
马铃薯抗寒种质资源分类及其利用
2019-03-05 09:12:26彭晓君梅文祥宋波涛
中国马铃薯 2019年1期
康 黎,彭晓君,陈 琳,涂 卫,梅文祥,宋波涛
(华中农业大学园艺林学学院/华中农业大学园艺植物生物学教育部重点实验室/农业农村部马铃薯生物学与生物技术重点实验室,湖北 武汉 430070)
现在广泛种植的马铃薯品种均为马铃薯普通栽培种(Solanum tuberosum),几乎都不耐低温霜冻,没有低温驯化能力,仅有‘郑薯5号’等少数几个品种具有极其微弱的抗寒能力[1],说明栽培种内不存在抗寒遗传变异,这使得从普通栽培种内部挖掘抗性材料可能性极低。目前,虽然也有一些具有抗寒能力的栽培品种被报道,如‘Alaska Frostless’[2]等,但离广泛推广应用还有很长一段距离,因此,加强对马铃薯抗寒种质资源的研究,引入新的抗性种质将有助于中国马铃薯产业的持续健康发展。
1 马铃薯抗寒种质资源
马铃薯的抗寒性不仅取决于其种的属性,也与低温驯化相关。低温驯化(也称冷驯化)是将植物置于低温环境中(0~12℃),植物体内发生一系列生理生化变化,使植物产生适应性反应,以提高对低温抵抗能力的过程(Cold acclimation,CA)[3]。马铃薯栽培品种的遗传背景十分的狭隘且几乎没有抗寒能力和低温驯化能力,但马铃薯野生种资源丰富,按Hawkes[4]的分类标准,可分为21个系235个种,栽培种数量为7个,野生种数量为228个,其中很多野生种存在耐低温霜冻的资源,而且还有一些资源具有较强的低温驯化能力。
Li[5]对60个野生种进行梯度降温处理,处理时间为24 h,结果表明S.acaule大部分的致死温度可达-6.5℃,抗寒能力最强,其次是S.boliviens、S.megistacrolobum和S.commersonii,致死温度可低至-5.0℃,此外,S.demissum、S.torflapanum、S.sanctae-rose存在致死温度为-4.0℃的无性系。Palta和Li[6]通过同样的方法,也发现S.acaule、S.multidissectum、S.demissum、S.megistacrolobum、S.sanctae-rose、S.commersonii、S.vernei存在有无性系能够忍耐-5℃以下低温的植株。Chen和Li[7]将结薯马铃薯种的抗寒能力和冷驯化能力进行了分类(表1),基于有无低温驯化能力和植株的抗寒能力将马铃薯的抗寒性分成5类:抗寒型马铃薯种并能够低温驯化;抗寒型马铃薯种不能够低温驯化;霜冻敏感型马铃薯种能够低温驯化;霜冻敏感型马铃薯种不能够低温驯化;霜冻敏感型马铃薯种。Vega和Bamberg[8]通过田间自然霜冻法,对101个种2 635份材料的抗寒性鉴定,并将抗性分为7个等级(0~6,0代表没有受到损伤,6代表植株死亡),结果显示S.acaule、S.albicans抗寒能力最强,等级为0;其次是S.commersonii,等级为0.05;此外,S.demissum、S.megistacrolobum、S.paucissectum、S.multidissectum、S.sanctae-rose 5个野生种的等级介于0.5~1。Luthra等[9]于早春时候将21个种共86个材料的植株置于室外生长7 d后,统计叶片的损伤率鉴定其抗寒能力,并划分10个等级(1~10,1表示叶片损伤率低于10%,10表示叶片损伤率高于90%),其中有18个材料的等级为1,分布在S.goniocalyx、S.arnezii、S.demissum、S.spegazzinii中。赵喜娟[10]通过人工冷冻法对18个野生种的97份材料进行抗寒性评价,处理温度-3.5℃,处理时间24 h,评级方法与田间自然霜冻法相同,结果表明S.acaule、S.bukasovii、S.commersonii等野生种均具有较强抗寒能力,评级介于0~1;而S.chomatophilum、S.demissum、S.megistacrolobum和S.acaule等野生种中也有部分植株抗寒能力较强,其中S.acaulel中77%的植株评级小于2。在所有具有强抗寒能力野生种当中,来自无茎系的野生种和S.commersonii被认为是抗寒能力最强的2类野生种,已有研究表明来自无茎系的野生种S.acaule在驯化前后分别能忍受-6和-9℃的低温,而S.commersonii驯化前后分别能忍受-4.5和-11.5℃的低温[7],是被研究和利用最多的耐冻性资源,以下将就这2类野生种的利用情况进行综述。
2 马铃薯无茎系野生种分类及其利用
2.1 无茎系野生种分类
S.acaule、S.×aemulans、S.albicans等无茎系野生种早在20世纪50年代就已经被发现,但直到1990年才由Hawkes最早提出了无茎系的概念,在其分类体系中,无茎系作为马铃薯组的一个系,包括有5个野生种。但早在无茎系概念还没有提出来,分类学家对上述野生的分类和命名产生了分歧,直到现在分歧仍然存在。总体来说,大部分分类学家对马铃薯无茎系内野生种划分的分歧主要是区别系内种与亚种之间的划分,特别是S.albicans和S.×aemulans的划分上。早期的分类中S.albicans则是作为S.acaule的亚种S.acaule var.albicans或 S.acaule ssp.albicans存在;而S.×aemulans一直以来都是作为S.acaule的亚种而被人所熟知,但Spooner等[11]却认为S.acaule ssp.aemulans和S.×indounii只是野生种S.×aemulans的2种不同倍性,相关的分类差异见表2[7,11-14]。
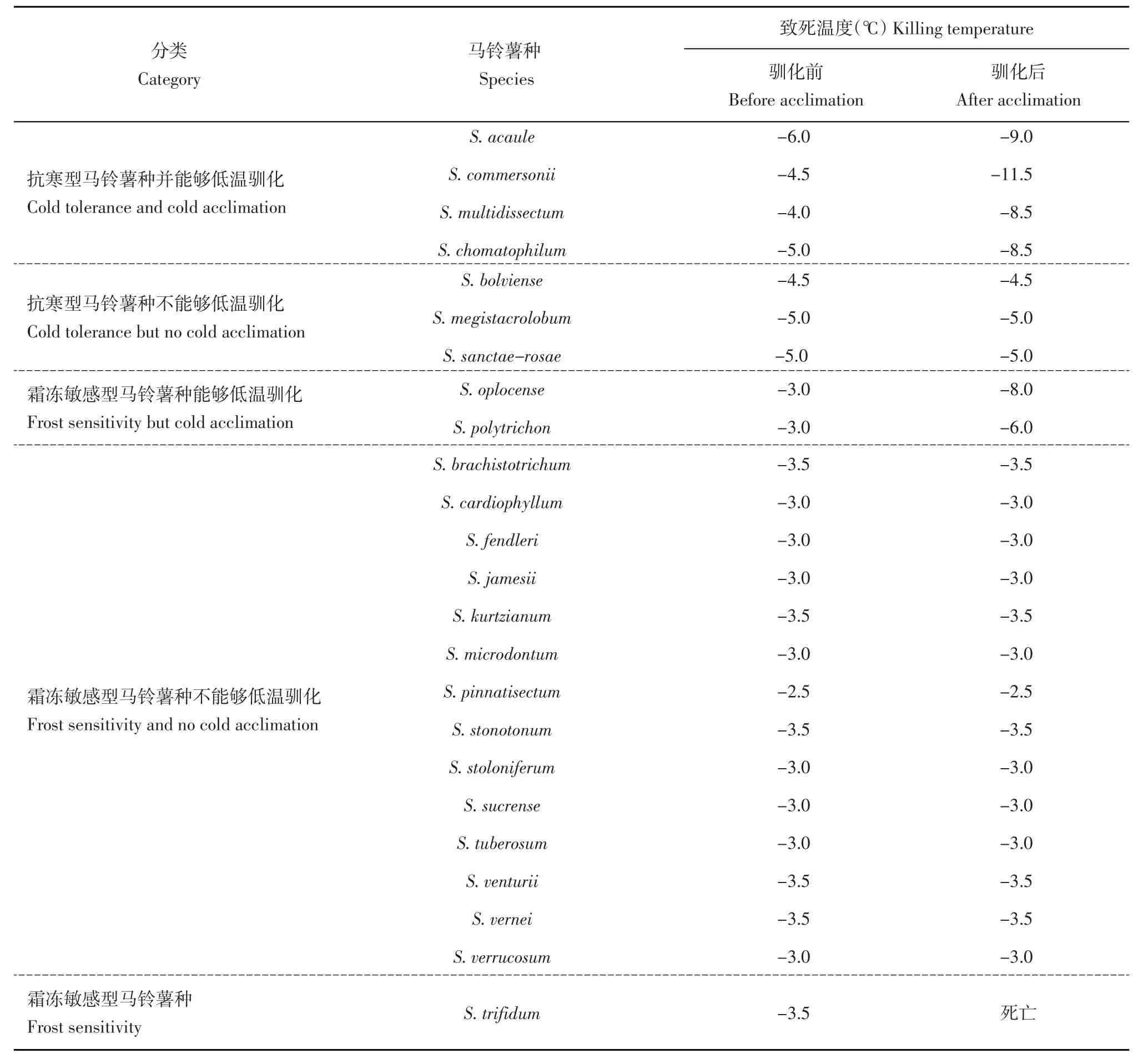
表1 马铃薯种的抗寒性和冷驯化能力分类Table 1 Classification of tuber-bearing Solanum species in terms of cold tolerance and cold acclimation
2.2 无茎系野生种起源与演化
马铃薯无茎系野生种主要分布在南美洲安第斯山脉,分布在海拔3 500~4 500 m[14]。无茎系的起源和演化是一个非常复杂的关系,是多个野生种相互渗透的过程。
早期的研究人员通过一些形态学的观察和杂交试验等研究发现S.acaule的植株形态与S.megistacrolobum非常相似,而叶片形态则与S.canasense非常类似,而且S.acaule的繁殖习性也与上述2个野生种有共同之处,这表明S.acaule的形成过程中可能有S.megistacrolobum与S.canasense杂交这个环节的参与[15]。Nakagawa和Hosaka[16]通过对上百份野生种的核DNA和叶绿体DNA进行RFLPs分析,发现S.acaule的叶绿体DNA在序列结构与S.megistacrolobum、S.sanctae-rose的非常相似,都属于C类型;S.acaule的核DNA与S.megistacrolobum、S.sanctae-rose同样具有极高的相似性,在进化树上距离非常近,都属于Cluster 1类型,由此推测S.acaule的起源可能和这2个野生种相关。此外,还有研究表明二倍体野生种S.boliviens可能在S.acaule形成过程中发挥了重要的作用。Goncarik[17]通过检测数百份野生种和栽培种中叶绿体简单重复序列(cpSSR)类型并对各个种血缘进行分析,发现S.acaule和S.boliviens的cpSSR具有高度的相似性,由此推断S.boliviens可能通过杂交的方式直接参与了S.acaule形成过程。上述研究表明,在S.acaule的形成过程中,二倍体野生种S.boliviens、S.megistacrolobum、S.canasense都可能参与其中,其中S.megistacrolobum和S.boliviens可能性最大;实际上,也有分类学家认为S.megistacrolobum,S.sanctae-rose和S.boliviens其实是同一个物种,即S.boliviens[11],这也进一步证明了马铃薯种的形成其实是野生种在进化过程长期相互渗透的过程。

表2 马铃薯无茎系野生种分类Table 2 Classification of Solanum acaule Juz
野生种S.acaule也与无茎系其他种的形成有着密切的联系。S.×aemulans可能是S.acaule ssp.acaule和二倍体野生种S.megistacrolobum的天然杂交后代[18],且因为S.megistacrolobum能产生2n配子,而导致了S.×aemulans存在三倍体和四倍体2种不同的倍性;S.albicans则有可能是S.acaule ssp.punae和一个与无茎系亲缘关系较远的二倍体杂交后经过天然加倍而形成的[4]。
除了参与无茎系其他野生种的形成,S.acaule还可能参与了其他野生种和原始栽培种的形成。S.×brucheri可能是S.acaule ssp.acaule和二倍体野生种S.infundibuliforme的天然杂交后代[19]。原始栽培种S.×juzepczukii和S.×curtilobum可能也会有S.acaule的血缘[20-22],具体演化的过程为:S.×juzepczukii为S.acaule与S.stenomum的天然杂交后代,S.×curtilobum则为S.×juzepczukii的2n配子与S.tuberosum ssp.andigenum的天然杂交后代(图1)。
2.3 无茎系野生种在抗寒育种中的应用
马铃薯无茎系野生种中拥有丰富的耐低温霜冻材料,是马铃薯抗寒育种的重要种质资源,S.acaule则是无茎系中利用最为普遍的野生种,是目前已知抗寒能力最强的野生种之一。英国早在20世纪40年代就试图利用S.acaule来培育抗寒马铃薯品种,但收效甚微。从目前已有的报道来看,利用S.acaule选育出来的并且具有抗寒能力品种只有‘Alaska Frostless’[2],该品种的选育过程中有一个加倍后的S.acaule无性系参与了其中,其经过驯化后LT50可以达到-4.6℃。除了‘Alaska Frostless’,其余的均为一些可用于抗寒育种的中间材料。Estrada[23]用四倍体的S.acaule和二倍体的S.phureja以及四倍体的普通栽培种依次杂交后成功获得了杂种后代,有27%的后代具有较强的抗寒能力;稍晚一些,Estrada[24]又直接从S.acaule和S.phureja的杂种后代中鉴定了一个四倍体acaphu,acaphu可直接和四倍体栽培种杂交产生四倍体的后代,且后代中某些植株能够忍耐-5℃低温2 h不死;Bamberg等[25]通过用加倍后S.commersonii作为桥梁种,成功获得了ACL-CMM-TBR 3者的种间杂种,该种间杂种具有一定的抗寒能力和驯化能力,可用于后续的抗寒育种。
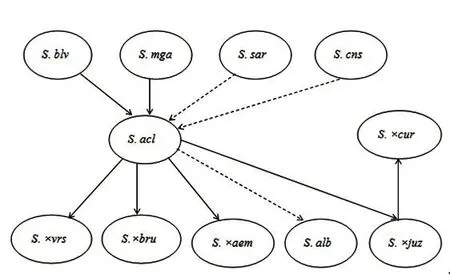
图1 无茎系的起源与演化Figure 1 Origin and evolution of acaule Juz
2.4 马铃薯野生种在抗病育种中的应用
除了抗寒能力之外,马铃薯无茎系野生种中还拥有丰富的抗马铃薯病毒材料,是马铃薯抗病毒育种的重要种质资源,特别是野生种S.acaule。据不完全统计,野生种S.acaule中存在有抗癌肿病,抗马铃薯X病毒(Potato virus X,PVX)、马铃薯卷叶病毒(Potato leafroll virus,PLRV)、马铃薯纺锤块茎类病毒(Potato spindle tuber viroid,PSTVd)的材料[26,27]。
野生种S.acaule在马铃薯病毒抗性育种中的利用已经非常的广泛,已经获得了非常多育种中间材料和一些品种。有研究用7XY.1、Bolona等四倍体的普通栽培种和S.acaule杂交,授粉第2 d再用S.phureja(IVP35)进行二次授粉,待第一次授粉后14~27 d采用胚抢救(Embryo rescue),成功地获得了3个可育的四倍体杂种,这3个杂种均检测出了PSTVd抗性,而且在随后的研究中表明,这3个四倍体均具有优良结薯表型[28,29];Watanabe等[30]通过染色体加倍和多次回交的方式,获得了含有S.acaule血缘的四倍体杂种,这些杂种中有部分植株检测到了PLRV和PSTVd抗性;还有研究人员通过将S.acaule和普通栽培种进行体细胞融合,获得一批倍性不一的体细胞杂种,这些体细胞杂种中均存在有PVX极端抗性植株,且有些植株结薯性状非常优良,可作为育种中间材料[31-33]。此外,波兰科学家利用染色加倍将S.acaule的一个基因型变成同源八倍体后与普通栽培种杂交后进行多次回交,成功的获得回交杂种MPI44.106/10,该杂种被用于抗PVX的育种亲本,并用其选育出了多个抗PVX的品种,典型的如‘Barbara’[34];中国黑龙江省农业科学院马铃薯研究所同样通过加倍的方法将野生种S.acaule的PVX抗性导入到了普通栽培种当中,并通过3~4代轮回选择后成功获得了多份抗PVX的育种亲本[35]。
3 S.commersonii及其在抗寒育种中利用
3.1 野生种S.commersonii概述
S.commersonii隶属马铃薯组孔目松系,为二倍体野生种,EBN(Endosperm balance number)=1,主要分布在阿根廷,乌拉圭,巴西等国家,生长于安第斯山脉海拔400 m左右的区域。在Hawkes[4]的分类体系中,S.commersonii分为2个亚种,分别是S.commersonii ssp.commersonii和S.commersonii ssp.malmeanum;但在Spooner等[11]的分类体系中,S.commersonii ssp.malmeanum不再是S.commersonii的一个亚种,而是作为一个独立的野生种,命名为S.malmeanum。
S.commersonii种内具有丰富的抗性资源,据不完全统计,S.commersonii种内不仅存在有马铃薯疮痂病、PVX、科罗拉多甲虫等病虫害的抗性,对某些不良环境也具有极强的耐性,其中最为典型的就是耐低温霜冻[11]。研究表明S.commersonii具有极强的低温驯化能力,驯化后其LT50达-10℃以下,低温驯化能力高于7℃以上[7],是已知低温驯化能力最强的马铃薯野生种。
3.2 S.commersonii在抗寒育种中的应用
目前在利用S.commersonii进行抗寒育种方面,已经取得了一些初步的成果。Caidi等[36]利用二倍体的S.commersonii和四倍体的栽培种进行体细胞融合后得到了雄性不育杂种后代,70%是四倍体,其中编号为cmm1的种间杂种的驯化能力达到了6℃;Nyman和Waara[37]利用二倍体栽培种和S.commersonii体细胞融合得到杂种,经检测,某些种间杂种驯化后能够忍受-9℃左右的低温。Chen等[38]同样利用S.commersonii和二倍体栽培种体细胞融合后获得了一个四倍体的杂种后代,该四倍体和栽培种‘Wis AG 231’回交后代中,有21%的后代低温驯化能力得到了显著提升(即驯化能力大于或等于2.5℃),其中某些植株的结薯产量甚至超过了回交亲本。Bastia等[39]同样利用体细胞融合的方法获得了不同倍性的杂种,在四倍体杂种中,一些杂种驯化后LT50低于-5℃,而在八倍体中,有些杂种驯化后LT50甚至可达-9℃。Seppänen等[40]也是通过原生质体融合的手段获得了S.commersonii和栽培种可育的体细胞杂种,其中一个编号为SH9A的杂种经过低温驯化其LT50为-5.2℃,且其自交后代中同样能检测抗低温植株存在,值得一提的,SH9A还具有对PVX的极端抗性。上述这些都是通过体细胞融合的方式得到杂种后代,而Carputo等[41,42]则是通过杂交的手段,成功获得了S.commersonii和四倍体栽培种‘UP88-P5’的F1代(三倍体),这些后代经过驯化后能够忍耐-6.2℃的低温2 h,其中编号为B10的植株可以忍耐-7.5℃的低温,在该三倍体与四倍体栽培种‘Tollocan’的BC1和BC2中,均有抗寒能力较强的植株存在(BC1中有植株的驯化抗寒性为-5.5℃,BC2中植株的驯化抗寒性也可达-4.8℃)。
4 马铃薯抗寒育种展望
马铃薯野生种拥有丰富的抗寒种质资源,但目前对抗性资源的利用尚未获得理想的效果,至今还没有能够被推广种植的抗寒品种。目前马铃薯育种技术主要有传统杂种育种、倍性育种、体细胞融合、桥梁育种、分子标记辅助选择育种以及转基因技术等方法。传统杂交育种是目前最主要的育种手段之一,但在抗寒育种上收效甚微,一方面,由于EBN不同的原因,大部分马铃薯抗寒资源难以直接和栽培种进行杂交;另一方面,大部分野生种抗寒能力是一个由多基因控制的数量性状,在多次回交的过程中容易导致抗性衰减。倍性育种、体细胞融合、桥梁育种虽然能够克服远缘杂交不亲和性,创制新的抗寒种质资源,但在后续的过程仍需要进行常规的杂交。在转基因技术上,由于目前对于抗寒能力的遗传规律和分子机制的解析尚不够完全,所取得的成果也比较少,民众对转基因技术也还存在较大的争议。此外,在分子标记开发上,目前极少有能应用于抗寒育种的分子标记被报道。因此,加强对马铃薯抗寒遗传规律和分子机制的研究,开发更多能应用于抗寒育种的分子标记,将极大的推动马铃薯抗寒育种的进程。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12 07:09:08
作物学报(2022年2期)2022-11-06 12:11:02
中国饲料(2022年5期)2022-04-26 13:42:38
中国烟草学报(2021年3期)2021-08-04 03:50:42
江苏农业科学(2017年15期)2018-02-06 07:17:42
现代园艺(2017年13期)2018-01-19 02:28:17
上海农业学报(2017年3期)2017-04-10 12:39:10
中成药(2016年4期)2016-05-17 06:07:54
中国果业信息(2015年6期)2015-01-23 17:38:07
植物营养与肥料学报(2012年1期)2012-10-26 02:49:46
