
生物有机肥对核桃园土壤细菌群落结构的影响
2019-02-27 07:24张凯煜王小娟
干旱地区农业研究 2019年6期
张凯煜,谷 洁,王小娟,高 华
( 西北农林科技大学资源环境学院,陕西 杨凌 712100 )
土壤微生物是土壤生态系统中的重要组成部分,对土壤物质转化和养分循环,维持土壤肥力和可持续利用至关重要,在农业土壤生态系统中的作用越来越受到关注[1-2]。核桃在黄龙县栽培历史悠久[3]。近年来,农民粗放式的施肥管理模式存在盲目性,长期大量施用化肥也会影响土壤微生物活性,进而降低作物的产量和品质,并造成一定的环境问题[4-5]。
生物有机肥是将外源功能菌接种到堆肥产品,即以畜禽粪便、农作物秸秆等为原料,经腐熟的有机肥产品。具有生物肥料和有机肥效应,有利于增产增收,还能培肥土壤,减少化肥的用量,同时实现农业废弃物资源化利用[6-7]。张婷婷[8]研究了不同施肥对核桃生长和品质的影响,提出施用生物菌肥和骨粉可有效提高核桃园土壤中有机质含量。杨建荣等[9]发现,生物有机肥的应用能显著提高核桃产量,改善土壤保水性和供肥能力。研究表明,施用生物有机肥能调节土壤中微生物组成[10],提高作物抗病性[11-12],使土壤健康发展。杜滢鑫等[13]利用PCR-DGGE和Biolog发现不同植物根际土壤微生物组成不同。Araújo等[14]通过T-RFLP方法发现,土地退化降低了土壤细菌的多样性,影响了细菌群落结构组成。由于高通量测序技术具有测序深度深和获得数据量大等优点,可揭示复杂微生物群落的多样性,加快了环境中不可培养和痕量微生物的研究[15],已成为研究微生物群落变化的主要方法之一。朱金山等[16]利用高通量测序技术对不同沼灌年限稻田土壤的微生物群落结构进行了分析,从门、纲、目、科、属5个水平上全面展示了稻田土壤微生物群落结构的分布。在化肥和生物有机肥处理下,土壤细菌群落的响应差异,不同处理土壤微生物区系的差异如何?哪些关键因素能够影响微生物进而调控土壤养分循环,还需要进一步研究。
本研究在试验区设置了不同施肥处理试验,采用高通量测序法比较不同施肥种类对土壤细菌群落结构和多样性的影响,并结合土壤性状揭示细菌群落变化的驱动因子,旨在从土壤微生物生态的角度丰富生物有机肥的促生理论,阐明施肥管理措施影响土壤肥力的生物学机制,为指导核桃合理施肥提供理论支撑。
1 材料与方法
1.1 试验设计
试验于2016年9月在陕西省延安市黄龙县三岔乡进行(109°83′ N,35°58E),供试品种为香铃。供试的生物有机肥由西北农林科技大学自制(添加的主要功能菌为解磷菌和解钾菌,有效活菌数达2.8×108cfu·g-1),有机质含量28.3%、氮(N) 3.8%、磷(P2O5) 2.6%、钾(K2O) 3.1%。试验设置3个处理,CK(不施肥),TN(常规施肥,施肥量为复合肥1 350 kg·hm-2,含N 15%、P2O515%、K2O 15%),TB(施生物有机肥,施肥量为5 325 kg·hm-2,以化肥养分氮含量折算即N 202.5 kg·hm-2、P2O5138 kg·hm-2、K2O 165 kg·hm-2)。一年两次施肥,即基肥、追肥,施肥量各占50%。
1.2 土壤样品采集
于2017年9月沿种植行,树干周围50 cm的根系密集分布区,使用“S形”五点法采集土壤样品。采样时去掉易受水分及各种环境因素影响的表层干土后,取5~20 cm土壤样品,用无菌镊子去除残留根系后装入无菌自封袋,用冰盒迅速带回实验室。取1份样品经冷冻干燥后,用于微生物群落结构分析,另1份风干过筛,用来测定理化指标。
1.3 测定项目与方法
1.3.1 土壤DNA提取 土壤样品DNA使用FastDNA SPIN Kit for Soil(MP Biomedicals,美国)进行提取。利用Nanodrop Spectrophotometer ND-1000 (Thermo Fisher Scientific,美国) 检测DNA浓度和纯度,确保样品浓度和纯度合格后,保存于-80℃冰箱。
1.3.2 高通量测序 将土壤DNA样品寄送至北京诺禾致源科技股份有限公司,进行16S rRNA基因V3-V4可变区高通量测序。通过UPARSE将序列在97%相似水平下进行聚类,获得可操作分类单元(OTUs)。利用QIIME进行抽平、RDP分类,得到样品OTUs详细的注释结果,进行后续分析。
1.3.3 土壤理化性质的测定 测定方法参考土壤农化分析[17]。
1.4 数据处理与分析
数据采用SPSSv 19.0进行统计分析和方差分析(LSD,P<0.05),用Microsoft Excel 2013进行绘图。利用CANOCO 4.5软件进行冗余分析,R软件绘制热度图。
2 结果与分析
2.1 土壤化学性质
从表1可以看出,与CK处理相比,TB处理中有机质、全氮、全磷分别增加了46.3%、30.6%和50.8%。生物有机肥的施用显著提高了核桃园土壤中全磷、全氮和有机质含量(P<0.05)。全钾含量在不同处理间无显著差异,在14.3~14.6 g·kg-1波动。各处理速效养分含量大小顺序为:TB>TN>CK,其中TB处理速效磷和速效钾含量分别达CK处理1.76倍和1.78倍。
2.2 土壤细菌群落多样性
高通量测序结果显示每个样品OTUs稀释性曲线均趋于平坦(图1),说明测序数据量和测序深度合理,可包括土壤样品中的大部分的细菌类型。分析了代表微生物群落α-多样性的辛普森指数、Chao1、香浓指数和ACE指数,其值越高表示微生物多样性、物种丰富度越高[18]。由图2可见,TB处理α-多样性指数均高于CK和TN处理,而TN处理香浓指数和ACE指数有所降低。
2.3 土壤细菌群落结构
基于门水平,分析了不同土壤样本序列(图3),共得到35个门水平类群。结果显示优势类群包括变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、酸杆菌门(Acidobacteria)、拟杆菌门(Bacteroidetes),芽单胞菌门(Gemmatimonadetes),共占细菌总量的85.2%~90.6%。进一步比对分析,共得到291个属水平类群,其中鞘脂单细胞菌属(Sphingomonas,phylum-Proteobacteria, 6.6%~10.0%)节杆菌属(Arthrobacter,phylum-Actinobacteria, 2.3%~4.7%)为优势类群(图4),TB处理较CK处理分别增加了17.6%和56.6%,较TN处理分别增加了51.5%和104.3%。
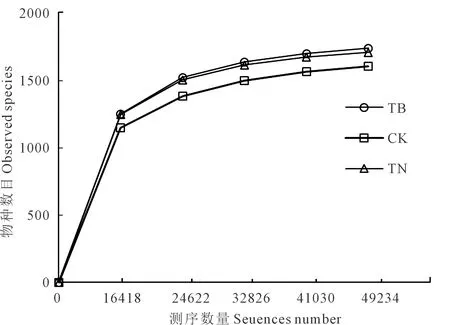
图1 土壤样本稀释曲线Fig.1 Rarefaction curves of soil samples
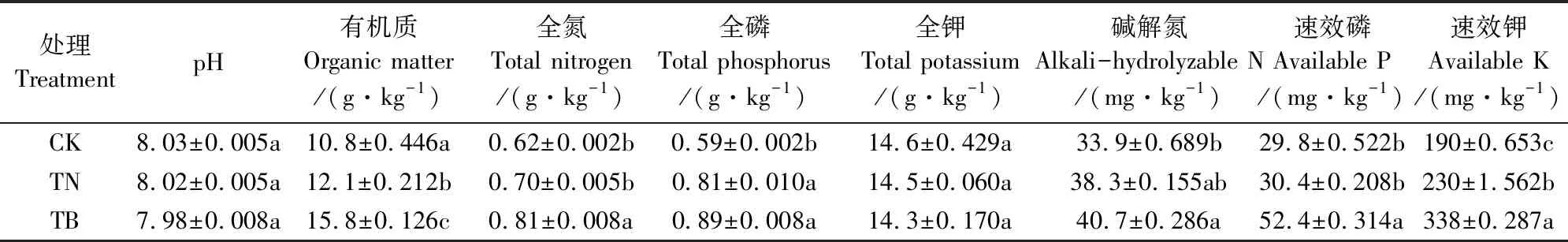
表1 不同施肥处理下的土壤化学性质
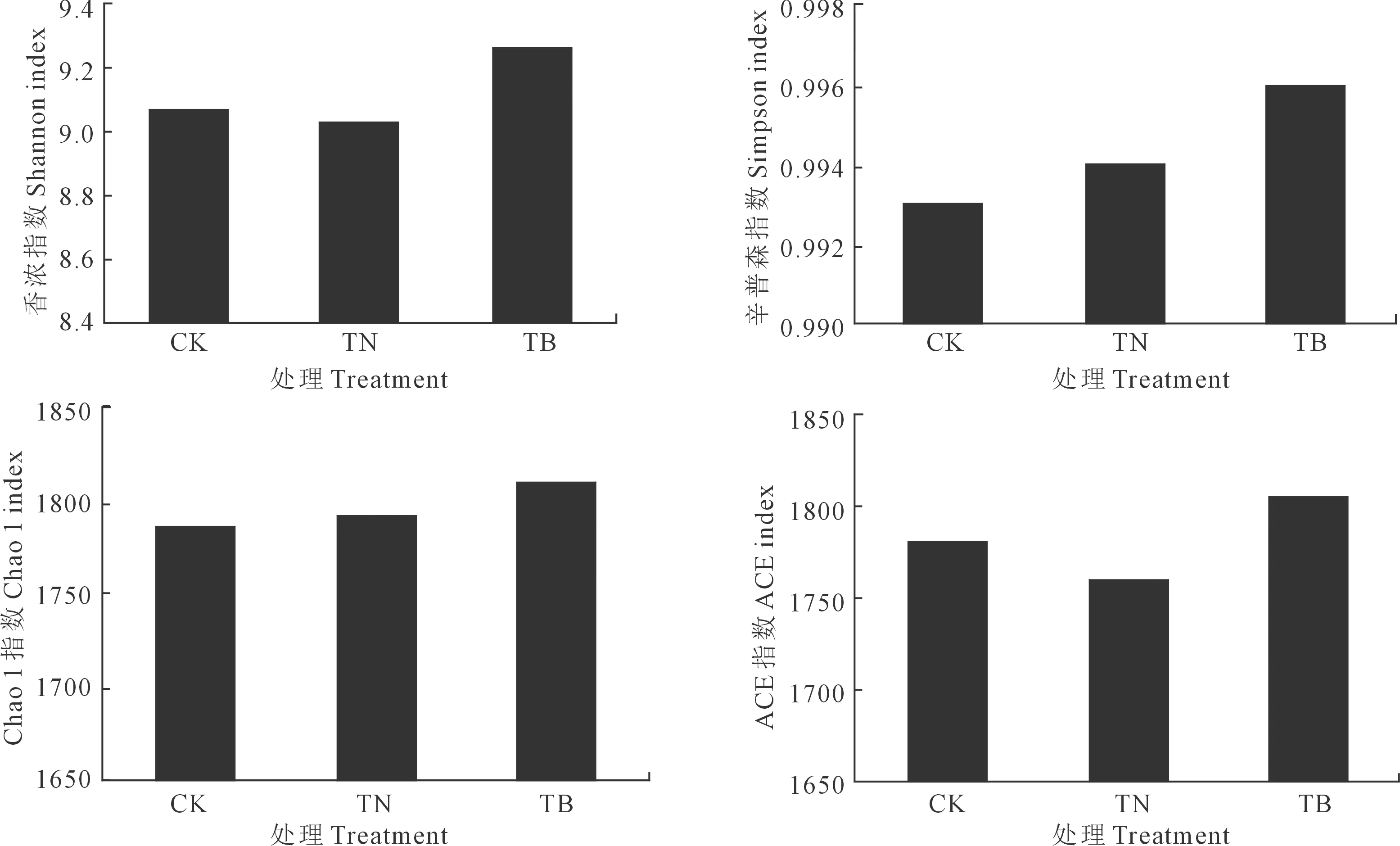
图2 不同施肥处理下的土壤细菌群落α-多样性指数Fig.2 Abundances of bacterial community α-diversity indexes under different fertilizer treatment
以门水平上细菌相对丰度和与土壤pH、有机质、碱解氮、全氮、速效磷、全磷、速效钾、全钾进行冗余分析(图5),可分析细菌群落和多个响应变量(土壤理化性质)之间的关系[19]。结果显示,所选取的环境因子共解释细菌群落变化的94.6%,RD1轴解释了82.5%,RD2轴解释了12.1%。其中土壤有机质、碱解氮和速效钾含量为不同施肥处理间细菌群落差异的主要驱动因子。
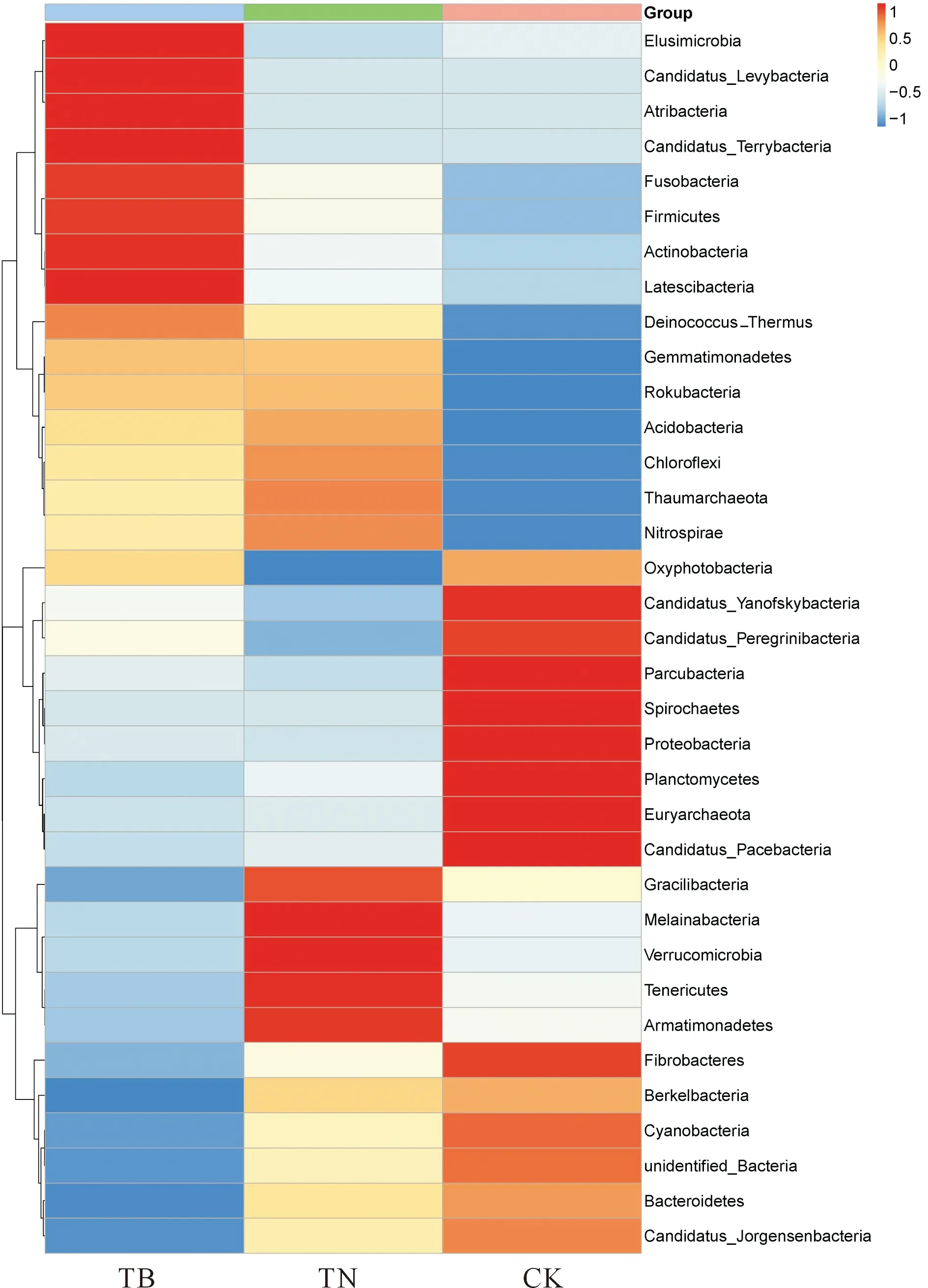
图3 基于门水平的不同施肥处理下土壤细菌群落组成Fig.3 Soil bacterial community composition in different fertilization treatments at phylum level
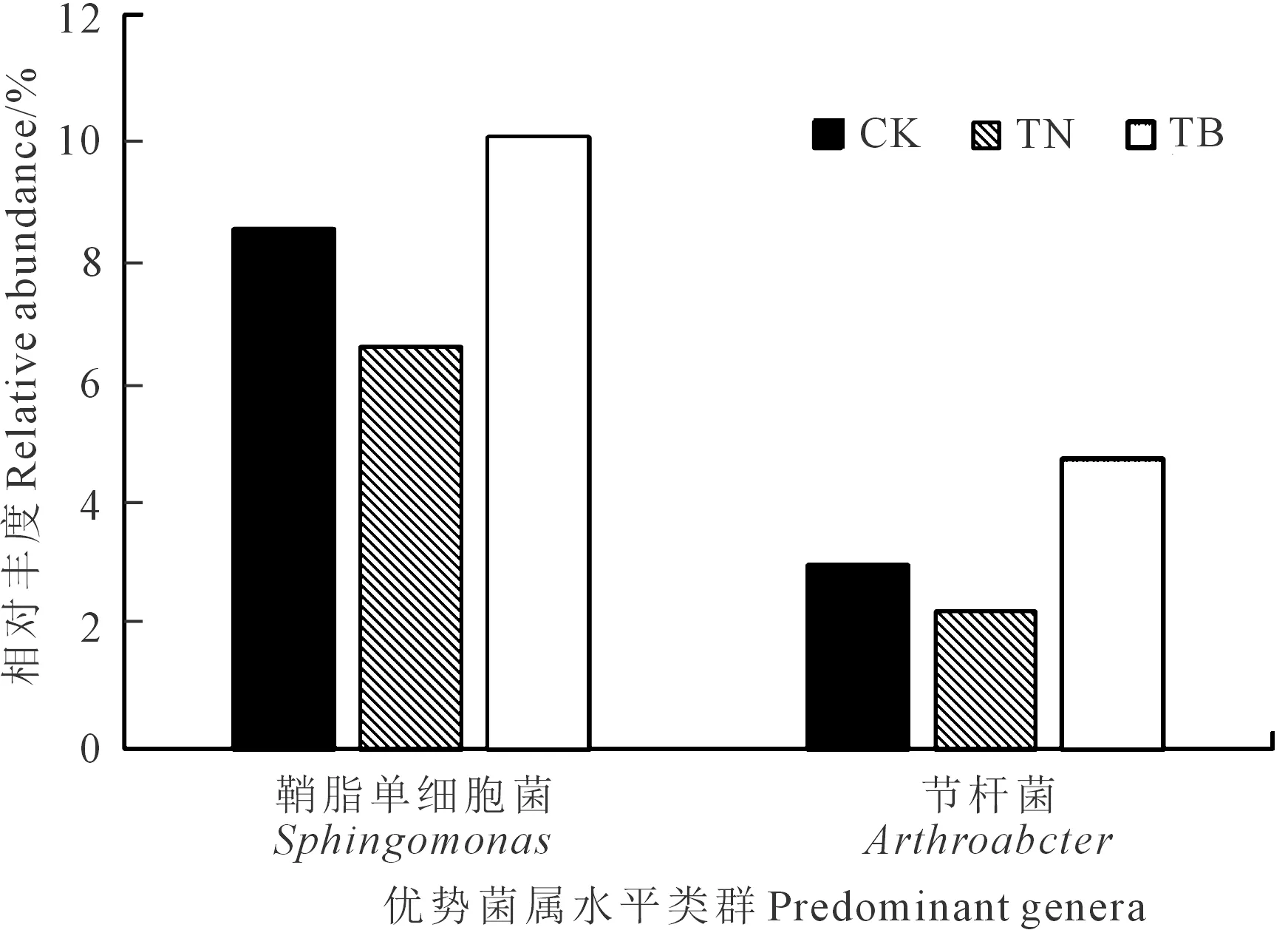
图4 不同施肥处理下的土壤细菌群落优势菌属Fig.4 Predominant genera of soil bacterial community in different fertilization treatments
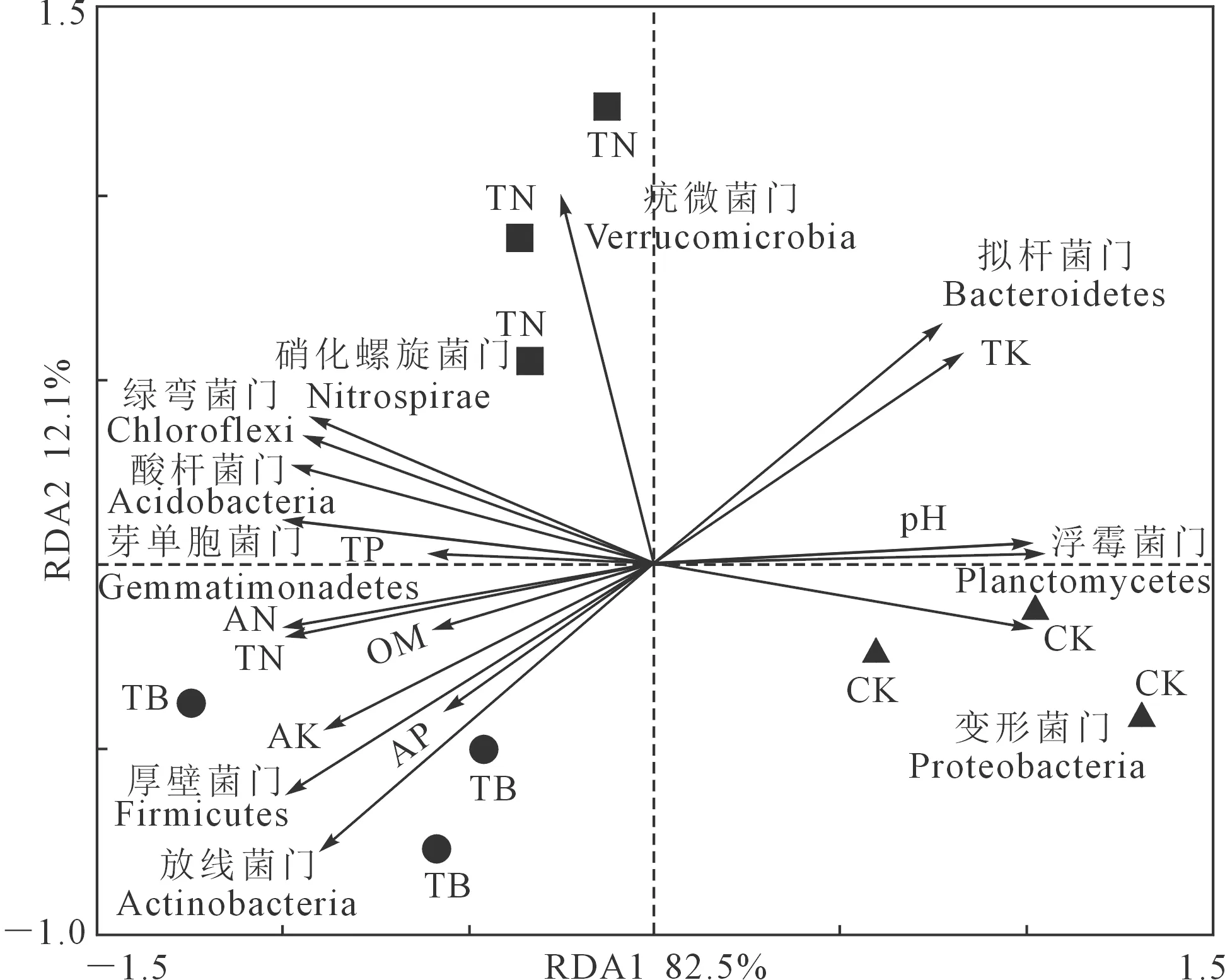
注:TP:全磷;AN:碱解氮;OM:有机质;AK:速效钾;AP:速效磷;TK:全钾;TN:全氮。Note: TP: Total phosphorus; AN: Alkali-hydrolyzable N; OM: Organic matter; AK: Available K; AP: Available P; TK: Total potassiam; TN: Total nitrogen.图5 不同施肥处理下的环境因子对细菌群落结构的RDA排序图Fig.5 Redundancy analysis between soil properties and the bacterial community structure
3 讨 论
生态系统稳定性及土壤肥力发生变化的关键因素是人为向土壤中大量输入外源物质,尤其是肥料[20-21],不同的施肥方式和肥料类型影响又有所不同。生物有机肥的施用能够提高土壤有机质和全氮含量,这与前人研究结果相似[22]。常规施肥对土壤速效养分的提高效果不如生物有机肥,尤其是土壤中速效钾和速效磷含量。这可能主要是因为生物有机肥中含有解磷菌、解钾菌可作用于土壤中难溶的磷和钾,释放出大量速效磷、速效钾。生物有机肥既能在作物生长前期快速提供一定的速效养分,又可在作物生长过程中通过微生物分解有机质和矿物质释放养分,发挥速效和长效兼有的作用,养分的缓慢释放还可为微生物提供更稳定的栖息环境,起到“接种”和“导入”作用[23- 24]。
土壤细菌在土壤养分循环和系统维稳中起重要作用[25]。本研究结果表明,施用生物有机肥显著增加了土壤细菌群落多样性,可能是因为施肥处理中较高的有机质和养分,为更多的微生物提供了生长和繁殖的条件,其中包括生物有机肥中本来携带的大量功能菌[26]。与不施肥相比,常规施肥却降低了细菌群落香浓指数和ACE指数,这与苏婷婷等[11]的研究结果相似。Liu等[27]通过Biolog方法发现施用有机肥处理香浓指数显著高于化肥处理,Chen等[28]发现接种微生物菌剂能够显著提高土壤丰富度指数。综上表明,生物有机肥优于常规肥料的关键因素就在于这些功能菌的生命活动。
通过对不同施肥处理对细菌群落结构的分析,得到35个门水平和291个属水平的类群。结果表明,不同处理中优势的门、纲和目水平上优势物种相似,但不同处理处理间,门、纲和目水平物种相对丰度不尽相同。变形菌门、放线菌门、酸杆菌门、拟杆菌门、芽单胞菌门为优势门类,这与某些前期研究结果相似[16, 29]。而不同研究中物种丰度差异较大,说明农田土壤中细菌群落优势物种种类相似,但其相对丰度会受土壤类型、种植作物类型以及施肥灌溉方式等的影响[30-31]。放线菌门在土壤氮素循环中起重要作用,还能促进土壤中植物残体的腐烂。本研究中,各处理放线菌门丰度大小为TB>TN>CK,与全氮和碱解氮趋势一致,这与王伏伟等[32]研究相似。他研究了施肥和秸秆还田对砂浆黑土细菌群落的影响,发现施肥和秸秆还田显著提高了放线菌门的相对丰度,并且土壤全氮含量是提高其相对丰度的重要原因。而杨亚东等[29]研究结果表明在OTUs水平上细菌群落结构与全氮含量显著相关,但门水平上则相关性不显著。这表明全氮含量对细菌不同分类学水平上的群落结构影响存在差异。在属水平上,不同处理中,鞘脂单细胞菌属和节杆菌属菌为优势物种,但在不同处理中相对丰度不同(TB>CK>TN)。鞘脂单细胞菌属可以促进根际营养吸收,抵抗多种病原菌,是降解土壤有毒物质最有效的微生物菌属之一。也有研究发现,鞘脂单细胞菌属中部分菌株具有固氮特性,在土壤氮素转化方面起重要作用[33]。节杆菌属可以通过固氮作用产生多种植物激素,以促进水分和矿质元素的吸收。由此可见,施用生物有机肥可以促进优势菌的富集,促进营养和水分等吸收,其是否能提高根际微生物抗病抗毒害能力,还需要今后长期试验来证实。
土壤微生物群落组成会受到环境变化的影响,可以作为土壤质量变化的指标。武发思[34]研究发现有机肥对根际土壤细菌多样性的增加和群落结构的改变作用明显。本文采用冗余分析(RDA)来研究环境因子对微生物群落的影响,将RDA分析加入高通量生物信息分析,能够更直观地表达影响研究区土壤细菌群落特征的主要土壤环境因素,是分析土壤微生物信息的有效方法,能够为土壤环境评价以及土地利用管理提供科学依据。RDA结果表明,施肥处理即TB和TN处理在RD1轴上与CK处理显著区分,有机质、碱解氮和速效钾是土壤细菌群落结构改变的主要驱动因子。有机质改变了土壤孔隙度、通气度与土壤团粒结构[35],具有一定的缓冲作用,作为土壤酶的重要载体,为土壤微生态环境提供化学反应场所,为细菌活动提供适宜条件。而碱解氮和速效钾对细菌群落结构影响较大,可能和核桃本身养分需求相关。
4 结 论
施生物有机肥可显著提高核桃园土壤养分,使土壤理化性质得到改善,且生物有机肥效果优于常规施肥。施生物有机肥增加了土壤细菌群落多样性,在一定程度上还提高了土壤中优势菌属的相对丰度。不同施肥处理改变了细菌群落结构和组成,冗余分析结果表明,土壤有机质、碱解氮和速效钾含量是影响细菌群落变化的主要驱动因子。本研究采用了高通量测序的方法,结合环境因子对土壤细菌群落对不同施肥处理的响应进行了分析,后续可以利用宏基因组和宏转录组的方法,对参与碳、氮、磷循环功能微生物的表达、代谢途径进行深入研究。
猜你喜欢
中国典型病例大全(2022年11期)2022-05-13
养生阅刊(2021年5期)2021-06-03
透析与人工器官(2020年1期)2020-11-16
中华养生保健(2020年7期)2020-11-16
中国现代中药(2019年5期)2019-07-03
农业与技术(2017年12期)2017-07-11
中老年健康(2016年12期)2017-01-18
绿色科技(2016年16期)2016-10-11
江苏农业科学(2016年6期)2016-07-25
江苏农业科学(2015年11期)2016-01-27
