
抗热应激性能奶牛选育的研究进展
2019-03-08 08:10张华林赵晓铎李倩倩刘光磊
中国奶牛 2019年2期
张华林,赵晓铎,李倩倩,刘光磊
(光明牧业有限公司/上海奶牛育种中心有限公司,上海 200436)
热应激会对奶牛造成严重危害(图1),进而影响牧场经济效益[1]。耐热奶牛品种(品系)培育受到国内外研究学者的广泛关注,研究资料表明,直肠温度是判断奶牛是否处于热应激状态的重要指标,其遗传力在0.25~0.65,在热应激状态下直肠温度较低的奶牛具有更强的抗热应激能力[2]。以日均直肠温度和早上6∶00直肠温度作耐热性标记,根据体温的高低估计耐热系数,系数越高表示耐热能力越强[3]。Turner报道,奶牛直肠温度和生产力存在负相关,即选育直肠温度低的奶牛品种,有利于提高奶牛生产力[4]。West的研究也发现,奶牛耐热性和生产性能的相关性大约为-0.3[5]。近年来分子遗传学作为一种新方法应用于研究奶牛热应激的敏感度和耐受力[6],有一大批抗热应激基因被发掘出来;此外全基因组检测分析(GWAS)也被应用于奶牛抗热应激分子标记的筛选工作[7,8]。本文对热应激给奶牛造成的危害及奶牛抗热应激育种相关研究进展作了简要综述。

图1 奶牛热应激状态下行为和生理指标发生变化的趋势
1 热应激对奶牛的危害
荷斯坦牛的最适泌乳温度为5~20℃(NRC,2001),通常用THI(温湿指数)来衡量奶牛热应激状态,学者根据奶牛生产性能的降低和体温及其他生理状态的改变进行界定[9]:①THI<72时奶牛处于正常状态;②72≤THI<80时奶牛处于中度应激状态;③THI≥80则奶牛处于高度应激状态;④THI超过96则会发生死亡。即THI大于72时奶牛均处于热应激状态[10]。最新研究表明,对于高产奶牛THI阈值应由72降至68[11],随着THI升高,奶牛的产奶量和繁殖受胎率会显著下降[1,12]。对牧场来说,热应激在奶牛生产性能和繁殖性能两个方面的危害表现最为突出(图2)。
1.1 热应激影响奶牛的健康和生产力
有研究发现,THI与泌乳牛干物质采食量和产奶量呈显著负相关[1,13,14],此外还会造成奶牛的代谢紊乱,使供给胃肠道的血流量下降,造成肠道缺血,导致肠道损伤,这也是热应激状态下牛奶产量下降的主要原因(占50%权重)[15,16]。热应激也会影响哺乳犊牛的生长发育,与环境温度20℃时相比,犊牛在热应激状态下(30~40℃)最初几天的采食量会降低12%[17],此时改善犊牛舍条件(尤其是下午)缓解热应激可显著增加采食量[18,19]。干奶期处于热应激状态会影响奶牛乳腺发育,进而降低下一泌乳期的产奶量。Tao等研究发现,当温度超过21℃时,对干奶牛采取风扇+喷淋的防暑降温措施,下一胎次泌乳早期的产奶量会增加5kg/d。热应激会降低新产牛的采食量,促使新产牛能量负平衡加剧,危害其健康和生产性能。与20℃相比,热应激状态下(30~40℃)奶牛的干物质采食量可降低28%,后续产奶量降低29%[20]。
1.2 热应激影响奶牛的繁殖性能
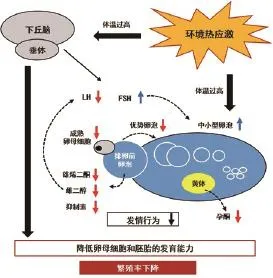
图3 热应激通过对下丘脑-垂体-性腺轴的调节而影响奶牛繁殖表现[21]
热应激会影响奶牛的内分泌系统(图3),进而影响奶牛的繁殖性能[21,22]。研究显示,配种前月平均每日最高THI与奶牛妊娠率存在高度相关(r=0.82)[23],配种后THI<55时无妊娠早期终止情况发生,超过65时妊娠早期终止率上升至8%,而THI超过69时则导致妊娠早期终止率达到12%[24]。
2 抗热应激性能奶牛选育的研究进展
2.1 通过奶牛红细胞钾水平选育耐热奶牛
奶牛红细胞钾浓度呈正态分布,符合数量性状分布规律,是评定奶牛耐热性的重要指标,耐热奶牛红细胞钾浓度较低。赖登明等建议以550mg/L作为选择标准[25];刘玉庆建议耐热母牛的选择标准是红细胞钾含量必须在800mg/L以下[26];史彬林则建议以550mg/L为标准,选择强度20%,以缩短耐热奶牛群选育周期[27]。对于红细胞钾的选择标准还要根据不同的品系、地理位置和育种目标等实际情况来确定。
2.2 通过杂交手段培育耐热品种(品系)
有学者认为,可以引入抗热能力较强的品种(如娟姗牛、瑞士褐牛)来改良本地奶牛,也可以引入热带地区的奶牛品系进行改良。国内有报道的是娟荷杂交模式,发现与荷斯坦牛相比,娟荷杂交牛耐热性能较强。刘庆华通过研究热应激对奶牛血液流变学、生化指标、免疫功能、抗氧化特性的影响,分析奶牛HSP70等热应激相关基因表达及其多态性与耐热性的关系,从耐热应激机理得出娟荷杂交牛比荷斯坦牛更能适应热带和亚热带地区高温高湿的气候条件[28]。梁学武等也在尝试培育含25%~75%娟姗血液的荷-娟耐热新品系[29]。
在通过杂交手段选育抗热应激品种方面,做得最成功的当属以色列。从20世纪40年代起,以色列以当地奶牛品种(大马士革牛、巴拉迪牛)为基础,开始引进国外公牛,但是从1963年以后就停止引种,基本使用国内的公牛冻精,并制定和严格执行国内育种改良计划,最终培育出了适应当地高热环境的高产品种——以色列荷斯坦牛。目前以色列95%的奶牛均为该品种牛,单产水平和抗热应激性能均表现优秀[30]。
2.3 通过分子标记辅助选择选育耐热品种(品系)
奶牛耐热性是数量性状,遗传力较低,常规育种效果不明显,而以分子标记辅助选择(MAS)为代表的分子育种可获得较快的遗传进展,已成为选育抗热应激奶牛的新突破点[31]。目前,在与奶牛抗热应激有关的遗传标记研究中,HSP70、S441、S463、S2011、S275、HSF1、ATP1A1和ATP1B2等基因可作为奶牛耐热性状的候选基因,相关研究进展如下:
2.3.1 HSP70基因
热休克蛋白70(HSP70)作为一种重要的分子伴侣,在热应激状态时,表达量迅速增加,能够缓解细胞所受伤害,可增强机体对热应激的抵抗力。陈强等采用PCR-SSCP方法对娟姗牛和荷斯坦牛HSP70基因5′-侧翼区多态性及基因型进行了初步检测和研究,认为其能否作为奶牛抗热应激选育的分子遗传标记还有待证实[32]。杜方磊等利用PCR-SSCP和测序技术研究了541头中国荷斯坦牛HSP70基因3'-非翻译区的多态性及其对奶牛产奶性状和耐热性状的影响,认为HSP70基因可以作为抗热应激的遗传标记[33]。Li等检测了890头中国荷斯坦牛HSP70基因3'-非翻译区的多态性,以及不同组织中HSP70基因的mRNA表达水平,进一步肯定HSP70基因可以作为抗热应激的奶牛选育分子遗传标记[34]。
2.3.2 S441和S463基因
胡雄贵等以产奶量数据变化和红细胞钾含量作为耐热评定指标,从高产奶牛群中筛选出耐热性状差异显著的10头高产荷斯坦牛作为试验材料,采用随机扩增多态性(RAPD)分子标记筛选方法,初步筛选出3个耐热分子遗传标记S441、S463和S93[35]。在该团队的后续研究中,又检测分析了湖南省213头种母牛和湖南省种公牛站以及北京、上海、南京、安徽等地46头种公牛,认为S441基因可以作为抗热应激候选基因[36]。谢江昂利用随机扩增多态性(PAPD)方法对筛选出的抗热应激分子标记进行序列分析,然后做序列特征化扩增区域(SCAR)转换,并对58头奶牛进行了检测,筛选出23头具有抗热应激分子标记的奶牛,发现有S441标记的奶牛对温热环境的适应能力明显好于无S441标记的奶牛,认为S441分子标记对奶牛抗热应激新品种的选育具有重要的参考价值[37]。胡雄贵等随机选取52头高产耐热荷斯坦牛(7~9月份产奶量平均下降率低于15%,红细胞钾含量低于468mg/L)和41头高产不耐热荷斯坦牛(7~9月份产奶量平均下降率超过36%,红细胞钾含量大于539mg/L),发现有无S441标记的母牛产奶量差异不显著,但在热应激条件下产奶量下降率差异极显著[38]。进一步研究发现,S441分子标记与抗热应激能力相关的各项生理生化指标均具有较强的相关性,证明S441可以作为一种奶牛抗热应激分子标记应用于品种选育和生产中[39]。刘彦等以S463为奶牛假定抗热应激的RAPD分子标记,在核心母牛场湖南省奶牛原种场测定不同世代的种母牛213头,在湖南省种公牛站以及北京、上海、南京、安徽等地测定种公牛46头,共检测出具有S463分子标记的母牛26头,公牛10头。发现有S463分子标记的奶牛在热应激期间不仅产奶量高,而且产奶量下降率非常低,认为S463可以作为抗热应激分子标记。由此推断,S441和S463基因抗热应激分子标记在耐热牛群中具有特异性[40,41]。
2.3.3 S2011和S275基因
张健选取耐热性能差异明显的荷斯坦牛,利用RAPD技术筛选出与耐热性能有关的分子标记,并结合奶牛的生理生化指标进行分析,推测引物S2011和S275可以作为筛选奶牛耐热性状的分子标记[42]。
2.3.4 HSF1 基因
李秋玲等[43]分析了638头中国荷斯坦牛HSF基因多态性与耐热性的关系,发现中国荷斯坦牛HSF1基因microRNA表达量与热应激反应存在显著相关,认为HSF1基因可作为选育高耐热性奶牛的新遗传标记应用于育种实践[44]。
2.3.5 ATP1A1和ATP1B2基因
南京农业大学王根林研究团队和山东农科院仲跻峰团队分别研究了中国荷斯坦牛ATP1A1和ATP1B2基因多态性与耐热性的关系及基因表达量,确认这两个基因可作为奶牛耐热性状的候选基因[45~47]。
2.4 通过全基因组检测分析筛选奶牛抗热应激分子标记
陆诚以30头中国荷斯坦牛和25头含25%以色列荷斯坦牛血统的杂交牛作为研究对象,采用Affymetix公司的基因表达谱芯片筛选了一批热应激相关基因,多组共有的表达量变化2倍以上的基因总共257个,其中基因表达上调139个,基因表达下调118个[48]。
美国弗罗里达大学Peter团队研究发现热应激条件下,奶牛的直肠温度高于39℃、呼吸频率大于60次/min时,奶牛的产奶和繁殖性能会受到影响[49]。通过监测在夏季热应激情况下奶牛的直肠温度、呼吸频率和出汗速度等与热应激密切相关的生理指标,利用包含了39 759个SNP位点的Illumina Bovine SNP50芯片对220头公牛的4 447头女儿进行全基因组检测分析,筛选了一批有潜力的热应激基因位点(表1)[7,8]。

表1 荷斯坦牛GWAS分析具有抗热应激生理特征(以直肠温度39℃为基准)最显著的20个基因位点
3 国外在抗热应激性能奶牛选育上的新发现
Olson等[50]研究了委内瑞拉的Carora牛发现携带少毛基因的个体夏季直肠温度较低,耐热性较强,Carora牛与荷斯坦牛的杂交后代耐热性增强,他们认为光滑短毛的控制HCT基因可以作为选育抗热应激奶牛的一个遗传标记。美国弗罗里达大学Peter团队[7,51]和新西兰家畜遗传改良公司(LIC)的Stephen团队研究发现[52],毛发光滑的奶牛比不光滑的奶牛抗热应激性能更强,在夏季热应激状态下产奶性能受损较轻,提出引起毛发光滑和长短的突变基因PRL基因及其受体PRLR基因可以作为选育抗热应激奶牛的关键遗传标记。
4 结语
奶牛耐热育种工作已经进入了新的历史阶段,除了一些基础性工作,还加强了分子标记辅助选择的耐热育种科研工作,为缩短世代间隔、提高育种效率提供了依据[53]。同时力图借助基因组检测手段快速筛选出一批具有抗热应激性能的基因位点,以期能有效应用于耐热品种(品系)的选育实践。
猜你喜欢
中国奶牛(2022年7期)2022-08-12
中国畜牧杂志(2021年1期)2021-01-20
三农资讯半月报(2020年17期)2020-09-26
中国畜牧杂志(2020年3期)2020-03-17
中国奶牛(2019年12期)2020-01-08
中国奶牛(2019年9期)2019-10-11
河南农业科学(2019年5期)2019-05-28
安徽农学通报(2015年4期)2015-07-30
食品工业科技(2014年13期)2014-12-16
