
同源四倍体矮牵牛的核型分析及有丝分裂观察
2019-03-07 06:26:00王媛花贾思振颜志明徐婷婷
贵州农业科学 2019年2期
魏 跃, 王 栋, 王媛花, 贾思振, 颜志明, 徐婷婷
(江苏农林职业技术学院,江苏 镇江 212400)
矮牵牛(PetuniahybridaVilm.)为茄科矮牵牛属草本花卉,夏秋季开花,色彩丰富,花期长,具有较高观赏价值,适用于花坛、花境及自然式布置,在世界各地广泛栽培,被喻为“世界花坛花卉之王”。魏跃等[1]曾利用秋水仙素处理二倍体矮牵牛(2n=2x=14),获得同源四倍体(2n=4x=28)新种质,其生长势强于二倍体,株高、茎粗、花瓣直径等性状变大,观赏性增加,显示多倍体矮牵牛具有较好的应用前景。近年来国内也有一些有关矮牵牛的细胞学研究,蔡华等[2]曾利用根尖进行核型分析,并证实二倍体矮牵牛染色体数目为2n=14,笔者等[3-4]也对二、四倍体矮牵牛花粉母细胞减数分裂过程进行过细胞学观察,但有关矮牵牛同源四倍体核型分析及有丝分裂等相关细胞学研究还未见报道。为此,笔者等对四倍体矮牵牛的核型及有丝分裂进行观察研究,确认其核型和有丝分裂的特异性,为同源多倍体细胞分类、染色体工程等方面研究提供借鉴。另外研究人员对观赏花卉进行核型研究时大多都以根尖、芽为材料[5-6],本试验采用幼嫩柱头进行核型分析,旨在为观赏花卉的细胞学研究提供新途径。
1 材料与方法
1.1 材料
供试材料矮牵牛四倍体品系09-5,由江苏农林职业技术学院花卉课题组提供,系利用秋水仙素诱导二倍体品种梦幻(购自浙江虹越花卉有限公司)经5~6代选育而成。
1.2 方法
雌蕊染色体制备参考张红梅等[7]方法进行,于夏季晴天早晨8∶00-10∶00点,取长度0.5~1 cm的幼小花蕾,置于0.002 mol/L 8-羟基喹啉水溶液中预处理3 h,蒸馏水冲洗3次,卡诺固定液(无水乙醇与冰醋酸体积比为3∶1)固定24 h,蒸馏水冲洗3次,0.075 mol/L氯化钾滴渗1 h,蒸馏水冲洗3次,1 mol/L HCl 60℃水浴解离6 min,蒸馏水冲洗3次,分别切取柱头、花柱和子房部位,用质量浓度为1%的醋酸洋红染色液充分染色后常规压片。观察3种部位的切片各30张,统计有丝分裂中期分裂相比率(中期细胞数目/观察细胞总数目×100%)。
核型分析:从四倍体材料中选5张中期分裂相良好的照片,采用Ikaros染色体图像分析软件进行染色体核型配对分析,并参照李懋学和陈瑞阳等[8-9]的植物核型分析标准进行分析测量,获得染色体参数,按STEBBINS[10]的分类标准进行核型分类。
2 结果与分析
2.1 雌蕊染色体观察最佳部位的筛选
在物镜(40×)同一视野下,柱头平均中期分裂相比率为3.8%,花柱和子房分别为1.1%和1.9%。表明四倍体矮牵牛雌蕊中柱头细胞处于有丝分裂中期的比率最高,适宜用于进行染色体观察与分析。
2.2 四倍体矮牵牛核型分析
从图1和表1可知,有丝分裂中期四倍体矮牵牛1~7号染色体绝对长度的变化范围在4.83~7.62 μm,相对长度的变化范围在11.3%~17.82%,相对长度系数(某染色体长度/染色体组的平均长度)[11]在0.79~1.25,其中1~3号染色体为中长染色体(M1),4~7号染色体为中短染色体(M2)。臂比(长臂/短臂)变化范围为1.04~2.43,着丝粒指数为29.12%~49.04%,1、4、6、7号染色体为中部着丝粒染色体(m),2、3、5号为亚中部着丝粒染色体(sm),随体在2号染色体。
四倍体矮牵牛终变期核型公式为K(2n)=4x=28=16m+12 sm(4SAT),染色体相对长度组成为2n=28=12M1+16M2。染色体长度比(最长/最短)比值为1.58,臂比大于2的染色体比例为28.57%,不对称系数为60.23%,为基本对称类型,按照STEBBINS[10]核型分类标准,矮牵牛核型分类为2A型属于较原始类型。
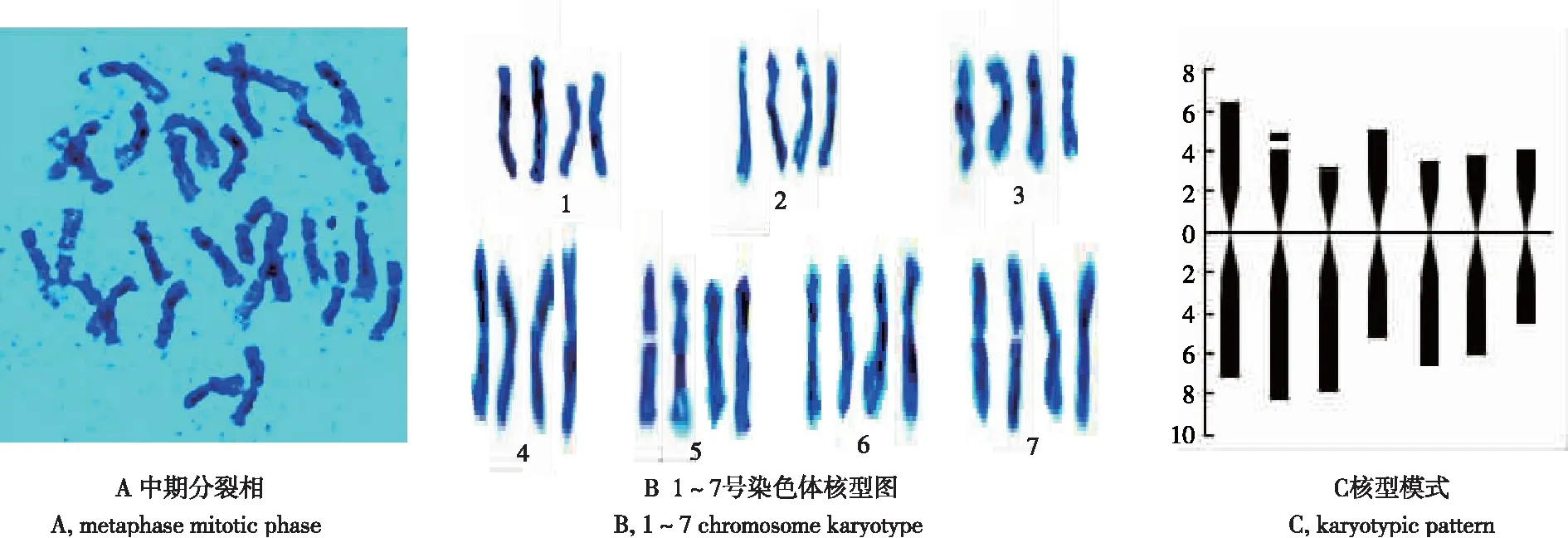
图1 同源四倍体矮牵牛染色体有丝分裂中期分裂相、核型及核型模式
Fig.1 Mitotic phase, chromosome karyotype and karyotypic pattern of autotetraploidP.hybridachromosomes at mitosis metaphase
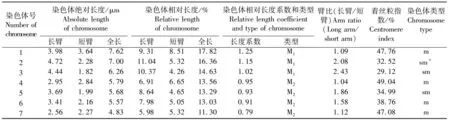
表1 同源四倍体矮牵牛染色体的核型参数
注:*为具随体的染色体.
Note:*, the chromosome with satellites.
2.3 有丝分裂的异常现象
同源四倍体矮牵牛雌蕊柱头体细胞有丝分裂过程中存在少量异常现象:在间期有微核仁出现(图2-1),细胞发生频率为3.17%;有丝分裂早中期姐妹染色单体部分交叉在一起呈链锁状发生染色单体交换(sister chromatid exchanges,SCE)(图2-2) 发生频率为0.8%;同源体细胞染色体配对成二价体(图2-3,2-4) 发生频率为3.7%;中期染色体排列在赤道板外(图2-5)、染色体丢失在细胞质中(图2-6)、后期出现落后染色体(图2-7)和染色体桥(图2-8 )合计发生频率为6.5%,此外还有中期和后期染色体出现凝集粘合(图2-9,2-10) 发生频率为2.9%。四倍体矮牵牛染色体数目应为2n=28,但少数细胞中出现2n=20(图2-11)和2n=24 (图2-12)非整倍体现象,发生频率约为1.7%。
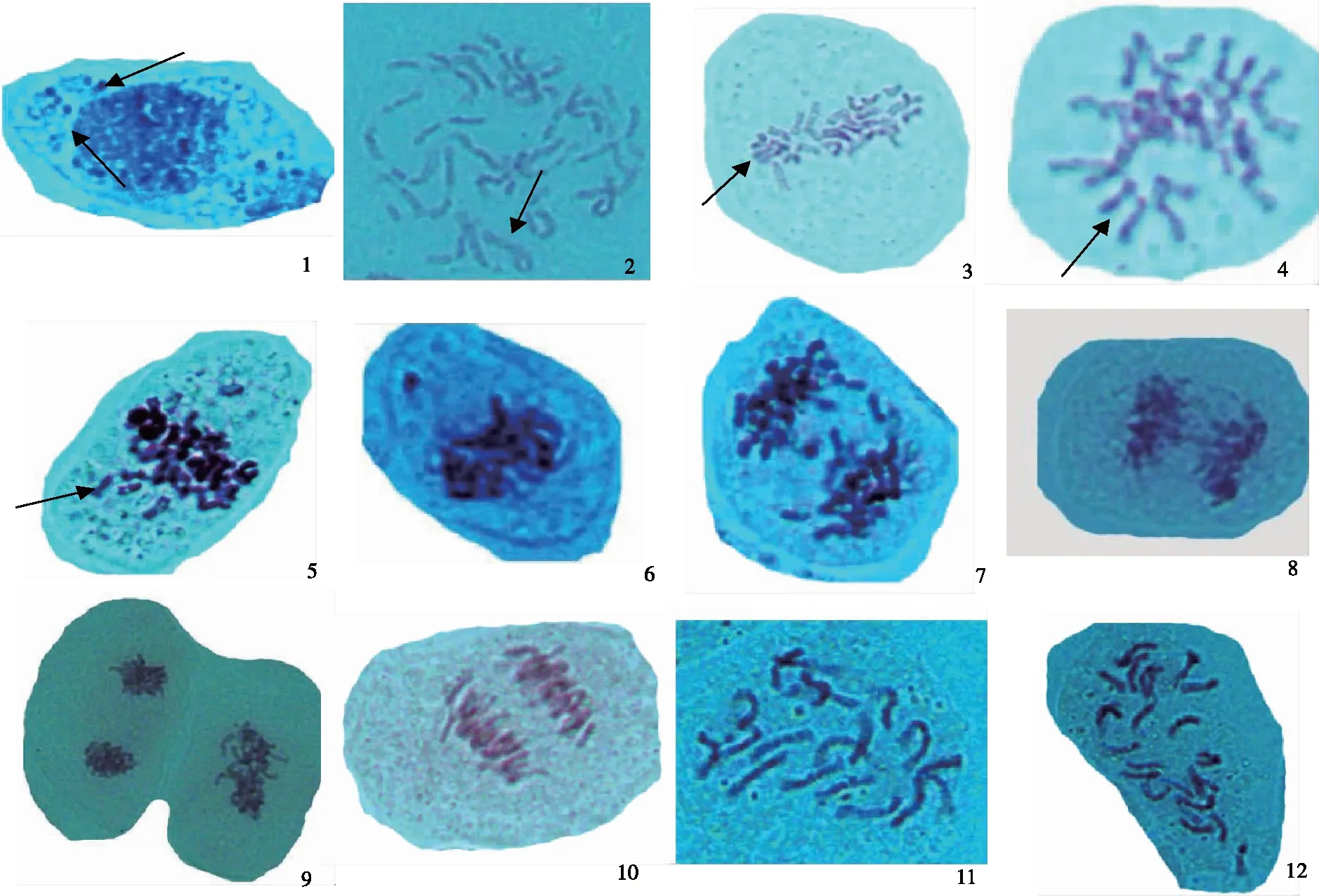
注:1为微核仁,2为姐妹染色单体交换(箭头示),3、4为体细胞同源染色体配对,5为赤道板外染色体,6为丢失染色体,为7落后染色体,8为染色体桥,9、10为凝集粘合,11、12为非整倍体。
Note:1.Micronucleolus; 2.Sister chromatid exchange; 3 and 4.Homoeologous pairing in somatic cell;5.Chromosomes outside the metaphase plate;6.Lost chromosome;7.Laggard chromosomes;8.Chromosome bridge;9 and 10.Chromosome condensation and bonding;11 and 12.Aneuploid.
图2同源四倍体矮牵牛异常有丝分裂及非整倍体(×1300)
Fig.2 Abnormal mitosis and aneuploid of autotetraploidP.hybridachromosomes (×1300)
3 结论与讨论
根尖是核型分析最常用的材料但对种子需求量较大,对结籽率低种子较少的作物不适用,且需要进行发芽培养,过程繁琐试验周期长。在理论上凡有细胞旺盛生长有丝分裂活跃的分生器官、组织都可以进行染色体观察,如陈劲枫等[12]用卷须代替根尖材料对黄瓜、菜豆和葡萄的染色体进行核型分析,蔡华等[13]利用新生子叶对牡丹进行核型分析,张红梅等[7]利用青花菜雌蕊子房进行染色体核型分析。本试验以幼嫩柱头为材料对四倍体矮牵牛进行核型分析,由于矮牵牛开花期较长,与根尖相比试验材料较多、取材方便且无需材料的培养准备,缩短了试验周期,简化了流程。染色体进行核型分析时取样大小也很重要,取材适宜其细胞分裂指数往往较高、中期分裂相也多。本试验同源四倍体矮牵牛幼小花蕾长度为0.5~1 cm时其中柱头直径为0.7~1.5 mm,此时柱头中有较多细胞处于有丝分裂旺盛期,中期分裂相较多,适合进行染色体研究。
核仁是真核生物细胞核特有的结构,并随着细胞周期发生分离和聚合。RISSO-PASCOTTO[14]在臂形草减数分裂前期I以外的各时期均观察到微核仁,认为微核仁是由前期的大核仁裂解而来,是基因发生自然突变所致。曹清河等[15]则认为细胞分裂周期中前核仁体由于无法聚合成大核仁而以游离状态存在于细胞中可形成微核仁。李悦有等[16]认为微核仁存在状态与细胞中蛋白质合成的旺盛程度密切相关。本试验结果显示,四倍体矮牵牛微核仁的发生频率平均为3.17%,高于二倍体的发生频率(0.92%),这可能与多倍体化后蛋白质合成旺盛程度增加相关。微核仁结构的存在有着复杂的生物学机制,其机理有待进一步探讨。
姐妹染色单体交换(SCE)是姐妹染色单体在相同位置发生等位片段交换,如发生不对等交换会导致基因扩增或丢失,SCE频率增加被认为是细胞环境中高浓度有害物质增加导致的,易导致染色体畸变[17]。仪慧兰等[18]使用NaCl溶液培养大麦幼苗导致根尖细胞有丝分裂指数下降,SCE频率增高且诱发有丝分裂染色体行为异常。本试验中观察到姐妹染色单体大片段多位点单体交换异常现象,可能与加倍后细胞内原有结构和功能改变, 导致DNA修复错误或抑制修复频率增加从而产生SCE[17]。
四倍体矮牵牛有丝分裂过程存在少量染色体凝集粘合、同源染色体配对、染色体丢失、赤道板外染色体、落后染色体、非整倍体等异常现象,而其中丢失和落后染色体、染色体桥可能是导致非整倍体产生的重要原因[19]。对同源四倍体异常有丝分裂的原因,张红梅等[7]认为可能是由于四倍体染色体加倍打破了二倍体中各种固有的平衡,造成内源激素不协调和生理生化过程不正常而造成的;STEBBINS等[10]认为同源四倍体有丝分裂不正常分离与染色体加倍造成特定基因位点纯合、隐性基因控制性状有关。四倍体矮牵牛体细胞有丝分裂产生异常现象的机理可能与上述因素相关但具体机理尚有待进一步探究。
猜你喜欢
科学导报(2023年60期)2023-09-11 06:38:25
农业科技通讯(2023年1期)2023-02-12 07:09:08
北京园林(2021年1期)2022-01-19 03:25:56
故事作文·低年级(2020年9期)2020-10-09 10:20:55
——矮牵牛
花卉(2020年15期)2020-08-11 09:22:24
现代园艺(2017年13期)2018-01-19 02:28:17
现代园艺(2017年18期)2017-02-04 15:45:39
中成药(2016年4期)2016-05-17 06:07:54
中国果业信息(2015年6期)2015-01-23 17:38:07
中学生物学(2014年4期)2014-09-25 01:28:11
