
复合微生物菌剂与氨基酸水溶肥组合施用对香蕉土壤理化性质及微生物群落的影响
2019-03-12 01:50柳晓磊陈剑山张曼丽何文娅
中国土壤与肥料 2019年1期
柳晓磊,齐 钊,闫 臻,徐 敏,陈剑山,张曼丽,骆 娟,熊 睿,程 玉,李 阳,何文娅,李 鹏,汤 华*
(1.海南大学热带农林学院,热带生物资源可持续利用重点实验室,海南 海口 570228;2.海南省植物保护总站,海南 海口 571100)
良好的土壤环境是作物生长的重要前提。土壤微生物作为土壤生态系统的重要组成部分,不仅是生态系统的分解者,也是物质循环与能量流动的承担者[1]。它们参与土壤有机质的分解,腐殖质的合成,土壤养分的转化与循环而促使土壤肥力增加[2]。土壤微生物多样性代表着微生物群落的稳定性,也可反映土壤的生态机制和土壤胁迫对微生物群落的影响[3]。微生物群落多样性与土壤生态系统的结构、功能密切相关,在维持土壤肥力和土壤生态平衡中发挥重要作用[4]。
近20年来,中国农业的肥料投入量过大(主要以复合肥为主),施肥不合理,造成农业生态环境问题及土壤问题日益严重。目前,影响耕地土壤的障碍因素包括有机质含量低、微量元素缺乏、土壤肥力差,供肥能力弱、土壤酸化、土壤生物活性下降、养分利用率低、根结线虫危害及土传病害等[5]。随着生态农业的发展和社会对环境保护的日益重视,探索新型肥料(尤其是生物菌肥)弥补和改善传统肥料不足的研究倍受关注[6]。复合微生物菌剂含有枯草芽孢杆菌、地衣芽孢杆菌等大量的有益活菌物质及多种天然发酵活性物质,能在根部土壤繁殖形成有利于作物生长的微生物优势菌群,调节根部的营养环境,恢复土壤微生态平衡,而且在其生命活动过程中还能产生各类植物生长激素,刺激植物生长。氨基酸水溶肥易于被作物吸收,也能为枯草芽孢杆菌等提供营养物质,有利于土壤微生物菌扩繁[7]。土壤中需要植物所必需的各种元素才能使植物发育良好,氨基酸水溶肥中含有的微量元素Mn、Zn、B、Fe等,均为植物所需的必需微量元素,能够改善土壤营养状况,保证植物对养分的需求,是植物健康生长的又一保障。土壤微生物通过微生物-土壤-植物模式影响植物的生长和发育[8]。复合微生物菌剂与氨基酸水溶肥组合施用在豇豆上,可以改善土壤理化性质,提高土壤肥力,对豇豆的枯萎病及根系线虫都有明显的抑制作用[7,9]。
香蕉枯萎病是典型的土传病害,一旦感染,对香蕉园是毁灭性的。根结线虫是一类在植物上危害极为严重的寄生线虫[10-11],其种类繁多,分布较广,致病性强,已引起世界各国的关注[12-14]。根结线虫在香蕉根部造成大量伤口,为枯萎病菌的入侵提供途径,促进了病害发生。组合施用复合微生物菌剂和氨基酸水溶肥,是否有利于防控香蕉枯萎病及根结线虫,未见相关报道。本研究通过香蕉大田试验,开展混合施用复合微生物菌剂与氨基酸水溶肥的试验,研究土壤理化性质及土壤微生物菌群的变化情况,为香蕉枯萎病的防控提供新的思路。
1 材料与方法
1.1 试验方案及样品采集
试验于2016年在海南省东方市红泉农场一队的香蕉种植示范区进行,试验香蕉品种为“南天黄”,第一造蕉种植面积14 hm2,试验地为红土坡地,坡度≤15℃,肥力中等,排灌设施良好,为滴灌。供试复合微生物菌剂的有效成分:枯草芽孢杆菌+地衣芽孢杆菌+解淀粉芽孢杆菌约2亿/mL,氨基酸水溶肥的有效成分:氨基酸=100 g/L,Mn+Zn+B+Fe=30 g/L。
本次大田试验共设2组处理,10 hm2地平均分成3部分并混合施用复合微生物菌剂与氨基酸水溶肥,为处理组(BE),处理组在香蕉的苗期、中期、孕蕾期3个阶段分别灌溉一次复合微生物菌剂(15 kg/hm2)和氨基酸水溶肥(15 kg/hm2);另外4 hm2为对照组(BC),同期处理时滴灌清水;其它的水肥管理和病虫害防控相同。在第3次处理结束后30 d,对两组处理分别采用5点取样法取样,选取5~15 cm土样混合,每个处理设置3个重复。
1.2 土壤理化性质的测定
土壤碱解氮的测定采用碱解蒸馏法,有效磷的测定参照NY/T 1121.7-2014,速效钾的测定参照NY/T 889-2004,有机质的测定参照 NY/T 1121.6-2006,pH值的测定参照NY/T 1121.6-2016。线虫的检测采用改良贝曼漏斗法分离24 h,用体视显微镜和生物显微镜对分离出的根结线虫进行数量检测,每个样品均测定3次。
1.3 土壤DNA提取与土壤微生物高通量测序
土壤微生物的DNA提取采用Genebank土壤微生物DNA提取试剂盒,取质量合格的DNA样品,采用Illumina Miseq 测序平台进行高通量测序。
使用 FLASH v1.2.7软件进行序列的拼接,然后使用 Trimmomatic v0.33 软件,对拼接得到的 Raw Tags进行过滤,最后使用 UCHIME v4.2软件,鉴定并去除嵌合体序列,得到有效数据。采用Excel 2016和 SPSS 21 进行数据分析和处理。
2 结果与分析
2.1 不同处理香蕉土壤理化性质的变化及分析
混合施用复合微生物菌剂与氨基酸水溶肥的为处理组(BE);滴灌清水的为对照组(BC)。对试验地土壤的碱解氮(Alk-hydr. N)、有效磷(AP,available phosphorus)、速效钾(AK,available potassium)、有机质(OM,organic matter)、pH 值以及根结线虫(RN,root-knot nematode)的检测表明(表1):相比于对照组,BE组土壤pH值没有明显变化;BE组有机质含量比BC组增加了80.34%;BE组土壤中碱解氮的含量显著上升(P<0.05),有效磷和速效钾的含量极显著上升(P<0.01)。根结线虫的数量显著下降(P<0.05)。可见,微生物菌剂和氨基酸水溶肥的联合使用对提高土壤中氮磷钾的含量影响显著,从而改善了土壤的营养情况。植物在微量元素充足的情况下,生理机能就会十分旺盛,因此氨基酸水溶肥中含有的微量元素,有利于香蕉对大量元素的吸收和利用。复合微生物菌剂与氨基酸水溶肥组合施用对土壤中根结线虫的繁殖具有抑制作用,根结线虫数量的减少对抑制香蕉枯萎病的发生具有积极意义。
2.2 香蕉土壤微生物的多样性及序列统计
在土壤微生物的高通量测序中,真菌和细菌的覆盖率可以反映土壤微生物的状况和测序数据可靠性。稀释曲线可以作为对各样本测序量是否充分的判断,曲线急剧上升表明测序量不足,需要增加序列条数;反之,则表明样品序列充分,可以进行数据分析。本研究的细菌稀释曲线和真菌稀释曲线都基本趋于平缓,说明测序量充足,覆盖率高。

表1 不同处理下土壤的理化性质及线虫数量变化
香蕉土壤微生物测序结果表明,细菌和真菌分别获得有效序列573 433和488 444条。细菌获得的分类单元OTU数是1 375个,真菌获得OTU数是668个(表2)。真菌和细菌的覆盖率都达到了99%以上,说明土壤中微生物的菌群结构和种类很稳定。从丰富度指数和香农指数可以看出,香蕉土壤中细菌的种类和数量远远高于真菌,进一步证明细菌是土壤微生物的主体部分,与其他相关报道结果一致[1,3]。
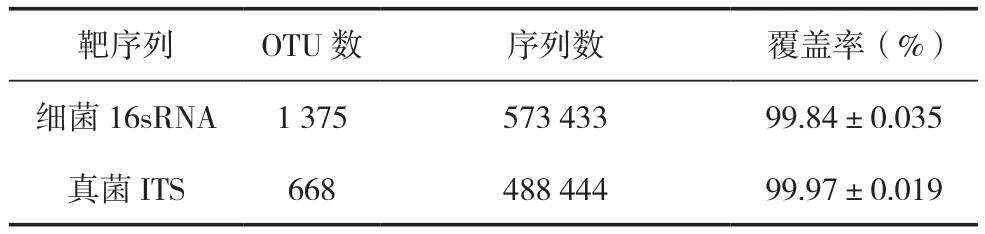
表2 香蕉土壤微生物OTUs多样性分析
2.3 香蕉土壤细菌和真菌的菌群结构分析
土壤细菌测序结果中,每个测序样品所得到的序列数约为79 916~ 80 223条,OTU数目在1 083~1 275条之间。如图1(左)所示,处理组BE和对照组BC之间在OTU的数量和种类上都存在差异。两组共有的OTU数为1 353个,BE特有的OTU数为18个,而BC特有的OTU数仅4个。
土壤真菌测序结果中,每个测序样品所得到的序列数约为79 881~ 80 152条,OTU数目在299~437条之间。如图1(右)所示,处理组BE和对照组BC之间在OTU的数量和种类上都存在差异。两组共有的OTU数为342个,BE特有的OTU数为51个,而BC特有的OTU数为42个。
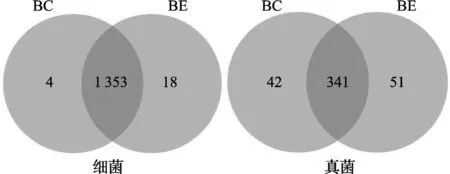
图1 香蕉土壤细菌和真菌的OTU-Venn图
2.4 香蕉土壤细菌和真菌的多样性分析
Ace和Chao1指数都是衡量物种丰度的指标,其值越高,物种的丰富度越高。如表3所示,香蕉地土壤细菌测序结果中,BC组的Ace和Chao1均高于BE组,说明BC组细菌菌群的物种丰富度高于BE组;真菌测序结果中,BC组的Ace和Chao1均低于BE组,说明了BE组真菌菌群的物种丰富度高于BC组。综上所述,不同处理下的香蕉地土壤细菌和真菌的物种丰富度均发生了变化,施肥后土壤细菌菌群丰富度降低的同时真菌菌群丰富度增加。
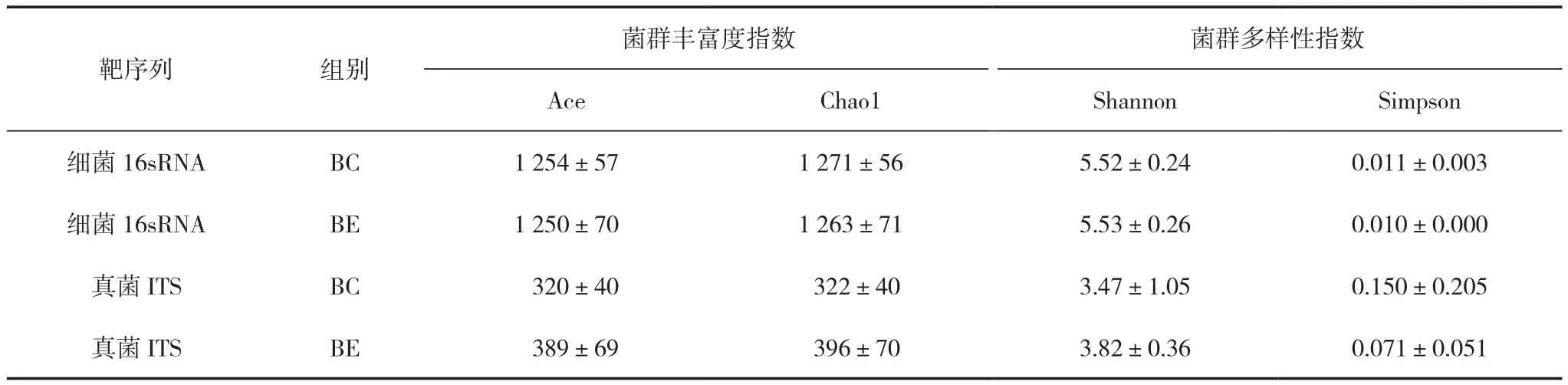
表3 香蕉土壤微生物的多样性分析
相同物种丰度的情况下,群落中各物种具有越大的均匀度,则认为群落具有越大的多样性,即香农指数(Shannon)值越大,辛普森指数(Simpson)值越小,说明样品的物种多样性越高。如表3所示,细菌测序结果中,BE组的Shannon高于BC组,Simpson低于BC组,说明了BE组的物种多样性高于BC组,这与真菌测序结果中Shannon与Simpson的指数对比情况相同。综上所述,不同处理下的香蕉地土壤细菌和真菌的物种多样性的变化规律保持一致,相较于BC组,BE组的物种多样性增加。
2.5 香蕉土壤细菌菌群群落分析
土壤的细菌测序,获得573 433条序列,隶属于23门59纲79目130科148属。在门水平上(图2),对照组BC中,相对丰度≥1%的菌门包括Proteobacteria(变形菌门)41.62%和Acidobacteria(酸杆菌门)25.85%,两者占比最高,是土壤中的优势菌种;其他分别为Actinobacteria(放线菌门)10.88%、Chloroflexi(绿弯菌门)8.12%、Gemmatimonadetes(芽单胞菌门)3.51%、Firmicutes(厚壁菌门)2.99%、Verrucomicrobia(疣微菌门)2.58%。而在处理组BE中,相对丰度≥1%的优势菌门仍然是Proteobacteria(变形菌门)42.65%和Acidobacteria(酸杆菌门)23.86%,其他菌门与对照组BC大致相同。
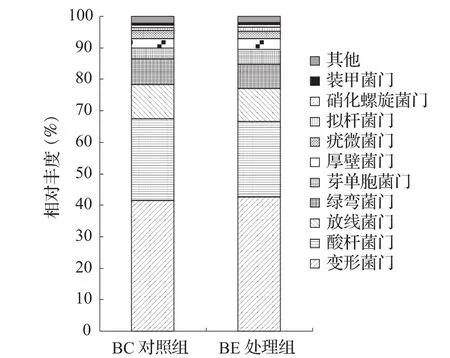
图2 香蕉土壤细菌门水平物种分布柱状图
在纲水平上(表4),BC组相对丰度≥2%的有Alphaproteobacteria(α-变形杆菌纲)25.63%、Acidobacteria(酸杆菌纲)17.36%、Betaproteobacteria(β-变形菌纲)8.91%、Thermoleophilia(嗜热油菌纲)6.68%、Gammaproteobacteria(丙型变形菌纲)5.14%、JG37-AG-4(分类未定)4.58%、Gemmatimonadetes(芽单胞菌纲)3.51%、Solibacteres(拟杆菌纲)3.71%、Clostridia(梭状芽胞杆菌纲)2.94%、Subgroup_2(分类未定)2.91%。BE组相对丰度≥2%的菌群种类与BC组没有明显差异,但在相对丰度上则存在较大差异:Acidobacteria(酸杆菌纲)相对丰度下降了13.47%;Betaproteobacteria(β-变形菌纲)下降了19.30%,Gammaproteobacteria(丙型变形菌纲)下降了45.52%;Gemmatimonadetes(芽单胞菌纲)增长了33.05%;Clostridia(梭状芽胞杆菌纲)则增长了11.90%。其中,BC和BE组中Acidobacteria、Gemmatimonadetes、Clostridia的相对丰度存在显著性差异。
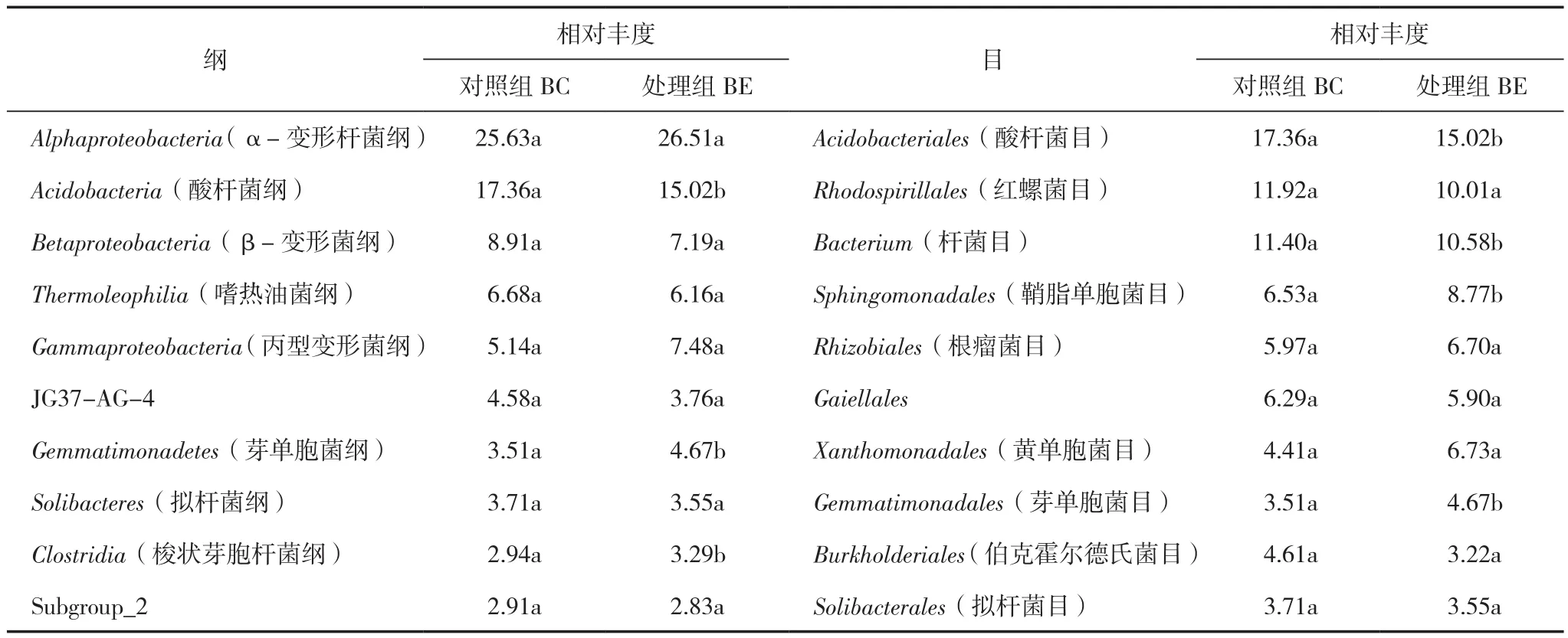
表4 纲和目水平细菌菌群组成及其相对丰度 (%)
在目水平上(表4),BC组相对丰度≥2%的前10位中,Acidobacteriales(酸杆菌目)17.36%、Rhodospirillales(红螺菌目)11.92%、Bacterium(杆菌目)11.4%是优势菌目;BE组相对丰度≥2%的前10位菌群与BC组相比,Sphingomonadales(鞘脂单胞菌目)相对丰度增长了34.3%;Rhizobiales(根瘤菌目)增长了12.23%;Xanthomonadales(黄单胞菌目)增长了52.61%;Gemmatimonadales(芽单胞菌目)增长了33.05%;Acidobacteriales(酸杆菌目)、Rhodospirillales(红螺菌目)、Burkholderiales(伯克霍尔德氏菌目)则分别下降了13.48%、16.02%和30.15%。其中BC组与BE组中的Acidobacteriales、Bacterium、Sphingomonadales、Gemmatimonadales的相对丰度呈显著性差异。
2.6 香蕉土壤真菌菌群结构分析
BC组真菌菌群隶属11门20纲44目66科106属。在门水平上(图3),对照组BC的土壤真菌主要有Ascomycota(子囊菌门)33.11%、Basidiomycota(担子菌门)6.64%、Rotifera 0.99%,还有丰度极少的 Mortierellomycota 0.15%、Glomeromycota(球囊菌门)0.19%、Mucoromycota(毛霉亚门)0.09%、Cercozoa(虫门)0.03%、Neocallimastigomycota(新丽鞭毛菌门)0.03%、Kickxellomycota(梳霉亚门)0.01%、57.04%的未知真菌和1.71%未能分类真菌。处理组BE的土壤真菌群落结构在门水平上出现了明显的变化,主要包括Ascomycota(子囊菌门)占38.51%,明显上升,Basidiomycota(担子菌门)占2.19%,明显下降。
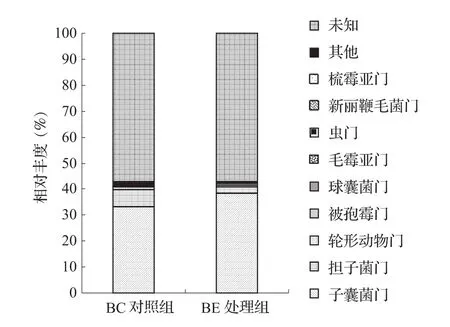
图3 香蕉土壤真菌门水平物种分布柱状图
在纲水平上(表5),BC组占优势的是子囊菌门的Sordariomycetes(子囊菌纲)23.76%和Eurotiomycetes(散囊菌纲)4.21%,以及担子菌门的Agaricomycetes(伞菌纲)5.80%。相较于BC组,BE组占优势的是子囊菌门的Sordariomycetes(子囊菌纲),物种丰度增加了15.32%,Dothideomycetes(座囊菌纲)升高了58.20%,而担子菌门的Agaricomycetes(伞菌纲)物种丰度降低了75%。但BC组和BE组的优势菌纲丰度并没有显著差异。
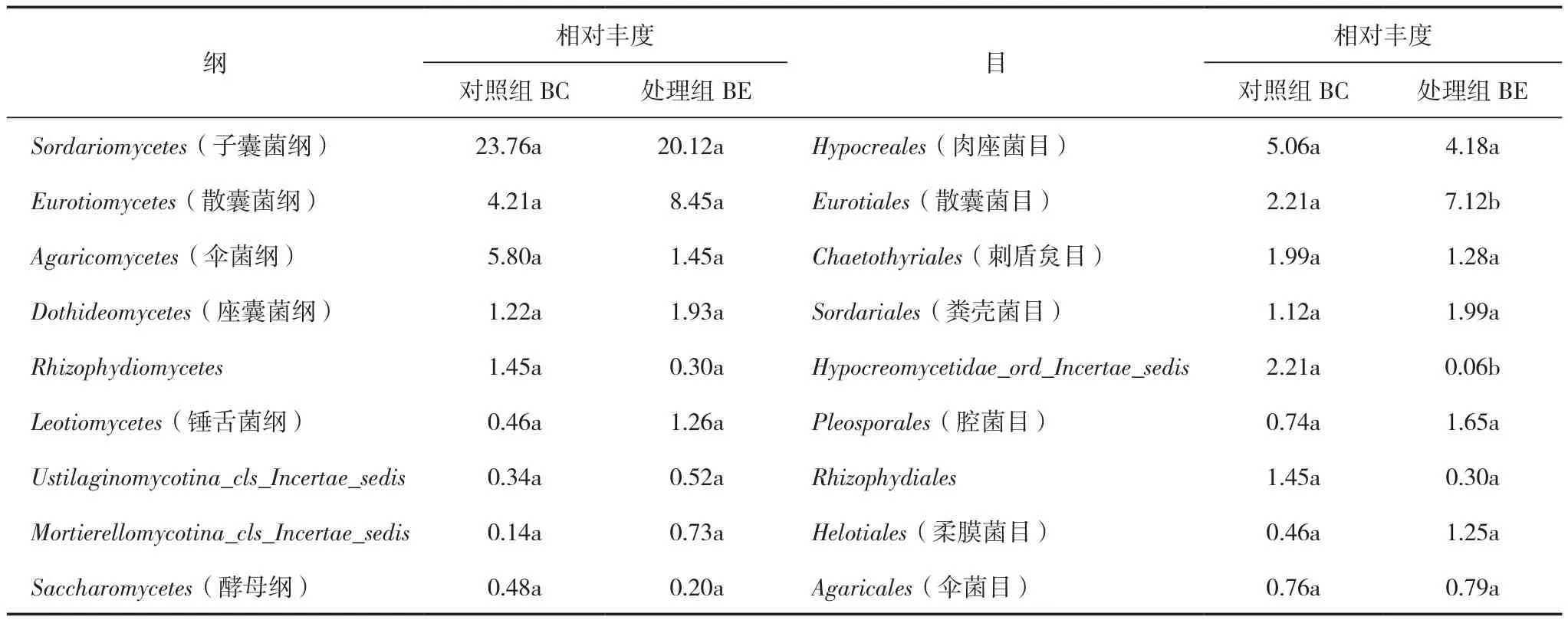
表5 香蕉土壤纲和目水平真菌菌群组成及其相对丰度 (%)
在目水平上(表5),BC组丰富度≥1%的前5菌目有子囊菌门的Eurotiales(散囊菌目)2.21%和Chaetothyriales(刺盾炱目)1.99%、Hypocreales(肉座菌目)5.06%、Sordariales(粪壳菌目)1.12%、Hypocreomycetidae_ord_Incertae_sedis 2.11%。BC 组中85.18%都是目前尚不知晓的属,物种丰度≥1%的菌群不多,主要包括Boothiomyces 1.45%、Colletotrichum(炭疽菌属)1.93%、Penicillium(青霉菌属)1.13%。BE组相对丰度≥1%前5的优势菌目有Eurotiales(散囊菌目)7.12%、Hypocreales(肉座菌目)4.18%、Chaetothyriales(刺盾炱目)1.28%、Sordariales(粪壳菌目)1.99%、Pleosporales(腔菌目)1.65%。其中BC组和BE组中Eurotiales、Hypocreomycetidae_ord_Incertae_sedis的相对丰度存在显著性差异。
香蕉枯萎病的病原菌为古巴尖镰孢菌4号小种[15](Fuaarium oxysporum f. sp. cubense Race 4),本次检验中,对照组和处理组土壤中并没有检测到含有此类病原菌,土壤状况良好,有利于香蕉的生长。测序结果分类显示,83.22%都是目前尚不知晓的属,其中Penicillium(青霉菌属)为5.66%,相较于BC组上升了400.88%,呈显著性差异;Colletotrichum(炭疽菌属)为0.03%,相较于BC组下降了98.45%,呈显著性差异。
2.7 不同处理下香蕉土壤细菌和真菌菌群结构与土壤理化性质的相关性分析
土壤环境因子5个,分别为AN(碱解氮)、AP(有效磷)、AK(速效钾)、OM(有机质)和RN(根结线虫)。RDA相关性分析发现,处理组与对照组样品分别分布在坐标图的两侧,微生物群落结构存在明显的差异,AK、AN、AP与处理组呈正相关,而RN则与对照组呈正相关。图4左为细菌群落-土壤环境因子,其轴1特征值为64.15%,轴2特征值为17.14%,基本覆盖了土壤的环境信息。其中,AP对细菌群落结构影响最大,Rhodanobacter与AP呈正相关,Bradyrhizobium、Variibacter、Acidibacter、Burkholderia-Paraburkholderia 与AP呈负相关。AK和AN对细菌群落结构的影响较大;OM和RN对细菌群落结构也具有一定影响。Sphingomonas对芳香化合物具有生物降解作用,具有高的代谢能力,存在于营养丰富的土壤中,与AK、AN、AP、OM呈正相关。
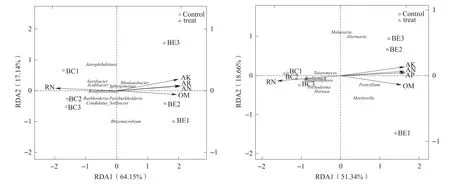
图4 香蕉土壤细菌群落-土壤环境因子(左)与真菌群落-土壤环境因子(右)相关性分析
图4 右是真菌群落-土壤环境因子,其轴1特征值为51.34%,轴2特征值为18.66%,基本上覆盖了大部分的土壤环境信息。其中,AP、AK、AN对真菌群落结构影响最大。Penicillium与三者呈正相关,Talaromyces、Boothiomyces、Colletotrichum与三者呈负相关。OM和RN对真菌群落结构影响较大。Penicillium与OM呈 正 相 关,Talaromyces与OM呈负相关;Talaromyces、Boothiomyces、Colletotrichum与RN呈正相关,Penicillium与RN呈负相关。由相关性可知,RN与AK、AN、AP、OM之间对真菌的影响恰恰相反,说明通过调节真菌菌群结构能够有效降低土壤线虫含量。
氨基酸水溶肥和微生物菌剂的联合使用,有效改善了香蕉土壤的理化性质,降低了根结线虫的数量,香蕉土壤中微生物的菌群结构也发生了变化,通过调节土壤微生物菌群结构来改善土壤性质,减少根结线虫的含量。
3 讨论
3.1 不同处理对香蕉土壤细菌群落结构的影响
本研究表明,香蕉土壤中细菌的种类和丰度远远高于真菌,细菌是土壤微生物组成的主要成员。在两个不同处理间(图2),在门水平上细菌占主要优势的是Proteobacteria、Acidobacteria和Actinobacteria,其中Proteobacteria是土壤中最广泛的优势菌门,该结果和国内外很多土壤多样性的研究结果相一致[16-20]。
也有研究发现Acidobacteria是土壤细菌群落中丰度最高的优势类群[6,21],Acidobacteria多存在于营养贫瘠的土壤环境中[21-22]。根际土壤较非根际土壤 pH 值低(分泌有机酸),有利于酸杆菌某些亚群细菌的生长,但根际的高营养环境又不利于酸杆菌细菌的定殖和繁殖[23]。在本研究中,Acidobacteria是第二大优势菌门,在处理组BE的土壤中,从门、纲、目水平上,Acidobacteria的物种丰度显著性降低,这可能是由于氮、磷、钾等营养环境改善引起的。
Proteobacteria中的Alphaproteobacteria中包括一些有益菌目,比如根瘤菌目,根瘤菌能够促进植物固氮;而β-变形菌纲和丙型变形菌纲中则包括了多数重要的病原菌,尤其是Betaproteobacteria中的伯克霍尔德氏菌目。处理组的土壤中,可能是有害的Betaproteobacteria和Gammaproteobacteria的物种丰度都有明显降低,Alphaproteobacteria的物种丰度则基本不变,但根瘤菌目的物种丰度增长了12.23%,Burkholderiales则下降了13.48%。
3.2 不同处理对香蕉土壤真菌群落结构的影响
在本研究中,香蕉土壤中真菌的优势菌门是Ascomycota和Basidiomycota(图3),其中,Ascomycota的丰度最高,且远高于第二位的Basidiomycota。子囊菌门的真菌多为腐生菌,对降解土壤有机质起着重要作用[24]。在处理组中,子囊菌门的子囊菌纲、座囊菌纲的物种丰度都有较大升高,对提高微生物分解有机质的能力具有促进作用。有研究表明,Colletotrichu中大部分为致病菌,可引起炭疽病[25],Penicillium真菌能产生许多具有抗菌、抗肿瘤和抗病毒功能的活性次生代谢产物,分泌多种高活性的水解酶类,降解多种单环和多环芳香类环境污染物[26]。在处理组中Colletotrichu物种丰度明显下降,而抗菌能力强的Penicillium则显著提高,说明复合微生物菌剂和氨基酸水溶肥的组合施用,提高了有益菌群的丰度,降低了有害菌群丰度。
4 结论
通过组合施用复合微生物菌剂与氨基酸水溶肥,有效地改善了土壤的营养成分,土壤有效磷、速效钾和碱解氮显著增加,土壤根结线虫数量呈显著下降。土壤微生物高通量测序结果表明:土壤菌群结构相对稳定,土壤有害细菌丰度明显下降,有益细菌丰度都明显升高;土壤真菌方面,炭疽菌属含量降低,青霉菌属的丰度有所升高。可见,采用复合微生物菌剂和氨基酸水溶肥组合处理,对香蕉土壤理化性状和土壤微生物群落结构具有明显的改良和改善作用,有利于香蕉枯萎病的防治。
猜你喜欢
科学技术与工程(2022年26期)2022-11-01
当代水产(2022年8期)2022-09-20
云南农业科技(2022年4期)2022-08-02
中国农学通报(2022年14期)2022-06-01
油气田环境保护(2022年2期)2022-05-09
昆明医科大学学报(2022年2期)2022-03-29
中南林业科技大学学报(2021年11期)2021-12-17
中国土壤与肥料(2021年5期)2021-12-02
食品安全导刊(2021年20期)2021-08-30
世界竹藤通讯(2020年5期)2020-11-11
