
飞蝗内表皮结构糖蛋白基因LmAbd-2的表达与功能分析
2019-03-18 06:09贾盼张晶杨洋刘卫敏张建珍赵小明
中国农业科学 2019年4期
贾盼,张晶,杨洋,刘卫敏,张建珍,赵小明
飞蝗内表皮结构糖蛋白基因的表达与功能分析
贾盼1,2,张晶1,2,杨洋1,2,刘卫敏1,张建珍1,赵小明1
(1山西大学应用生物学研究所,太原 030006;2山西大学生命科学学院,太原 030006)
【目的】分析飞蝗()内表皮结构糖蛋白(endocuticle structural glycoprotein)基因的序列特征和表达特性,利用RNA干扰(RNA interference,RNAi)技术结合Hematoxylin and eosin(H&E)染色方法以及透射电镜(transmission electron microscopy,TEM)技术探究其生物学功能,探讨其对飞蝗内表皮发育的影响。【方法】基于课题组飞蝗转录组数据库获得的cDNA编码序列,并克隆验证。利用ExPASy工具翻译LmAbd-2氨基酸序列,然后利用SignaIP4.1Server和SMART软件分别对其信号肽和结构域进行分析;利用NetOGlyc4.0 Server对其糖基化位点进行预测,使用GENEDOC软件进行多序列比对,并利用MEGA 6.0软件中neighbor-joining(NJ)方法与其他昆虫同源序列进行聚类分析。采用荧光定量PCR(reverse- transcription quantitative PCR,RT-qPCR)分析在飞蝗5龄第2天不同组织以及4龄(N4D1—N4D6)、5龄(N5D1—N5D8)和成虫(AD1—AD4)表皮不同发育时期表达模式。采用RNAi技术结合H&E染色方法以及TEM技术观察干扰后对飞蝗生长发育和表皮结构的影响。【结果】序列分析结果显示,飞蝗cDNA编码序列长为683 bp,开放阅读框为459 bp,编码152个氨基酸;功能域分析发现LmAbd-2含有1个信号肽和1个几丁质结合域(chitin binding domain 4,ChtBD4),属于CPR家族RR-1亚类;序列比对分析发现其在物种间具有较高的保守性,其中与沙漠蝗()SgAbd-2序列相似度为92.5%,且在N末端含有一个保守基序PTPPPIP,其中第2位苏氨酸为糖基化位点;系统发育分析结果显示LmAbd-2与SgAbd-2聚为一支,亲缘关系最近;RT-qPCR分析结果显示在体壁中高表达,且在蜕皮后(内表皮形成时期)高表达;沉默后飞蝗没有肉眼可见的表型,但H&E染色结果显示与对照组相比飞蝗表皮层变薄,进一步超微结构分析发现内表皮片层结构减少。【结论】LmAbd-2是一种内表皮结构糖蛋白,属于CPR家族RR-1亚类,其编码基因主要在体壁中高表达,而且在飞蝗4龄、5龄和成虫内表皮形成时期呈周期性表达;RNAi试验表明,在飞蝗蜕皮过程中参与内表皮的形成。
飞蝗;内表皮;糖蛋白;;RNA干扰
0 引言
【研究意义】昆虫体壁主要由上表皮(epicuticle)、外表皮(exocuticle)、内表皮(endocuticle)和真皮细胞层(epidermal cell)组成[1],其在维持虫体外部形态、防止体内水分蒸发、阻止病菌入侵等方面具有重要作用[2]。昆虫在蜕皮过程中首先合成上表皮和外表皮(在蜕皮前合成),然后在蜕皮后逐渐合成内表皮[3]。表皮蛋白(cuticle protein,CP)是昆虫体壁的重要组成成分,其编码基因在昆虫基因组总基因中所占比例超过1%[4],在昆虫生长发育、表皮物理特性以及虫体形态构成中具有重要作用[1]。飞蝗()是一种渐变态世界性农业害虫,对RNA干扰(RNA interference,RNAi)具有敏感性,是基于RNAi研究基因功能的理想模型。因此,以飞蝗蜕皮后表皮蛋白基因为研究对象,探讨其分子特性、表达模式及生物学功能,可为蜕皮后表皮蛋白基因在渐变态昆虫蜕皮过程中参与内表皮形成的作用机制研究提供依据。【前人研究进展】目前,随着黑腹果蝇()、赤拟谷盗()、家蚕()、冈比亚按蚊()、褐飞虱()、烟草天蛾()等昆虫基因组测序的完成,越来越多的CPs或CP-like蛋白被鉴定[5-10]。根据所含基序不同,这些CPs被分为不同家族,如数目最多的CPR家族(包括RR-1、RR-2和RR-3 3个亚类)、CPAPs家族、Tweedle家族、CPF/CPFL家族、CPG家族以及CPH家族等。在双翅目黑腹果蝇、鞘翅目赤拟谷盗、鳞翅目家蚕、半翅目褐飞虱等昆虫中CPR家族表皮蛋白在表皮分化、鞘翅片层结构形成、翅原基发育以及虫体生长发育等方面具有重要作用[10-17]。同样,CPAPs家族表皮蛋白在昆虫体形和生长发育中具有重要作用[18-20]。目前,对其他家族表皮蛋白基因的功能研究也取得了重要进展,如甜菜夜蛾()CPG家族蛋白基因(、和)的缺失导致表皮黑化变硬和蜕皮致死[21];LU等干扰褐飞虱未分组表皮蛋白(不属于任何已分类家族)基因(主要表达于蜕皮后时期)可导致虫体发育异常或致死,并且影响内表皮的形 成[17];家蚕蜕皮后高表达表皮蛋白基因在维持虫体形态和内表皮的形成中具有重要作用[16]。在蝗虫中,20世纪90年代,丹麦科学家Andersen[22]通过双向电泳结合质谱分析方法从沙漠蝗()成虫腹部表皮中鉴定出了8个内表皮结构蛋白(endocuticle structural protein),分别命名为SgAbd-1、SgAbd-2、SgAbd-3、SgAbd-4、SgAbd-5、SgAbd-6、SgAbd-8、SgAbd-9。在飞蝗中,根据这8个内表皮结构蛋白氨基酸序列笔者课题组前期基于飞蝗转录组数据库获得了相应的同源蛋白序列[23],分别命名为LmAbd-1、LmAbd-2、LmAbd-3、LmAbd-4、LmAbd-5、LmAbd-6、LmAbd-8、LmAbd-9,它们均属于CPR家族RR-1亚类,并且研究发现主要表达于外胚层形成的组织中,可能参与飞蝗内表皮的形成[24]。然而,这些表皮蛋白基因在飞蝗蜕皮过程中的具体功能尚不清楚。【本研究切入点】目前,尽管表皮蛋白基因在模式昆虫生长发育中的功能研究取得了较大进展,但对于渐变态昆虫飞蝗表皮蛋白的研究主要集中于表皮蛋白的鉴定和分类,对其编码基因的表达模式及功能尚缺乏系统研究,尤其是蜕皮后表皮蛋白基因在内表皮结构形成中具有怎样的作用机制有待进一步研究。【拟解决的关键问题】以重要农业害虫——飞蝗为研究对象,基于转录组数据获得的内表皮蛋白基因,研究其分子特性、组织和时期表达特性,并基于RNAi方法结合H&E染色以及透射电镜技术分析其生物学功能,探讨其在飞蝗蜕皮过程中对内表皮结构形成的影响,为阐明蜕皮后表皮蛋白在内表皮形成中的作用机制提供依据。
1 材料与方法
试验于2017—2018年在山西大学应用生物学研究所完成。
1.1 供试昆虫与试剂
将飞蝗虫卵(购自河北沧州蝗虫养殖基地)包裹于已扎孔透明塑料袋中,置于昆虫恒温培养室内进行孵化,孵化后转移至纱笼(30 cm×30 cm×30 cm)内,控制昆虫培养室内环境:温度为(30±2)℃,相对湿度为(40±10)%,光周期为14 h﹕10 h(L﹕D),以新鲜小麦苗和麦麸饲喂。
试剂:动物组织RNA提取试剂(RNAisoTMPlus)、cDNA反转录试剂(Reverse Transcriptase M-MLV,5×Reverse Transcriptase M-MLV Buffer,Recombinant RNase Inhibitor,dNTP Mixture)、实时荧光定量试剂(TB GreenTMPremix Ex TaqTMⅡ)购自TaKaRa公司;dsRNA合成试剂盒(T7 RiboMAXTMExpress RNAi System)购自Promega公司;RNase Free H2O等其他试剂购自生工生物(上海)有限公司。
1.2 LmAbd-2 cDNA序列分析
根据飞蝗转录组的cDNA编码序列,克隆测序后,利用NCBI中的Open Reading Frame Finder(ORF Finder)(http://cms.ncbi.nlm.nih.gov/orf finder/)工具找到的全长ORF序列,利用ExPASy网站在线翻译工具翻译成氨基酸序列(https://web. expasy.org/translate/),并预测蛋白分子量大小及等电点(pI)(https://web.expasy.org/compute_pi/);利用SignaIP 4.1 Server(http://www.cbs.dtu.dk/services/ SignalP/)预测其信号肽,利用SMART(http://smart. embl-heidelberg.de/)分析其结构域,利用GENEDOC软件进行多序列比对,利用NetOGlyc4.0 Server预测其糖基化位点(http://www.cbs.dtu.dk/services/NetOGlyc/),利用WebLogo(http://weblogo.berkeley.edu/logo.cgi)分析其保守基序。
1.3 序列比对与系统发育分析
在NCBI网站下载不同昆虫物种Abd-2氨基酸序列,利用MEGA 6.0软件中neighbor-joining(NJ)方法将LmAbd-2氨基酸序列与沙漠蝗、德国小蠊()、赤拟谷盗、斜纹夜蛾()、褐飞虱等不同目物种的同源氨基酸序列进行聚类分析,独立分析1 000次,数值代表bootstrap估算值。不同目物种Abd-2序列和飞蝗LmAbd-2序列的GenBank登录号见表1。
1.4 LmAbd-2组织特异性和时期表达分析
组织特异性表达分析:解剖发育整齐的5龄第2天(N5D2)飞蝗若虫的体壁(integument,IN)、前肠(foregut,FG)、中肠(midgut,MG)、后肠(hindgut,HG)、胃盲囊(gastric caeca,GC)、马氏管(Malpighian tubules,MT)、脂肪体(fat body,FB)和翅芽(wing pad,WP)8个不同组织,设置4个生物学重复,每个重复3头虫体。时期表达分析:选取并解剖4龄第1—6天飞蝗若虫(N4D1—N4D6)、5龄第1—8天飞蝗若虫(N5D1—N5D8)和成虫第1—4天(AD1—AD4)第2腹节处体壁组织,设4个生物学重复,每3头虫体组织为一个生物学重复,组织样品冻存于液氮中。利用动物组织RNA提取试剂RNAisoTMPlus(TaKaRa公司)提取上述不同组织和不同发育时期体壁组织的总RNA,使用1%的琼脂糖凝胶电泳检测RNA的质量,利用Nanodrop 2000对总RNA进行定量。以1 µg总RNA为模板,使用cDNA反转录试剂(TaKaRa公司)合成cDNA模板。采用荧光定量PCR方法(reverse-transcription quantitative PCR,RT-qPCR)检测在5龄第2天飞蝗不同组织部位及不同发育时期体壁中的表达情况。RT-qPCR反应体系和程序参考文献[25]。(KX276642)作为内参基因,引物见表2(由上海生工生物公司合成)。
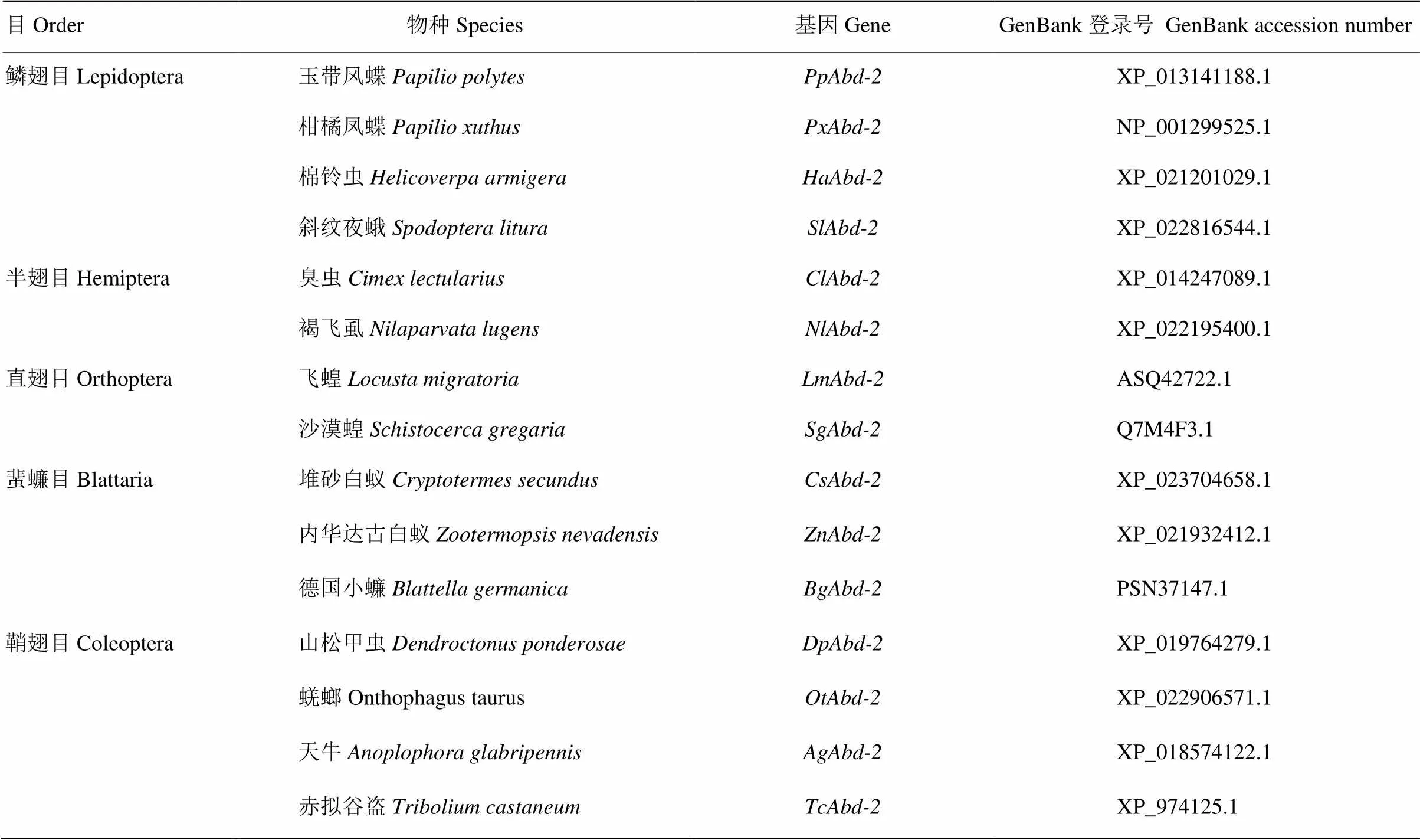
表1 本研究中用于构建系统进化树的物种和Abd-2的GenBank登录号

表2 本研究中所用引物
1.5 基于RNAi的LmAbd-2生物学功能分析
为了探讨在飞蝗表皮形成中的作用,利用T7 RiboMAXTMExpress试剂盒(Promega公司)体外合成该基因双链RNA(dsRNA),以绿色荧光蛋白基因()为对照。利用微量注射器向4龄第3天(N4D3)若虫第2—3腹节处注射10 μg ds和ds,正常饲养;若飞蝗正常蜕皮后,再向5龄第4天(N5D4)若虫注射10 μg的ds和ds,24 h后利用1.4中RT-qPCR方法检测沉默效率,其余若虫正常饲养、观察表型。
1.6 表皮显微和超微结构观察
为了观察干扰后对飞蝗表皮结构的影响,取5龄蜕皮后72 h的对照组和处理组腹部表皮样品(第3腹节)于3%戊二醛溶液中4℃固定过夜。固定的样品进行石蜡包埋、切片,然后用苏木精和伊红(hematoxylin-eosin staining,H&E)染色[26-27]、封片后利用显微镜(Olympus BX51,Japan)观察表皮结构。
为进一步从超微结构分析对飞蝗内表皮形成的影响,用透射电镜技术观察表皮结构变化。用于透射电镜观察的腹部表皮样品固定、切片制备和染色等方法参考文献[25]。
1.7 数据分析
基因相对表达量采用2-ΔΔCt法[28]进行分析,采用Student t-test方法进行差异表达分析,*表示在<0.05水平差异显著,**表示在<0.01水平差异显著,***表示在<0.001水平差异显著。
2 结果
2.1 LmAbd-2的序列结构特征
根据飞蝗转录组数据库获得cDNA编码序列并进行了克隆和测序验证,其编码序列长为689 bp,其中ORF为459 bp, 编码152个氨基酸,预测其分子量为16.98 kD,等电点为5.28,为偏酸性表皮蛋白。序列分析发现其含有1个信号肽和1个几丁质结合域(chitin binding domain 4,ChtBD4)(图1-A),并且具有RR-1保守基序,属于CPR家族RR-1亚类(图1-B)。

A:LmAbd-2的序列分析Sequence analysis of LmAbd-2;下划线为信号肽The underline: signal peptide,方框内为几丁质结合域The box: chitin binding domain 4, ChtBD4。B:Weblogo分析LmAbd-2保守基序Analysis of conserved motifs by Weblogo (http://weblogo.berkeley.edu/logo.cgi)
2.2 Abd-2序列比对与系统发育树分析
多序列比对分析发现LmAbd-2与沙漠蝗、褐飞虱、赤拟谷盗和棉铃虫()等Abd-2序列具有较高相似性,其中与沙漠蝗SgAbd-2的氨基酸序列相似度高达92.5%(信号肽除外),几丁质结合域序列完全一致(图2-A)。同时发现Abd-2 N末端存在一个保守基序PTPPPIP,且第2位苏氨酸(T)是O-连接糖基化位点(O-GalNAc)(图2-A),与SgAbd-2类似,是一个糖基化蛋白[22]。将获得的飞蝗Abd-2氨基酸序列与其他昆虫的Abd-2氨基酸序列进行聚类分析,结果显示LmAbd-2与沙漠蝗SgAbd-2聚为一支,具有较近的亲缘关系(图2-B)。
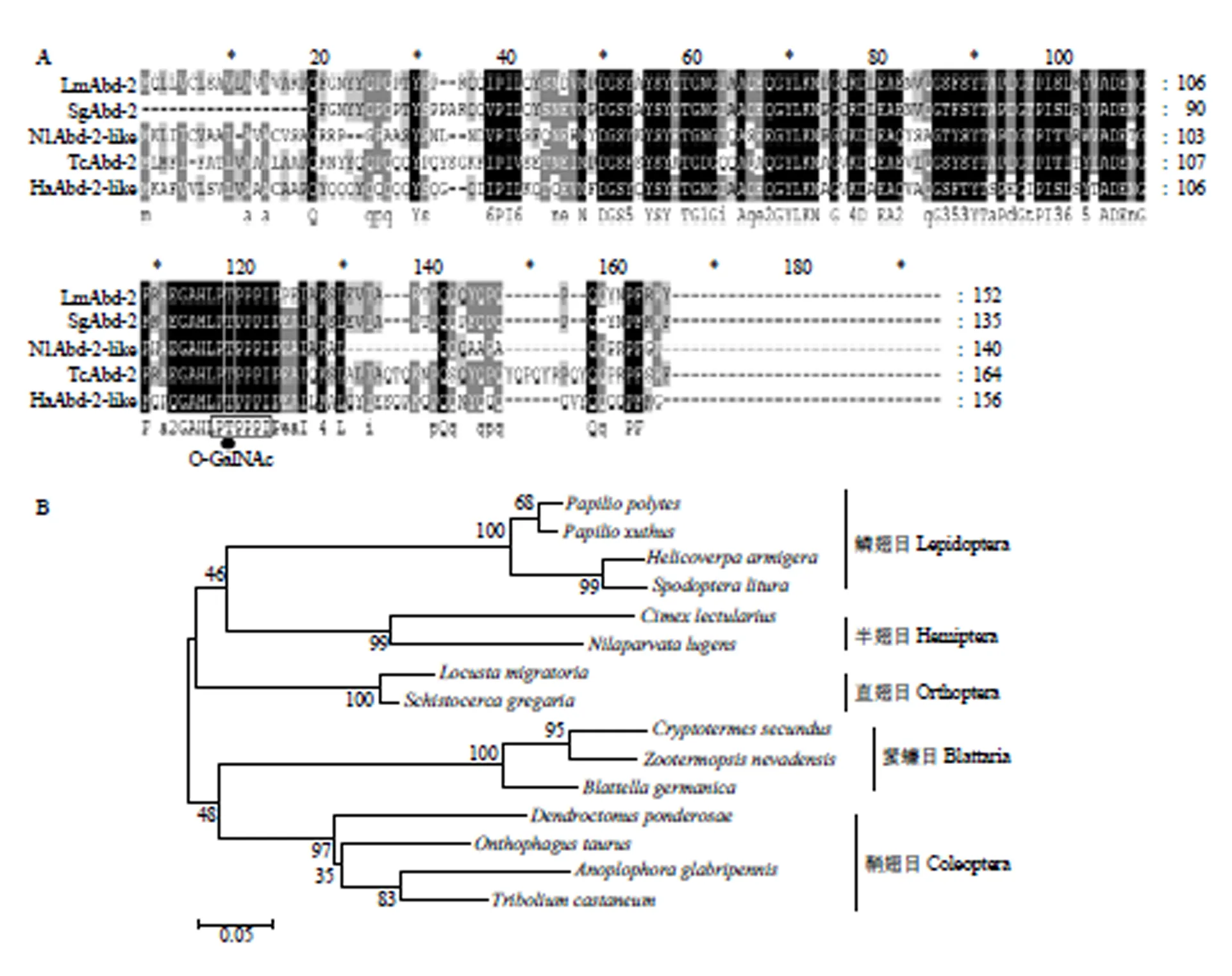
A:LmAbd-2与其他昆虫表皮蛋白Abd-2多序列比对 Multiple sequence alignments of LmAbd-2 and Abd-2 from different insect species;B:LmAbd-2与其他昆虫Abd-2的系统聚类分析 the phylogenetic analysis of LmAbd-2 and Abd-2 from different insect species
2.3 LmAbd-2组织特异性和发育时期表达
RT-qPCR分析结果显示在体壁中高表达,在其他组织中低表达或不表达(图3-A)。发育时期结果显示,在4龄(N4D1—N4D2)、5龄(N5D1—N5D3)和成虫早期(AD1—AD3)呈周期性高表达,其他时期表达较低(图3-B),这种表达模式与内表皮形成时间一致,推测其可能参与飞蝗内表皮结构的形成。
2.4 LmAbd-2的生物学功能
为了探讨在飞蝗表皮形成中的作用,体外合成dsRNA并向4龄第3天飞蝗若虫注射10 μg的ds和ds,正常饲养至蜕皮。结果发现处理组和对照组均能正常蜕皮至5龄且无肉眼可见表型(被显著沉默)。向该5龄期若虫再次分别注射10 μg的ds和ds,结果显示再次注射ds的若虫和对照组仍均能正常蜕皮,尽管在蜕至成虫72 h后表达量显著降低(处理组中表达量显著降低了97.8%)(图4-A),但未出现肉眼可见的异常表型。H&E染色结果显示,与对照组相比,注射ds的处理组表皮层结构变薄(减少约40%)(图4-B);透射电镜结果显示,与对照组相比,处理组内表皮片层数减少(约减少6层),使得整个表皮层结构变薄(图4-C),表明参与飞蝗内表皮结构的形成。
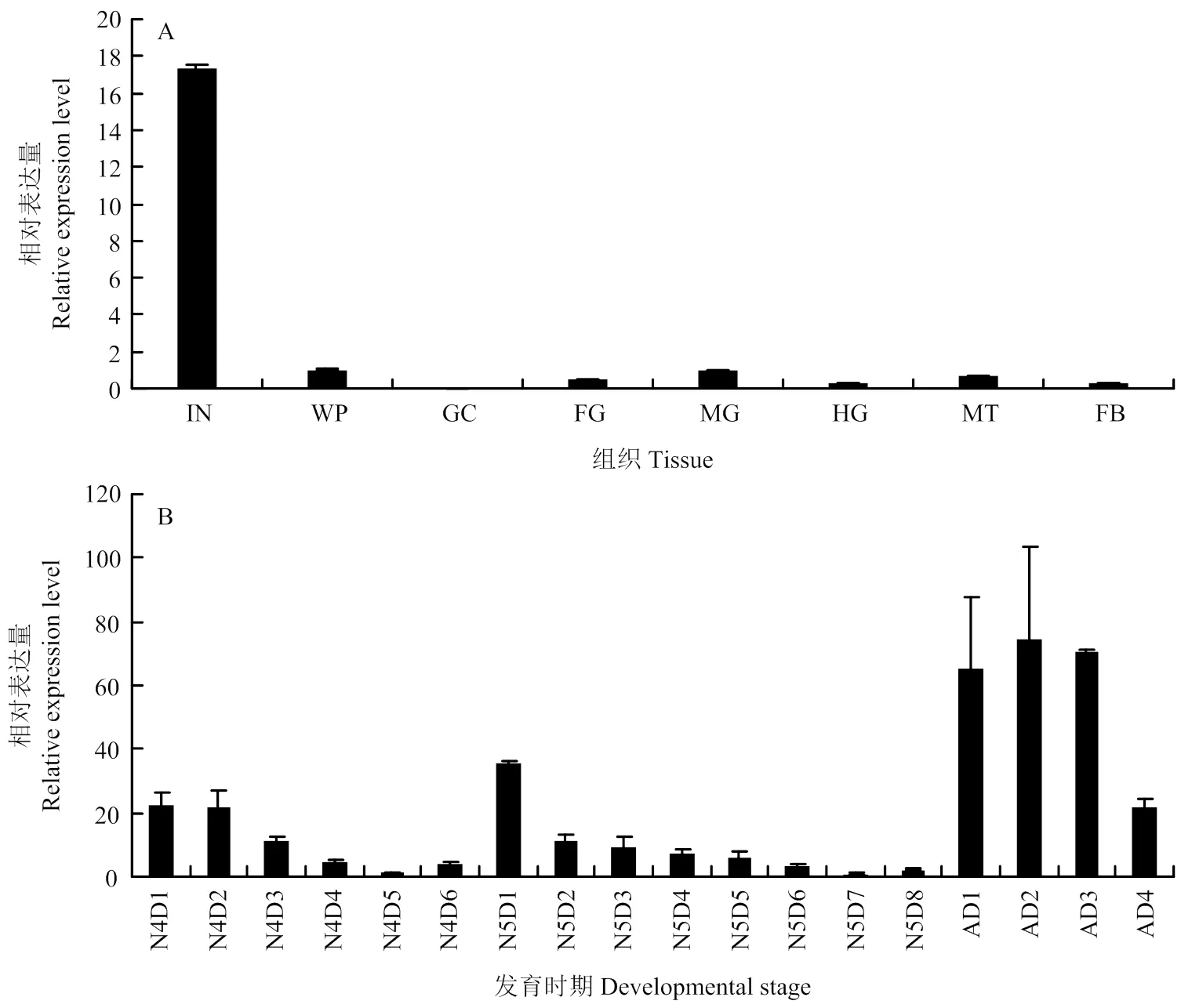
A:LmAbd-2在5龄飞蝗不同组织中的表达 The relative expression of LmAbd-2 in different tissues of 5th instar nymphs。IN:体壁 Integument;WP:翅芽 Wing pads;GC:胃盲囊Gastric caeca;FG:前肠 Foregut;MG:中肠 Midgut;HG:后肠 Hindgut;MT:马氏管Malpighian tubules;FB:脂肪体 Fat body;B:LmAbd-2在飞蝗4龄(第1—6天)、5龄(第1—8天)和成虫早期(第1—4天)不同发育天数的表达The relative expression of LmAbd-2 in different days of 4th instar nymphs (Day 1 to day 6), 5th instar nymphs (Day 1 to day 8), and early stages of adult (Day 1 to day 4)
3 讨论
体壁是覆盖在昆虫身体表面的保护性和支撑性的结构,其主要成分是表皮蛋白和几丁质,其成分的缺失或含量变化都可能影响体壁结构的形成,进而影响昆虫的正常生长发育。本文根据飞蝗转录组数据库获得一个蜕皮后表皮蛋白基因,其编码蛋白序列特征分析发现含有一个RR-1基序(GxFxYxxPDGxxxxVxYxADENGYQPxGAHLP),与沙漠蝗SgAbd-2的氨基酸序列具有92.5%的相似度(信号肽除外),均属于CPR家族RR-1亚类[4]。随着基因组学和转录组学的发展,除直翅目飞蝗和沙漠蝗外,有很多昆虫物种的Abd-2或Abd-2 like被鉴定,如鳞翅目棉铃虫、半翅目褐飞虱、蜚蠊目德国小蠊、以及鞘翅目赤拟谷盗等。然而,这些基因仅仅是序列在NCBI中报道,尚未有功能研究报道。
有研究报道,RR-1亚类的表皮蛋白主要存在于柔软表皮中,如双翅目和鳞翅目幼虫表皮、蝗虫节间膜和内表皮中[6,22]。昆虫在蜕皮过程中表皮依次形成片层结构,其中蜕皮前主要形成上表皮和外表皮,而内表皮主要在蜕皮后开始形成和沉积,蜕皮后约72 h完全形成[29]。组织和时期表达结果显示主要在飞蝗体壁中高表达,而且发现其在所试飞蝗各个龄期内表皮形成时期呈周期性高表达(图3),暗示其可能参与飞蝗内表皮的合成和沉积。利用飞蝗对RNAi的敏感性,本文首先在4龄若虫期沉默的表达,随后在5龄若虫期连续干扰的表达,虽然均未出现肉眼可见的异常表型,但从显微和超微结构分析发现内表皮片层结构减少导致表皮层结构变薄(图4),表明参与飞蝗内表皮结构的形成。该基因与家蚕假设表皮蛋白基因、褐飞虱未分组表皮蛋白基因以及飞蝗其他家族表皮蛋白基因功能类似[16-17,27],均在昆虫内表皮结构形成具有重要作用。
根据表皮片层结构形成时期的不同,Andersen利用MALDI-MS分析方法从沙漠蝗蜕皮后表皮组织中鉴定出8种RR-1亚类蜕皮后表皮蛋白[22],课题组前期从飞蝗转录组数据库中鉴定到与这8种沙漠蝗蜕皮后表皮蛋白同源序列[23],其中研究发现LmAbd-5参与飞蝗内表皮的形成[24]。然而,沉默的表达后,结果与LmAbd-2类似均未引起飞蝗肉眼可见的异常表型。Pan等[10]利用RNAi方法对褐飞虱表皮蛋白基因功能研究发现仅部分基因产生致死表型,推测表皮蛋白可能与其他表皮蛋白相互作用而存在互补效应。在飞蝗中,这8种蜕皮后表皮蛋白在蜕皮过程中可能存在协同或互补作用,但尚有待进一步的研究。

A:若虫期连续注射dsLmAbd-2蜕皮至成虫72 h后LmAbd-2的相对表达量 The relative expression of LmAbd-2 determined by RT-qPCR after injected with dsLmAbd-2 72 h。B:注射dsRNA后,蜕皮至成虫72 h的表皮显微结构 After injection of dsRNA, microstructure and magnification of cuticle by H&E staining;红色为细胞质及细胞外基质,蓝色为细胞核cytoplasm and extracellular matrix are stained red with eosin, nuclei are stained dark blue with hematoxylin (Scale bars, 20 μm),Cut:表皮Cuticle;Ec:表皮细胞Epidermal cell。C:注射dsRNA后,蜕皮至成虫72 h的表皮超微结构观察Ultrastructure observation of adult cuticle at 72 h after injected with dsRNA。Epi:上表皮Epicuticle;Exo:外表皮Exocuticle;Endo:内表皮Endocuticle
4 结论
飞蝗是一个内表皮结构糖蛋白基因,其编码的蛋白含有一个信号肽和一个几丁质结合域ChtBD4,属于CPR家族RR-1亚类;主要在飞蝗体壁组织中高表达,而且在所试飞蝗各龄期内表皮形成时期呈周期性高表达;连续在若虫期干扰后未出现肉眼可见的表型,但内表皮片层结构减少,表明其参与飞蝗内表皮结构的形成。
[1] Moussian B, Seifarth C, Müller U, Berger J, SchwarzH. Cuticle differentiation duringembryogenesis., 2006, 35(3): 137-152.
[2] Vincent J F, Wegst U G. Design and mechanical properties of insect cuticle.,2004, 33(3): 187-199.
[3] Noh M Y, Muthukrishnan S, Kramer K J, Arakane Y. Development and ultrastructure of the rigid dorsal and flexible ventral cuticles of the elytron of the red flour beetle,.,2017, 91: 21-33.
[4] Willis J H. Structural cuticular proteins from arthropods: annotation, nomenclature, and sequence characteristics in the genomics era.,2010, 40(3): 189-204.
[5] Karouzou M V, Spyropoulos Y, Iconomidou V A, Cornman R S, Hamodrakas S J, Willis J H.cuticular proteins with the R&R Consensus: annotation and classification with a new tool for discriminating RR-1 and RR-2 sequences.,2007, 37(8): 754-760.
[6] Cornman R S, Togawa T, Dunn W A, He N, Emmons A C, Willis J H. Annotation and analysis of a large cuticular protein family with the R&R Consensus in.,2008, 9: 22.
[7] Futahashi R, Okamoto S, Kawasaki H, Zhong Y S, Iwanaga M, Mita K, Fujiwara H. Genome-wide identification of cuticular protein genes in the silkworm,.,2008, 38(12): 1138-1146.
[8] Ioannidou Z S, Theodoropoulou M C, Papandreou N C, Willis J H, Hamodrakas S J. CutProtFam-Pred: detection and classification of putative structural cuticular proteins from sequence alone, based on profile hidden Markov models.,2014, 52: 51-59.
[9] Dittmer N T, Tetreau G, Cao X, Jiang H, Wang P, Kanost M R. Annotation and expression analysis of cuticular proteins from the tobacco hornworm,.,2015, 62: 100-113.
[10] Pan P L, Ye Y X, Lou Y H, Lu J B, Cheng C, Shen Y, Moussian B, Zhang C X. A comprehensive omics analysis and functional survey of cuticular proteins in the brown planthopper.,2018, 115(20): 5175-5180.
[11] Guan X, Middlebrooks B W, Alexander S, WassermanS A. Mutation of TweedleD, a member of an unconventional cuticle protein family, alters body shape in.,2006, 103(45): 16794-16799.
[12] Arakane Y, Lomakin J, Gehrke S H, Hiromasa Y, Tomich J M, Muthukrishnan S, Beeman R W, Kramer K J, Kanost M R. Formation of rigid, non-flight forewings (Elytra) of a beetle requires two major cuticular proteins.,2012, 8(4): e1002682.
[13] Qiao L, Xiong G, Wang R X, He S Z, Chen J, Tong X L, Hu H, Li C L, Gai T T, Xin Y Q, LIU X F, CHEN B, XIANG Z H, LU C, DAI F YMutation of a cuticular protein,, alters larval body shape and adaptability in silkworm,.,2014, 196(4): 1103-1115.
[14] Mun S, Young Noh M, Dittmer N T, Muthukrishnan S, Kramer K J, Kanost M R, Arakane Y. Cuticular protein with a low complexity sequence becomes cross-linked during insect cuticle sclerotization and is required for the adult molt.,2015, 5: 10484.
[15] Noh M Y, Muthukrishnan S, Kramer K J, Arakane Y.RR-1 cuticular protein TcCPR4 is required for formation of pore canals in rigid cuticle.,2015, 11(2): e1004963.
[16] Xiong G, Tong X L, Gai T T, Li C, Qiao L, Monteiro A, Hu H, Han M J, Ding X, Wu S Y, XIANG Z H, LU C, DAI F YBody shape and coloration of silkworm larvae are influenced by a novel cuticular protein.,2017, 207(3): 1053-1066.
[17] Lu J B, Luo X M, Zhang X Y, Pan P L, Zhang C X. An ungrouped cuticular protein is essential for normal endocuticle formation in the brown planthopper.,2018, 100: 1-9.
[18] Petkau G, Wingen C, Jussen L C, Radtke T, Behr M. Obstructor-A is required for epithelial extracellular matrix dynamics, exoskeleton function, and tubulogenesis.,2012, 287(25): 21396-21405.
[19] 王燕, 李大琪, 刘晓健, 李涛, 马恩波, 范仁俊, 张建珍. 飞蝗表皮蛋白家族基因的分子特性及基于RNAi的功能分析. 中国农业科学,2015, 48(1): 73-82.
WANG Y, LI D Q, LIU X J, LI T, MA E B, FAN R J, ZHANG J Z. Molecular characterization and RNAi-based functional analysis offamily genes in., 2015, 48(1): 73-82. (in Chinese)
[20] Tajiri R, Ogawa N, Fujiwara H, Kojima T. Mechanical control of whole body shape by a single cuticular protein obstructor-E in.,2017, 13(1): e1006548.
[21] Jan S, Liu S, Hafeez M, Zhang X, Dawar F U, Guo J, Gao C, Wang M. Isolation and functional identification of three cuticle protein genes during metamorphosis of the beet armyworm,.,2017, 7: 16061.
[22] Andersen S O. Amino acid sequence studies on endocuticular proteins from the desert locust,.,1998, 28(5/6): 421-434.
[23] Zhao X M, Gou X, Qin Z Y, Li D Q, Wang Y, Ma E B, Li S, Zhang J Z. Identification and expression of cuticular protein genes based ontranscriptome.,2017, 7: 45462.
[24] 赵小明, 贾盼, 勾昕, 刘卫敏, 马恩波, 张建珍. 飞蝗内表皮蛋白基因的表达与功能分析. 中国农业科学,2017, 50(10): 1817-1826.
ZHAO X M, JIA P, GOU X, LIU W M, MA E B, ZHANG J Z. Expression and functional analysis of endocuticle structural glycoprotein genein., 2017, 50(10): 1817-1826. (in Chinese)
[25] Zhao X M, Qin Z Y, Liu W M, Liu X J, Moussian B, Ma E B, Li S, Zhang J Z. Nuclear receptor HR3 controls locust molt by regulating chitin synthesis and degradation genes of.,2018, 92: 1-11.
[26] Song T Q, Yang M L, Wang Y L, Liu Q, Wang H M, Zhang J, Li T. Cuticular protein LmTwdl1 is involved in molt development of the migratory locust.,2016, 23(4): 520-530.
[27] 杨亚亭, 赵小明, 秦忠玉, 刘卫敏, 马恩波, 张建珍. 飞蝗表皮蛋白基因的分子特性及功能分析. 中国农业科学,2018, 51(7): 1303-1314.
YANG Y T, ZHAO X M, QIN Z Y, LIU W M, MA E B, ZHANG J Z. Molecular characteristics and function analysis of cuticle protein gene., 2018, 51(7): 1303-1314. (in Chinese)
[28] Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2ΔΔCtmethod.,2001, 25(4): 402-408.
[29] Charles J P. The regulation of expression of insect cuticle protein genes.,2010, 40(3): 205-213.
(责任编辑 岳梅)
Expression and Function Analysis of Endocuticle structural Glycoprotein Gene
Jia Pan1,2, Zhang Jing1,2, Yang Yang1,2, Liu WeiMin1, Zhang JianZhen1, Zhao XiaoMing1
(1Research Institute of Applied Biology, Shanxi University, Taiyuan 030006;2College of Life Science, Shanxi University, Taiyuan 030006)
【Objective】The objective of this study is to analyze the sequence and expression characteristics of endocuticle structural glycoprotein genethat identified fromtranscriptome, explore its function by Hematoxylin and eosin (H&E) staining and transmission electron microscopy (TEM) based on RNAi with ds, and to discuss the effect on the formation of endocuticle duringmolting.【Method】The cDNA sequence ofwas obtained according to the transcriptome database of, and was cloned by reverse-transcription PCR (RT-PCR) and sequenced. The amino acid sequence of LmAbd-2 was translated by ExPASy tool, and its signal peptide and structural domain were analyzed by using SignaIP4.1Server and SMART software, respectively. Meanwhile, the O-linked glycosylation sites were predicted by using NetOGlyc4.0 Server. The homologous sequences of Abd-2 from locust and other insect species were aligned by GENEDOC software and evolutionary tree was constructed by using the MEGA 6.0 software with the neighbor-joining (NJ) method. Expression profiles ofdifferent tissues at day 2 of 5th instar nymphs and developmental days of 4th instar nymphs (N4D1-N4D6), 5th instar nymphs (N5D1-N5D8), and the early stage of adults (AD1-AD4) were assayed by using reverse-transcription quantitative PCR (RT-qPCR). The biological function ofwas analyzed by combination of RNAi and H&E staining and TEM methods. 【Result】The results ofsequence analysisshowed that the length of thecDNA is 683 bp and full length of its ORF is 459 bp, encoding 152 amino acids. The functional domainanalysis showed that the protein encoded bycontains one signal peptide and one chitin binding domain (ChtBD4), which belongs to RR-1 subclass of the CPR family. Sequence alignment analysis showed that it is highly conserved among species, and the similarity with SgAbd-2 is 92.5%, and it contains conserved motif (PTPPPIP) in N-terminal and has a potential O-linked glycosylation site (T116) at which glycosylation modification may occur. Phylogenetic tree analysis showed that LmAbd-2 has a close genetic relationship with SgAbd-2 of. The results of RT-qPCR revealed thatwas highly expressed in the integument, but lower or no expression in other tested tissues, and showed periodic expression during molting. After injection of dson day 3 of 4th instar nymphs and day 4 of 5th instar nymphs, the insects could normally molt to adults and showed no macroscopic phenotype; however, the cuticle of the adults was thinner, and there were significantly fewer endocuticular lamellae than those in the control.【Conclusion】An endocuticle structural glycoprotein genewas obtained according to thetranscriptome database. The protein encoded bybelongs to RR-1 subclass of CPR family.was mainly expressed in integument among all the tested tissues and showed periodic expression at the early stage of 4th instar nymphs, 5th instar nymphs and adults. The results of RNAi suggested thatwas involved in the formation of endocuticle duringmolting.
; endocuticle; glycoprotein;; RNAi
10.3864/j.issn.0578-1752.2019.04.007
2018-09-29;
2018-11-15
国家自然科学基金(31702067,31640075)、山西省青年科学基金(201601D021102)
贾盼,Tel:0351-7018871;E-mail:120768708@qq.com。 通信作者赵小明,Tel:0351-7018871;E-mail:zxming@sxu.edu.cn
猜你喜欢
玩具世界(2022年3期)2022-09-20
农业工程学报(2022年10期)2022-08-22
今日农业(2022年3期)2022-06-05
天津农业科学(2022年5期)2022-05-31
建材发展导向(2021年14期)2021-08-23
饲料博览(2020年11期)2020-12-30
甘肃教育(2020年2期)2020-09-11
祝您健康·文摘版(2019年4期)2019-06-11
小学生必读(低年级版)(2018年11期)2018-03-13
江苏农业科学(2017年12期)2017-09-16
