的量,但处理效果并未得到明显的改善,故可以排除进水IC值偏低的推测。
根据厌氧氨氧化反应的机理,在纯菌条件下,以最佳比例进水的反应器脱氮率约为88.8%。厌氧氨氧化阶段总氮去除率的变化,见图10。
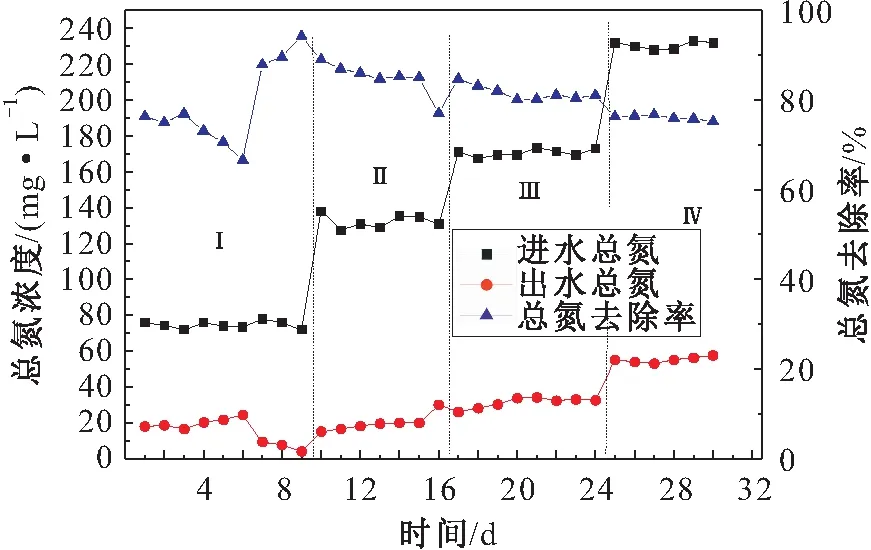
图10 厌氧氨氧化阶段总氮去除率的变化Fig.10 Removal rate of total nitrogen in Anammox stage
由图10可见,在第一阶段中,总氮的去除率较低,这是由于负荷太低会抑制厌氧氨氧化细菌的活性,导致处理效果较差且不稳定;在第二和第三阶段中,总氮的去除率均高于80%,表明脱氮效率较高;在第四阶段中,由于厌氧氨氧化污泥的浓度不足,导致总氮的去除率降低。
2.2.2 厌氧氨氧化阶段的固碳能力分析
迄今为止,已发现的无机碳固定途径主要有Calvin循环、厌氧乙酰辅酶A、还原性三羧酸循环、3-羟基丙酸和琥珀酰辅酶A 5种途径[22]。其中,厌氧乙酰辅酶A途径是近年来在严格厌氧细菌中新发现的自养微生物的无机碳固定途径。图11为厌氧氨氧化阶段中IC消耗量与脱氮量的关系。
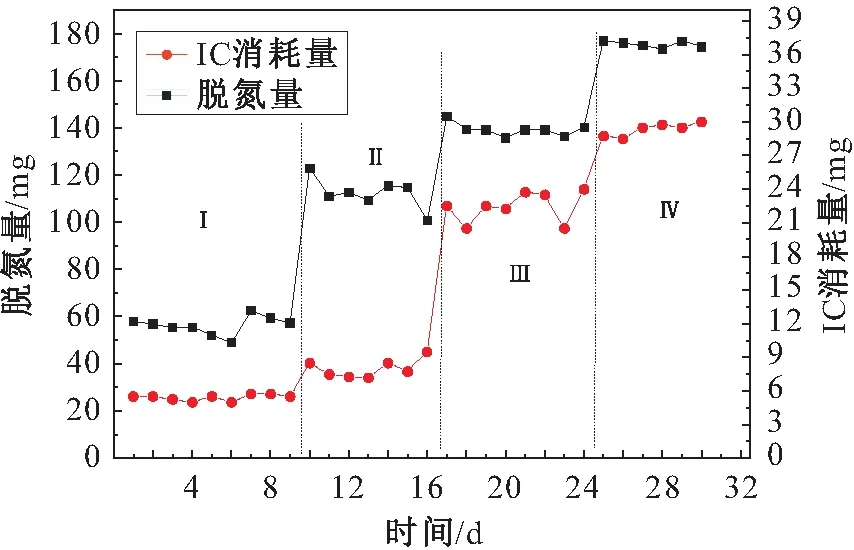
图11 厌氧氨氧化阶段IC消耗量与脱氮量的关系Fig.11 Relationship between IC consumption and denitrification in Anammox stage
由图11可见,厌氧氨氧化反应有较高的固碳能力,随着脱氮量的增加,厌氧氨氧化反应阶段中IC的消耗量也逐渐增大。
以脱氮量为自变量x,IC消耗量为因变量y,利用SPSS软件进行线性回归分析,可以得出如下线性拟合方程:
y=0.204x-8.122 (R2=0.842,sig=0.00)
其中,R2=0.842说明方程拟合度良好,在Anova中符合F校验;Sig.<0.05,说明显著性良好,表明两者之间呈正相关性。
y=0.271a+0.089b-9.397 (R2=0.890,Sig.=0.00)
由图12可见,除第一阶段外,CO2释放量都与脱氮量呈现相同的变化趋势。第一阶段CO2释放量较高,可能是因为在低负荷下厌氧氨氧化细菌的活性会受到抑制甚至会出现细菌解体的现象,导致产生较多的CO2;而随后的三个阶段中,CO2释放量的变化趋势都较为稳定。去掉本试验中第一阶段的数据后,以脱氮量为自变量x,CO2释放量为因变量y,利用SPSS软件进行线性回归分析,可以得出如下线性拟合方程:
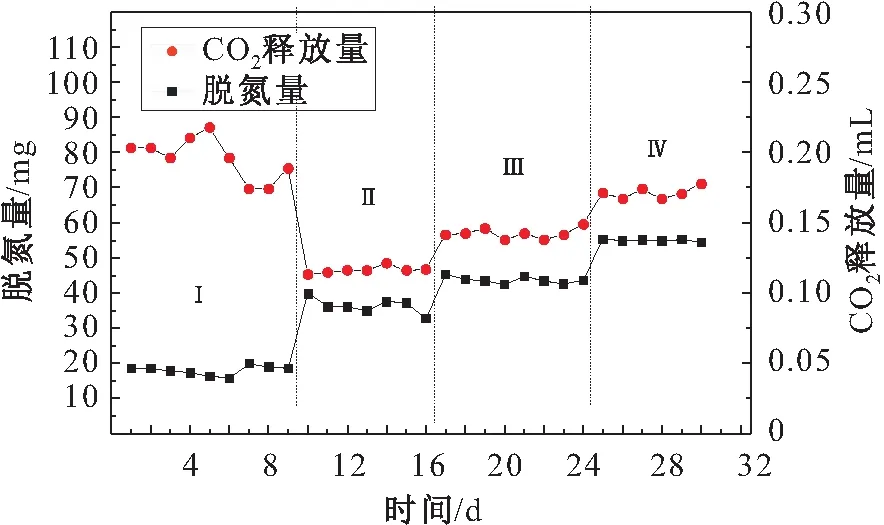
图12 厌氧氨氧化阶段CO2释放量与脱氮量的关系Fig.12 Relationship between CO2 emission and denitrification in Anammox stage
y=0.003x+0.016 (R2=0.936,Sig.=0.00)
其中,R2=0.936说明方程拟合度很好,在ANOVA中符合F校验;Sig.<0.05,说明显著性良好,表明两者之间呈正相关性。
2.3 微生物种群多样性分析
图13为本次试验中短程硝化阶段污泥系统发育树。由于经过长时间驯化,故比较短程硝化各个阶段污泥中的微生物种群,并没有较大区别,且各个阶段的条带强度变化不大。经过分析,在短程硝化阶段的污泥中并没有发现亚硝化单胞菌属,但是发现了与其同属于变形菌门(Proteobacteria)的微生物,即D16与D17。对比本试验组相同短程硝化阶段的微生物分析以及污泥系统发育树(见图14),结果发现两次微生物分析均没有得到亚硝化单胞菌属,但均可证明污泥中有类似D16、D17的变形菌门(Proteobacteria)的微生物作为优势菌种存在。结合反应器中的短程硝化反应,故推测D16、D17这两种微生物可能与亚硝化单胞菌属具有类似的功能。

图13 短程硝化阶段污泥系统发育树Fig.13 Phylogenetic tree of partial nitrification sludge

图14 试验组短程硝化阶段污泥系统发育树Fig.14 Phylogenetic tree of partial nitrification sludge in experimental group
图15为本试验中厌氧氨氧化阶段污泥系统发育树。

图15 厌氧氨氧化阶段污泥系统发育树Fig.15 Phylogenetic tree of Anammox sludge
由图15可见,厌氧氨氧化中污泥阶段的细菌主要为浮霉菌门(Planctomycetes)、放线菌门(Actinobacteria)、变形菌门(Proteobacteria)这三类。其中,浮霉菌门(Planctomycetes)是已知的厌氧氨氧化细菌,其形态通常为卵状,且随着试验的进行,代表浮霉菌门的D10条带强度也随之加强,故判断其为厌氧氨氧化阶段的优势菌种。
3 结 论
(3) 微生物种群多样性分析表明,短程硝化反应器中的优势菌种分别与AcidobacteriaBacteria(酸杆菌)、Chlorobi(绿菌)、ProteobacteriaBacteria(变形杆菌)具有99%以上的同源性;厌氧氨氧化反应器中的优势菌种分别与PlanctomyceteBacteria(浮霉菌)、ActinobacteriaBacteria(放线菌)、ProteobacteriaBacteria(变形杆菌)具有98%以上的同源性。
猜你喜欢
玩具世界(2022年2期)2022-06-15 07:35:58
选煤技术(2021年6期)2021-04-19 12:21:58
环境保护与循环经济(2017年7期)2018-01-22 02:58:12
工程建设与设计(2016年4期)2016-02-27 10:51:11
中国环境科学(2016年3期)2016-02-08 15:07:14
风能(2015年10期)2015-02-27 10:15:39
应用海洋学学报(2014年2期)2014-11-26 01:20:46
植物营养与肥料学报(2014年1期)2014-03-11 20:24:27
植物营养与肥料学报(2012年2期)2012-10-26 02:52:24
植物营养与肥料学报(2012年1期)2012-10-26 02:49:12
