
高寒草甸地上净初级生产力与物种多样性及土壤养分的相关性
2019-03-07 06:30:28欧为友
贵州农业科学 2019年2期
欧为友
(青海省草原总站,青海 西宁 810016)
生物多样性是研究对象内植物、动物和微生物等物种所有基因,以及生物与环境相互作用的生态过程总和[1],生物多样性的作用在于控制生态系统稳定性、生产力及养分的动态变化[2]。可见,生物多样性对生态系统功能的实现具有重要作用。植物群落生物量作为生态系统生产力的基础,是生态系统功能的重要表现形式,探讨植物群落生产力与生物多样性的关系,对于阐述植物群落生产力对生态系统功能的作用具有重要意义[3]。目前,关于生物多样性与植物群落生产力的关系已成为生态系统的研究热点之一[4],已有的研究结论存在正相关[5]、负相关[6]、单峰曲线关系[7]和不相关4种观点。关于物种多样性与植物群落生产力的关系尚存疑问,如物种多样性是否是影响植物群落生物量的主要因子,物种多样性、群落生物量及土壤养分间的关系等。为此,以典型嵩草草甸为研究对象,研究其地上净初级生产力与物种多样性及土壤养分的相关性,以弄清植被群落物种多样性与地上净初级生产力的关系,为生产应用提供参考依据。
1材料与方法
1.1研究区概况
玉树县位于青藏高原东部,地处玉树藏族自治州东部,地理位置东经95°41′40″~97°44′34″,北纬33°44′44″~33°46′44″,总面积1.57万 km2,全县地形以高原为主,由西向东呈不断降低走势,平均海拔4 493.4 m,属典型的高原高寒气候,全年无四季之分,只有冷暖两季之别,冷季长达7~8个月,年均温2.9℃,年降水量487 mm。
1.2试验方法
1.2.1样地选择与处理在研究区清水河镇文措村阳坡面坡中(坡度20o)随机选取10 m×10 m的5个固定样地,样地优势牧草分别为小嵩草(Kobresiapygmaea)、蔵嵩草(Kobresiatibetica)、线叶嵩草(Kobresiacapillifolia)、矮嵩草(Kobresiahumilis)和小嵩草-杂类草草甸,每种类型样地为1个处理,记为T1~T5,样地海拔4 214~4 377 m(表1)。
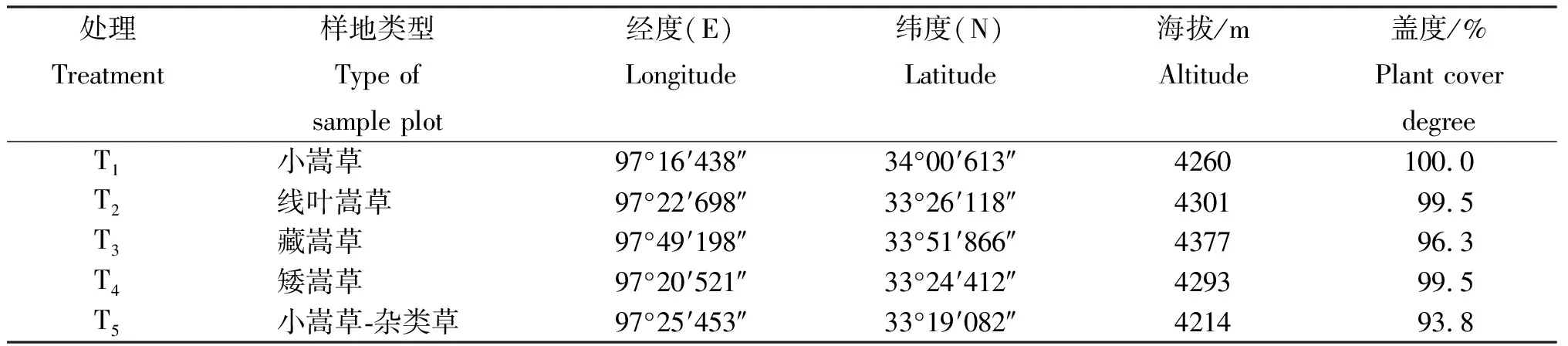
表1 5个固定样地的基本情况
1.2.2地上植物量调查及土样采集
1) 地上植物量。在每个样地随机选取1 m×1 m的3个小样方,于2008年7月底(高寒草甸植被生长旺盛期)对小样方地上植物量进行调查,调查内容包括植物种类、盖度、高度和密度,并于室内准确鉴定植物标本,统计科、属、种及其组成。在样方内,对角线设2个0.25 m2(0.5 m×0.5 m)的观测样方,剪去植物地上部后,莎草科牧草、其他杂类草分别称重量,两者之和即为地上植物量。
2) 土样。不分层采集0~30 cm土壤,装入无菌袋,带回室内测定土壤养分的含量,包括全氮、全磷、全钾、碱解氮、速效钾、有效磷和有机质。全氮(TN)采用半微量凯氏定氮法,碱解氮(AN)采用碱解蒸馏法,全磷(TP)采用钼锑抗显色法,有效磷(AP)采用碳酸氢钠法,全钾(TK)采用NaOH熔融-火焰光度法,速效钾(AK)采用乙酸铵-火焰光度法。
1.3物种多样性
物种多样性指标包括丰富度指数(R)、群落多样性指数(H)、均匀度指数(J)和生态优势度指数(I)[8]。
R=S
I=(相对高度+相对盖度+相对密度)/3
式中,Pi为种i的相对重要值(相对高度+相对盖度+相对频度)/3,S为种i所在样方的物种总数。
1.4植被类群
植物类群分优良牧草和杂类草,禾本科、莎草科归入优良牧草一类,乳白香青(Anaphalislacteal)、火绒草(Leontopodiumleontopodioides)、微孔草〔Microulasikkinensis(Clarke)hen sl〕、星状风毛菊(SaussureastellaMaxim)、秦艽(Gentianamacrophylla)、唐松草(Thalictrumalpinum)、龙胆(Gentianaspp.)、蒲公英(Taraxacummongolicum)等则归入杂类草。
1.5数据处理
采用Excel 2010对数据进行处理, 利用SPSS 19.0进行方差分析、相关分析和主成分分析。
2结果与分析
2.1地上植物量及多样性指数
从表2看出,不同草地类型地上植物量及多样性指数存在差异。T3(蔵嵩草草甸)地上植物量和优良牧草植物量均最大,分别为268.33 g/m2和137.25 g/m2,均显著高于其他类型草甸(P≤0.05),T5(小嵩草-杂类草)地上植物量和优良牧草植物量最小,分别为167.33 g/m2和13.67 g/m2。不同样地群落多样性指数(H)为2.20~2.71,T5最大,为2.71,显著高于其余样地(P≤0.05);均匀度指数(J)为0.86~0.91,不同样地间变化不大,T1~T2与T3~T5差异显著(P≤0.05);丰富度指数(R)为16.33~28.67,T1最大,为28.67,显著高于其余样地(P≤0.05)。
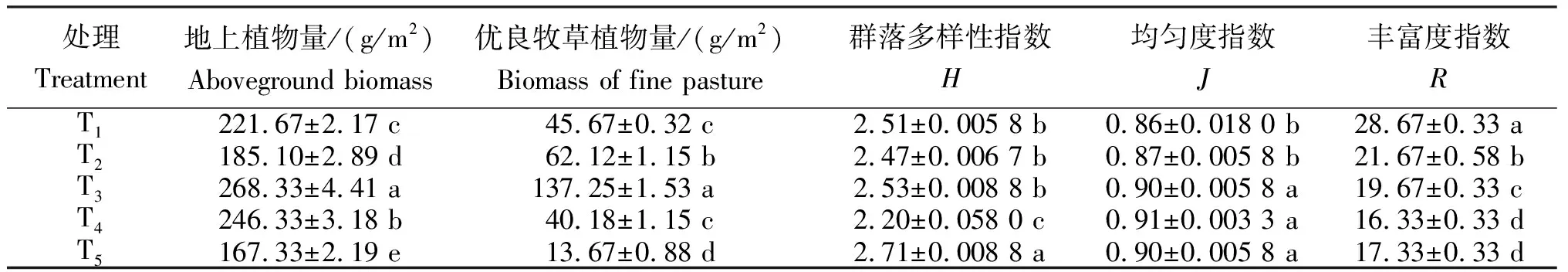
表2不同样地的地上植物量及生物多样性指数
注:同列不同小写字母表示差异显著(P≤0.05),下同。
Note: Different lowercase letters in the same column indicate significance of difference atP≤0.05 level. The same below.
2.2不同样地土壤养分的含量
从表3可知,不同样地土壤养分的含量存在差异。全氮含量,T3最高,为7.27 g/kg;T5其次,为6.76 g/kg;T4最低,为2.96 g/kg;T3、T5与T1、T4间差异显著(P≤0.05)。全磷含量,T1最高,为3.22 g/kg;T3其次,为2.47 g/kg;T5最低,为1.68 g/kg;T1、T2、T3与T4、T5间差异显著(P≤0.05)。全钾含量,T4最高,为23.40 g/kg;T1其次,为22.36 g/kg;T2最低,为19.74 g/kg;T1、T4与T2、T3、T5间差异显著(P≤0.05)。有机质含量,T3最高,为158.81 g/kg;T5其次,为141.70 g/kg;T4最低,为43.97 g/kg;T3与T1、T2、T4间差异显著(P≤0.05)。碱解氮、有效磷和速效钾含量分别以T3、T3和T5最高,T4、T1和T1最低。
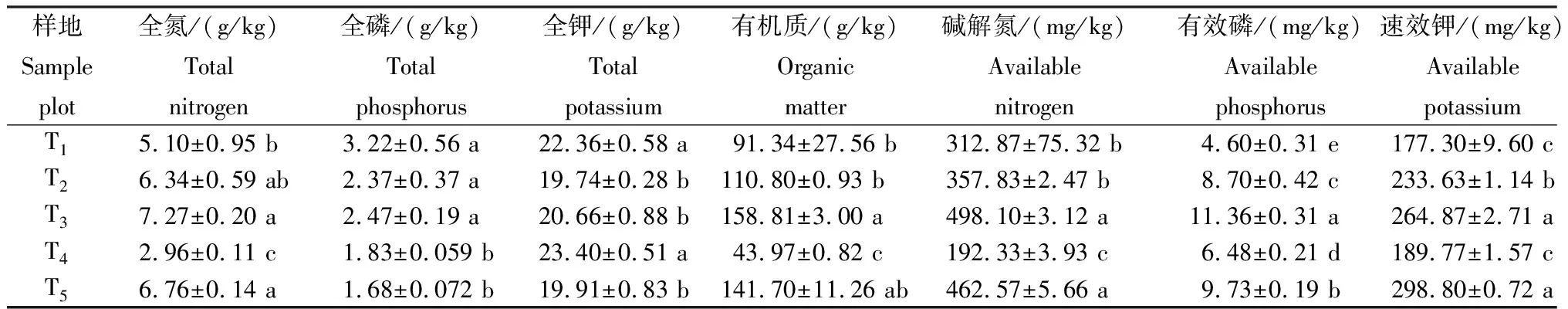
表3不同样地土壤的养分含量
2.3土壤养分及植物量与多样性的相关性
从表4看出,全氮、有机质、碱解氮和速效钾与群落多样性指数呈极显著负相关(P≤0.05),全氮、全磷、全钾、有机质、碱解氮、有效磷和速效钾与均匀度指数相关性不显著(P>0.05)。丰富度指数与有效磷呈显著负相关(P≤0.05),而与全氮、全磷、全钾、有机质、碱解氮和速效钾相关性不显著(P>0.05)。多样性指数与地上植物量呈极显著负相关(P≤0.01),而与优良牧草植物量不呈显著相关(P>0.05)。优良牧草植物量与有效磷呈显著正相关(P≤0.05)。
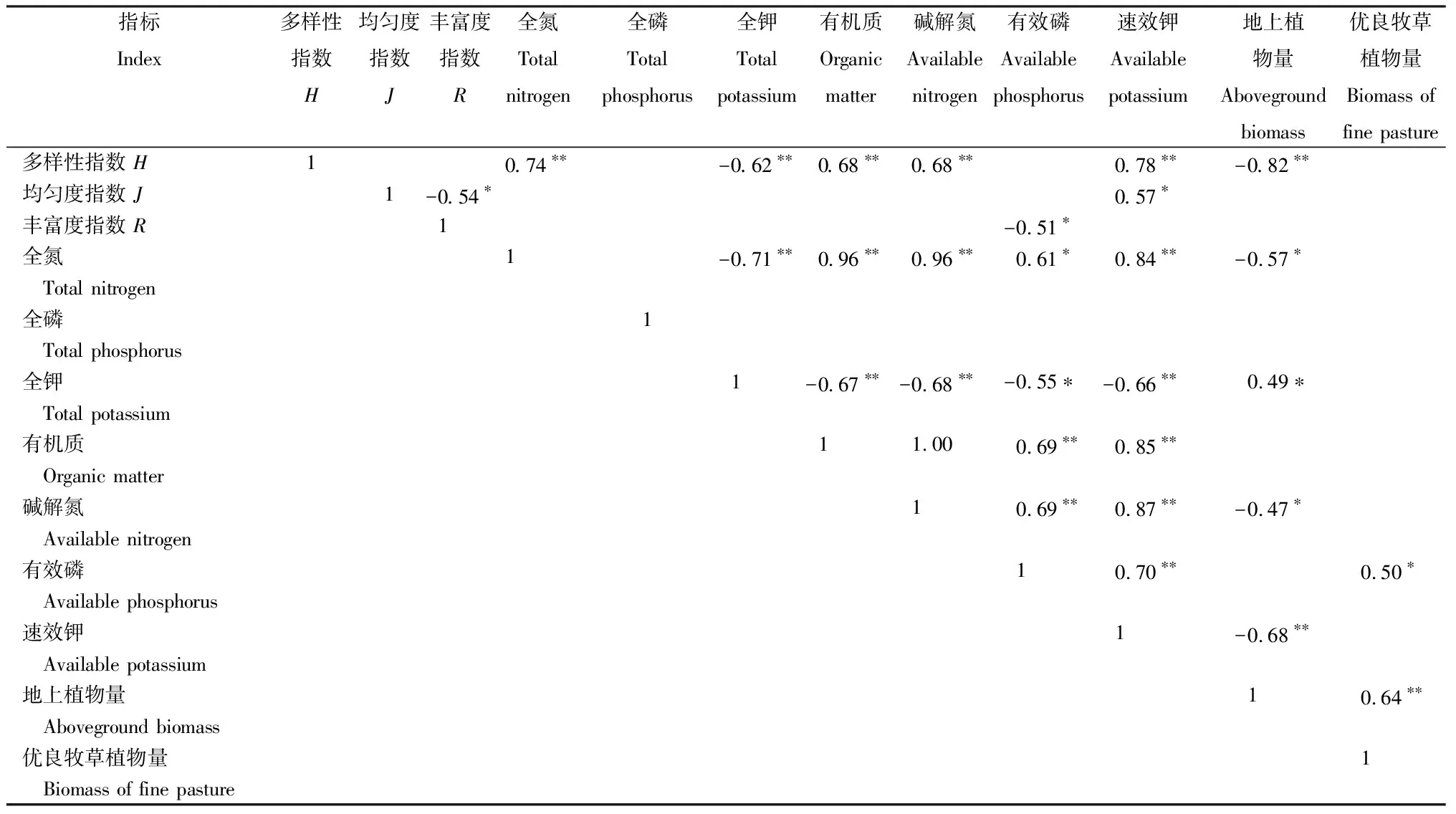
表4 土壤理化性质、植物量和生物多样性指数的相关性
注:*为显著相关(P≤0.05),**为极显著相关(P≤0.01)。下同。
Note: * and ** indicate significance of difference atP≤0.05 andP≤0.01 level respectively。The same below.
2.4植物量、物种多样性指数及土壤化学性质主成分分析
从表5看出,经样地主要因子主成分分析的前3个主成分的特征值均大于1,累计贡献率达87.26%。其中,第1主成分的方差贡献率达49.76%,与有机质、全氮、碱解氮、有效磷、速效钾和多样性指数呈显著正相关(P≤0.05),与全钾呈显著负相关关系(P≤0.05);第2主成分与丰富度指数、土壤全磷呈显著正相关(P≤0.05),而与均匀度指数呈显著负相关(P≤0.05)。
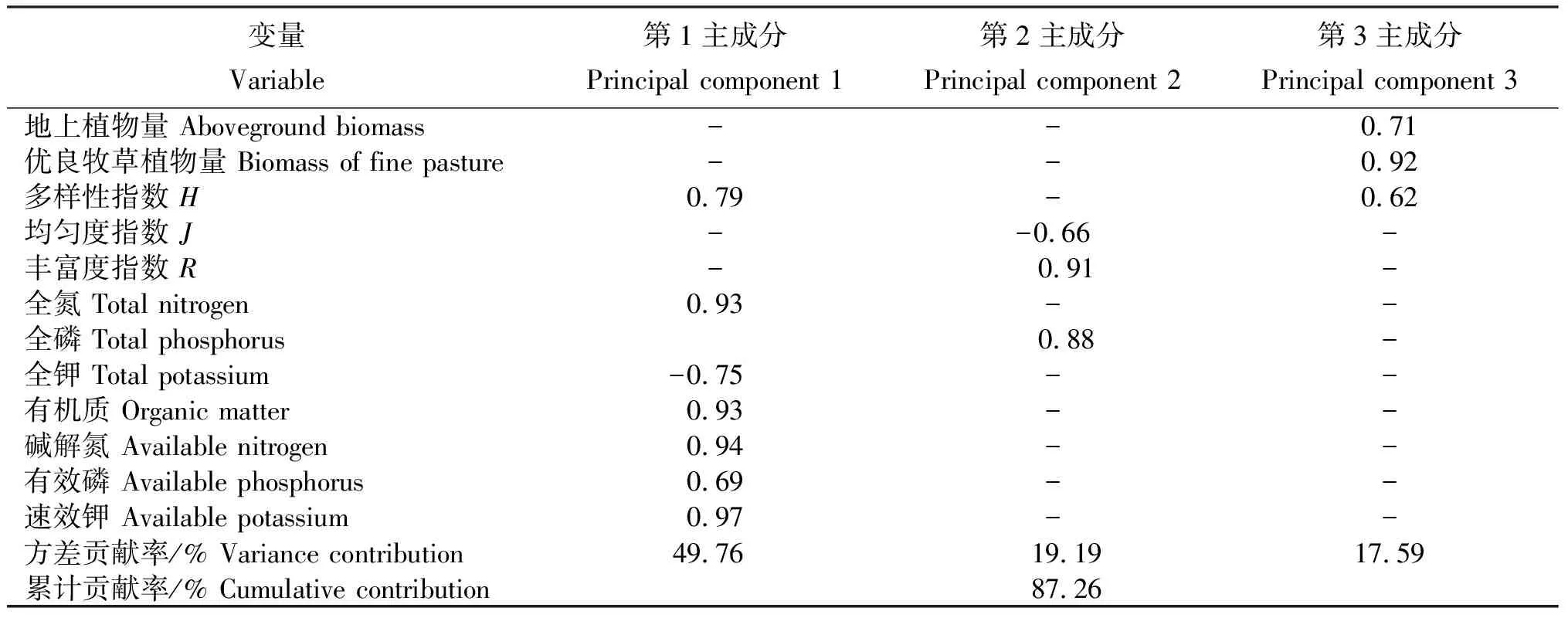
表5样方指标因子主成分分析矩阵
注:-为无数据。
Note:-, no data.
3结论与讨论
3.1植物群落多样性、土壤养分和地上植物量对植物群落的影响
植物群落多样性指数、土壤养分和地上植物量是植物群落的3个主要组成部分,三者相互制约、相互作用,共同决定植物群落的变化。一方面,异质性群落环境,即不同的植物群落多样性是土壤养分差异的重要因素。刘晓琴等[9]通过比较不同封育年限的群落组分和物种多样性变化指出,封育后,凋落物的分解引起土壤养分的变化是导致物种多样性降低的重要原因,而凋落物的产生就是由植被群落结构决定的。另一方面,资源供给率[10]也是影响植物群落物种多样性与生产力的重要因子,而这种资源主要体现在土壤养分上。陈宁等[11]通过分析退化高寒草甸群落生产力和物种多样性指出,随着退化程度加剧,群落优势种发生变化,功能群变化导致土壤C、N含量非线性降低引起植物群落多样性变化。有研究发现,在健康群落中,物种多样性和土壤养分的综合作用是导致植物群落差异的主要因子,原因在于较低肥力的土壤中,群落多样性指数受有限养分的制约[10];而健康的植被群落结构较充足的土壤养分某种程度上可能弱化了种间竞争的作用,导致物种数随土壤养分的增加而增加,降低了不同物种对资源利用的差异,使得物种多样性对地上植物量的贡献未能完全表现。同样,杜家丽[12]在分析施肥、刈割等干扰条件对高寒草甸的影响也指出,在施肥条件下,由于低温环境的影响,高寒草甸土壤N、P养分的矿化率较低,可利用N、P含量仅占土壤养分的1%左右,限制了植物的生长,而施肥能有效解除土壤的养分限制,导致群落地上净初级生产力增加。
3.2植物群落多样性指数、土壤养分和地上植物量的关系
分析植物群落多样性指数、土壤养分和地上植物量的关系主要从自然和人工干扰的角度展开。在自然条件下,退化、气候变化和环境等是主要方面。大量研究表明,随着退化程度加剧,由于杂类草的比重增加,种间竞争加剧[11]以及土壤较强的抗退化能力等[13],土壤碳含量、体积含水量和砾石质量比对地上生物量无显著影响,而土壤碳、氮含量与物种多样性指数呈显著负相关。在大尺度下,耿元波等[14]在分析近50年气候条件对高寒草甸生产力的影响发现,降水量与生产力显著相关,而与温度则相关性不显著。在小尺度下,坡向、海拔等都通过对局部小气候产生影响,作用于植物群落结构。在人工干扰条件下,施肥、刈割和放牧等都产生不同的作用。杜家丽[12]研究表明,刈割和施肥都能增加群落地上植物量,但功能群多样性对地上植物量的贡献大于物种多样性。然而,在放牧干扰下,由于土壤紧实度的作用,土壤物理因子为主的土壤因子是引起地上植物量和物种多样性变化的主要原因[15]。显然,干扰条件往往主导植物群落多样性指数、土壤养分和地上植物量的变化。研究结果表明,在自然条件下,有机质、全氮和速效养分与地上植物量呈显著正相关,说明土壤养分是影响地上植物量的主要因子。进一步发现,土壤养分对植物群落结构的影响,引起群落多样性指数的变化,植物群落多样性Shannon-Wiener指数和土壤养分的共同作用造成了不同高寒草甸类型的变化,这一推断从贡献度就可以看出,验证了杨兆平等的研究结果[16-18]。至于植物多样性和功能群多样性的相对贡献程度则有待于进一步研究。
综上所述,通过探讨不同高寒草甸类型群落多样性指数、土壤养分和地上植物量的变化发现,多样性指数与地上植物量呈显著负相关,有机质、全氮和速效养分与多样性指数呈显著正相关,土壤养分和多样性指数对高寒草甸的影贡献率达49.76%,其结果反映出高寒草甸土壤养分是影响地上植物量差异的最主要因子,高寒草甸群落多样性指数和土壤养分共同影响高寒草甸的植被特征。
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
青海草业(2022年2期)2022-07-23 09:34:58
现代园艺(2021年23期)2021-12-01 07:47:44
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
活力(2019年21期)2019-04-01 12:17:10
现代农业(2016年3期)2016-04-14 12:35:28
土壤与作物(2015年3期)2015-12-08 00:47:01
河池学院学报(2014年5期)2014-02-27 13:37:18
