
黄土高原丘陵区枣和酸枣叶脉序特征分析
2019-02-27 07:24:10李晓鹏李新岗
干旱地区农业研究 2019年6期
李晓鹏 ,李新岗
(1.兰州财经大学,甘肃 兰州 730020;2.西北农林科技大学林学院,陕西 杨凌 712100)
叶脉系统不仅是植物水分和养分运输的重要器官[1],而且对叶片具有机械支撑作用,以保证叶片的生理活动[2]。叶脉序表征了叶脉在叶片里的分布和排列方式[3],被称为植物的“血管”。不仅叶脉序特征具有一定的分类学意义[4-5],而且叶水分利用、光合能力、碳构建等也备受关注[2, 6-7]。叶脉系统具有等级特征,末端叶脉的供给和运输作用最为关键[2]。Roth-Nebelsick等[8]通过研究叶脉形态和功能的关系以及叶脉的演化历程,认为不同植物类群具有不同类型的叶脉序。不同植物类型的叶脉密度、叶脉宽度和叶脉间距离等性状共同决定了其叶片水分供给和运输能力,也反映了叶脉系统对养分和光合产物的运输能力[9-10]。随着叶脉序研究的不断深入,其应用领域也在不断拓展,已成为研究叶功能多样性、叶片碳平衡的基础领域[11]。有关叶脉序特征与环境[3, 12-13]、与植物系统进化[14-15]的研究已取得了一些成果,对植物生理学和生态学的研究及相关预测都具有重要的意义[2]。
关于枣和酸枣适应性方面的研究,主要集中在胁迫条件下,对枣叶片表皮、含水量和叶绿素含量等方面[16-18]。在栽培(枣)和野生(酸枣)生境条件下,研究枣和酸枣叶脉序特征,探讨其叶脉序适应机制具有重要意义。
枣(ZiziphusjujubaMill.)和酸枣(Z.jujubavar.spinosa)为枣属植物,枣是由酸枣演化而来[19]。晋陕黄河峡谷一带作为我国枣的原产地和栽培中心,一直保留着枣的原生类型和很多过渡型酸枣。本研究以晋陕黄河峡谷的枣、酸枣和过渡型为材料,研究枣和酸枣叶脉序特征,揭示其叶脉序特征及其叶片结构建成,探讨其适应机制,有助于深化对枣和酸枣在原始生境中生产力、抗性机制、生态适应能力和生存机制的理解,为枣资源的高效利用提供理论依据。
1 材料与方法
1.1 材料的收集与准备
采样区位于陕晋黄土高原丘陵(黄河峡谷)枣区,主要采样地为陕西佳县、清涧县、延川县、吴堡县,以及山西稷山县和中阳县(图1)。该区属暖温带半干旱大陆性气候,海拔480~1 300 m,年降雨量为380~400 mm,降雨主要集中在秋季,年蒸发量为1 400~2 000 mm。枣区多在河流沿岸的滩地、或者海拔较低的河谷和丘陵区。
枣和酸枣在形态上各有特点,枣为乔木,托刺易脱落,叶缘疏且较大,果核少仁或无仁,果实可食率94%~97%;酸枣为灌木、小乔木、乔木,托刺多、长,不容易脱落,叶缘锯齿细密,果核饱满、多仁,果实可食率仅64%~87%。酸枣与枣间存在很多过渡类型,多为乔木,托刺少、短、容易脱落,叶缘、果核、果实可食率等性状介于酸枣和枣之间。
7月中、下旬到9月中旬,待叶片发育完全后,采集90份代表种质,其中包括枣55份、酸枣28份和过渡型7份。枣品种有狗头枣、狗稀疙瘩枣、狗牙枣、灵宝圆枣、木枣、稷山圆枣、牛奶脆枣、帅枣、团枣、板枣、锥枣、蜂蜜罐、伏脆蜜、江淮1号、晋枣、冷白玉、七月鲜、月光枣和猪腰枣等。利用GPS定位,记录各枣品种和酸枣所处的地理位置。
采样时,针对每个枣品种或酸枣类型,随机选取树龄30 a以上的健康植株10 株,每株上采摘3~5 a生枣股上的枣吊10~20个,标记后放入自封袋中,带回实验室后在 -4℃ 冰箱内保存。
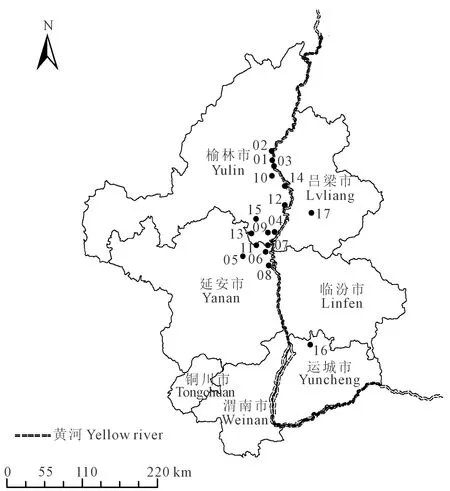
注:1-15为陕西样品:佳县样品(1.佳芦镇;2.朱家坬镇;3.峪口镇;10.店镇;14.蟋镇);清涧县样品(4.高杰村镇;9.老舍古乡;13.宽州镇;15.石咀驿镇);延川县样品(5.禹居镇;6.延水关镇;7.眼岔寺乡;8.土岗乡;11.贺家湾乡);吴堡县样品(12.守家川镇);16-17为山西样品:16.稷山县稷峰镇;17.中阳县下枣林乡。Note: 1-15 represents Shaanxi sample: Samples in Jiaxian County(1. Jialu town; 2. Zhujiagua town; 3. Yukou town; 10. Dian town; 14. Xi town); Samples in Qingjian County(4. Gaojiecun town; 9. Laoshegu town; 13. Kuanzhou town; 15. Shizuiyi town); Samples in Yanchuan County(5. Yuju town; 6. Yanshuiguan town; 7. Yanchasi town; 8. Tugang town; 11. Hejiawan town); Samples in Wubao County(12. Shoujiachuan town); 16-17 represents Shanxi sample: 16. Jifeng town in Jishan County; 17. Xiazaolin town in Zhongyang County.图1 陕晋黄土高原丘陵枣区(黄河峡谷)17个采样地分布情况Fig.1 Distribution of 17 sampling sites of jujube leaves on Loess Plateau of Shaanxi and Shanxi (The Yellow River Canyon)
1.2 叶脉序信息获取
定位3~5 a生二次枝上的中部枣股,选取其枣吊中部3~5个叶片,采用背光扫描法采集叶脉信息,并采用叶脉序信息转换软件LEAF GUI(Leaf Extraction and Analysis Framework Graphical User Interface)[20]进行信息转化,获取叶脉数据信息。选取叶面积、叶周长、网眼数、环形结构、叶脉宽度、网眼面积、节点数、叶脉间距离、叶脉密度、叶脉总长度、叶脉总面积、叶脉边缘的平均二维面积、叶脉边缘的平均三维面积和闭合环状结构平均体积等14个特征指标[21]。其中,叶脉密度指单位面积叶脉的总长度;环形结构即单位面积网眼数,指叶片内闭合环形结构的数量除以叶面积;节点数指叶片中一、二、三级脉等交汇点的总个数;叶脉间距离指叶脉所围成的每个闭合区域的最大当量直径;网眼面积是叶片的一、二、三级脉等围成的不规则闭合区域的平均面积。
1.3 数据处理
用ArcGIS 10.2软件制作采样图;SPSS 19.0软件对各叶脉序特征指标数据进行方差分析(ANOVA)、多重比较检验(post-hoc tests)和多样性分析(SPSS_Inc. 2004);用NTSYSpc V2.1和MEGA 5.1 聚类分析软件对其叶脉序进行聚类分析[22];并使用 Sigmaplot 11软件(Systat Software, Richmond, CA, USA)和R 3.5.1软件做图。
2 结果与分析
2.1 枣、酸枣和过渡型叶脉序指标比较
陕北枣、酸枣和过渡型酸枣的14个叶脉序特征,通过主成分分析(PCA)筛选出适合研究枣叶脉序的8个特征指标,即叶面积、叶周长、环形结构、叶脉间距离、网眼面积、节点数、叶脉密度和叶脉宽度。8个叶脉序指标方差分析(SPSS_Inc. 2004)(图2)表明,就叶面积和叶周长而言,枣、酸枣和过渡型酸枣之间均有显著差异(P<0.05),且枣最高、酸枣最低。枣、酸枣和过渡型的叶脉密度也有显著差异,且枣 < 过渡型 < 酸枣(P<0.05)。对叶脉序环形结构、叶脉间距离、网眼面积和节点数来说,枣与酸枣均有显著差异(P<0.05),但枣和过渡型之间无差异。同时,枣、酸枣和过渡型的叶脉宽度无显著差异(P>0.05)。
多样性分析表明,枣叶脉序指标的多样性指数(DI),单位面积网眼数(1.53)、叶脉间距离(1.64)、网眼面积(1.65)、节点数(2.03)、叶脉密度(1.74)均显著高于酸枣(P<0.05),过渡型酸枣叶脉序指标的多样性指数处于枣和酸枣之间。酸枣叶脉序指标中,环形结构的多样性指数最高,网眼面积的多样性指数最低;枣叶脉序指标中,节点数的多样性指数最高,叶脉宽度的多样性指数最低;过渡型酸枣的叶脉序指标中,节点数的多样性指数最高,叶脉间距离的多样性指数最低(图2)。枣叶脉序特征指标的变异系数(CV)范围(15.24%~74.69%)比酸枣(20.37%~52.84%)更宽。其中,枣环形结构的变异系数最大(74.69%),枣叶脉间距离的变异系数最小(15.24%)。酸枣中,环形结构的变异系数最大(52.84%),叶脉间距离的的变异系数最小(20.37%)。枣、酸枣和过渡型环形结构的变异系数均较高,叶脉间距离的变异系数均较低。
2.2 枣、酸枣和过渡型叶脉序聚类分析
2.2.1 陕晋黄土高原丘陵(黄河峡谷)枣区枣、酸枣和过渡型的叶脉序聚类分析 由于延川县存在着较多典型的古酸枣树和过渡型酸枣,具有一定的代表性。故首先选取该地区代表性枣、酸枣和过渡型进行叶脉序NJ聚类(Neighbor-joining)分析,结果发现,在相似系数为1.8 时,将其分为两大类,第I类为酸枣,第II类为枣(图3)。

图2 枣、酸枣和过渡型酸枣的叶脉序特征及多样性指数比较Fig.2 Comparisons of eight leaf venation characteristics and diversity index for cultivated jujube, wild jujube, and transitional type

图3 陕北酸枣、过渡型酸枣和枣的NJ聚类分析Fig.3 Dengrogram of neighbor-joining clustering analysis of wild, transitional type and cultivated jujube accessions in Northern Shaanxi.
陕晋黄土高原丘陵枣区的枣、酸枣和过渡型的叶脉序 NJ 聚类分析(图4)表明,品种间遗传多样性高,杂合度高。在相似系数为1.69时,将其分为两大类,第I类为酸枣,第II类为枣;并且,过渡型酸枣的佳县酸团枣、苦味酸枣、老牙酸枣和酸味酸枣,在两个类群中都有,说明酸枣经过渡型向枣的演化。同时,在酸枣类群中,清涧酸枣和延川酸枣之间距离较近,而佳县酸枣与清涧酸枣距离较远。另外,在枣中,佳县木枣、清涧木枣和延川木枣的距离较近,聚为一类,为木枣类群。而延川团枣、清涧团枣与木枣中的部分佳县、清涧木枣的距离较近。狗头枣、中阳木枣、伏脆蜜、佳县白枣、晋枣的距离较近,七月鲜、狗牙枣与牛奶脆枣的距离较近。
2.2.2 陕晋黄土高原丘陵(黄河峡谷)枣区主栽枣品种的叶脉序聚类分析 为了进一步研究陕晋黄土高原丘陵(黄河峡谷)枣区主栽枣品种叶脉序之间的关系,19个枣品种UPGMA聚类分析(图5)表明,聚类中存在不同的层次(L1-L3),在相似系数为1.29时,枣被分为4个类群。
类群1包括白枣、猪腰枣、伏脆蜜、江淮1号、延川狗头枣、狗稀疙瘩枣、团枣、冷白玉、七月鲜和月光枣等11个品种;类群2包括晋枣、牛奶脆枣、锥枣和帅枣4个品种,主要为鲜食和制干的兼用品种;类群3有脆脆枣、狗牙枣和灵宝圆枣3个品种;类群4只有木枣。从叶脉序特征的角度分析,类群3中的脆脆枣、狗牙枣、灵宝圆枣和类群4中的木枣之间的距离较近。
3 讨 论
3.1 枣、酸枣和过渡型叶脉序特征比较
植物在进化过程中,为适应环境变化自身会启动相应的基因改变或调整生理生化代谢过程,表现出相应的生态适应性[23]。植物叶片形态结构与其生理功能和环境关系密切,其对环境的变化最敏感,可塑性最大[24]。
枣和酸枣叶脉序特征指标方差分析表明,酸枣叶脉间距离显著低于枣(图2),说明其叶脉与气孔间的距离较短。当终端叶脉之间的距离越小、叶脉直径越大时,叶脉运输水分的能力越强[11],碳通量越高[25]。因此,与栽培条件下的枣相比,野生条件下的酸枣具有较高的碳通量和水分运输能力,并且其环形结构显著高于枣和过渡型。有研究表明,当植物受到损伤时,叶脉的环形结构能通过对叶片水分运输途径的重新布局,把损伤减少到最小[3, 26],环形结构的不断演化也与气孔的分布及密度相关[8]。与枣相比,酸枣的生存环境更为恶劣,正是由于酸枣较高的环形结构,使其在遭受机械损伤后,能够通过增加更多水分运输途径的方式,弥补和减少外界伤害。酸枣叶脉密度显著高于枣和过渡型(图2)。对较高叶脉密度植物的研究发现,减少土壤有效水分和植物所需养分均可提高植物的叶脉密度,而植物可通过较高的碳投入量来补偿土壤中水分和营养的缺失[11]。酸枣在干旱环境中,具有比枣更高的叶脉密度,说明其单位面积有更多的水分、养分运输通道和碳投入量,发达的输导组织能保证和提高水分输导效率,从而保证叶肉组织不因失水受到伤害。以上结果均从叶脉序的角度上解释了酸枣能在极度寒冷、干旱的环境中正常生长的原因。
叶脉序聚类分析表明,第I类为酸枣,第II类为枣(图3和4),说明叶脉序可以作为一个重要的指标来划分枣和酸枣。同时,过渡型酸枣在两个类群中都有,这是由于过渡型酸枣的叶脉序特征代表着酸枣到枣演化过程的不同阶段,叶脉序也反映了枣是由酸枣经过渡型酸枣演化而来。
3.2 枣、酸枣和过渡型叶脉序的多样性分析
植被表型特征的变异是其不断应对不同环境的一种生存机制[11]。本研究表明,酸枣叶片形态和大部分叶脉序特征的变异系数显著高于枣(图2)。Richards[27]研究发现,表型可塑性较高的植物,其对环境条件具有较强的耐受性,因而分布范围较广;反之分布范围较小。长期进化过程中,过渡型和枣多为乔木,而酸枣有乔木和灌木两种类型。这说明酸枣在水平空间分布上有生态位重叠,是一种互利共生,也能减少或避免它们之间对生态资源的直接竞争,表现出酸枣对有限资源充分利用的一种有效生存机制。

图4 黄土高原丘陵(黄河峡谷)枣区枣、酸枣和过渡型酸枣的叶脉序特征及其聚类分析Fig.4 Leaf venation characteristics and clustering analysis for cultivated, wild, and transitional-type jujube sampled on hilly region Loess Plateau(The Yellow River Canyon)
本研究发现,枣叶脉序特征的多样性指数均显著高于酸枣(图2)。枣的栽培和驯化受人为选择和自然选择的影响,尤其是人为选择,深刻影响着枣品种选择,从而形成了适应当地环境的品种和生态类型,并在各地进行广泛的交流[28]。各异的生长环境,为枣品种多样性提供了充分的外在条件。
同时,酸枣叶脉序特征的变异系数显著高于枣和过渡型,使其具有比枣更宽的生态幅。张继义等[29]研究发现,与生态位宽度较小的植物相比,生态位宽度较大的植物对资源的有效利用能力较强,分布范围较广。与低表型可塑性植物相比,高表型可塑性的植物通常对不同环境条件具有较强的耐受性,因而分布广[30]。在长期进化过程中,酸枣面对干旱、贫瘠的生长条件,为保证生长发育顺利进行,通过表型性状(尤其是叶片)的变异适应环境变化,表现出较高的表型可塑性和较强的生态适应性。
3.3 基于叶脉序的枣品种关系
陕晋黄土高原丘陵(黄河峡谷)枣区19个枣品种UPGMA聚类结果表明,在相似系数为1.29时,枣分为4个类群(图5)。类群1中有11个品种,其中白枣、猪腰枣、伏脆蜜、冷白玉、七月鲜、月光枣、江淮1号和团枣是鲜食品种,狗稀疙瘩枣为制干品种,延川狗头枣为鲜食与制干兼用品种。类群2中,晋枣和牛奶脆枣为鲜食品种,锥枣和帅枣分别为制干和蜜枣品种。类群3中,脆脆枣和狗牙枣是鲜食品种,灵宝圆枣是制干品种。类群4只有木枣品种,为该枣区的原生品种群。该部分的划分更多是源于叶脉序特征及其功能相近的结果,与枣品种类别关系不大。但也看到,类群1中的狗头枣属关中品种群,与类群4中的木枣的距离较远,这与枣遗传多样性的研究结果一致[31]。
陕北地区保存着较为原始的枣品种类型,如木枣品种群,虽然它们在树形、果形等差异很大,但其叶脉序特征差异较小。现存的千年古树与其分化的木枣亲缘关系较近,原生木枣品种群的遗传变异很小[32]。叶脉序特征聚类结果与栽培枣遗传多样性的分布格局基本一致。从叶脉序特征的角度分析,类群3中的脆脆枣、狗牙枣、灵宝圆枣和类群4中的木枣之间的距离较近。这与其遗传演化研究结果基本一致,亲缘关系较近,可能是由于脆脆枣、狗牙枣和灵宝圆枣是从原始品种木枣中的变异株系中形成的。Geber和Griffen[33]研究发现,植物在进化过程中,植物会选择易适应当地条件的表型。
4 结 论
本研究分析了我国枣起源地——黄土高原丘陵(黄河峡谷)枣区的枣、酸枣和过渡型的叶脉序特征,发现叶脉作为稳定性状可用于酸枣和枣的鉴别。过渡型叶脉特征在栽培枣与酸枣之间,从叶形态上证实了酸枣到栽培枣的进化。在干旱、贫瘠生境中生长的酸枣,其叶脉间距离、网眼面积和节点数都显著低于栽培枣,而环形结构和叶脉密度显著高于枣。说明枣和酸枣通过权衡叶脉序而采取不同的适应策略。枣叶脉序的多样性指数(DI)中,单位面积网眼数(1.53)、叶脉间距离(1.64)、网眼面积(1.65)、节点数(2.03)、叶脉密度(1.74)均显著高于酸枣。枣叶脉序特征指标的变异系数范围(15.24%~74.69%)比酸枣(20.37%~52.84%)更宽。这是由于在枣的栽培生产中,人类往往以自身需要为目的选择枣品种,因此深刻影响着品种的选择方向。酸枣叶脉序变异系数显著高于枣和过渡型,说明其具有更宽的生态幅,表现出较高的表型可塑性和较强的生态适应性,体现了对有限环境资源充分利用的生存机制。
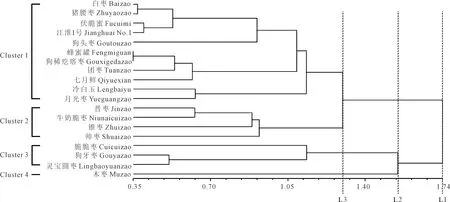
图5 陕晋黄土高原丘陵(黄河峡谷)枣区19个主栽枣品种的UPGMA聚类图Fig.5 Dengrogram based on clustering analysis of unweighted pair group method with arithmetic average for 19 varieties of cultivated jujube sampled on Loess Plateau of Shaanxi and Shanxi (The Yellow River Canyon)
猜你喜欢
今日农业(2022年13期)2022-09-15 01:18:00
动漫界·幼教365(大班)(2021年11期)2021-11-20 07:49:02
中成药(2021年5期)2021-07-21 08:38:46
发明与创新(2020年26期)2020-06-27 07:25:06
河北果树(2020年1期)2020-02-09 12:31:42
小学生学习指导(低年级)(2019年9期)2019-09-25 07:43:16
文萃报·周五版(2019年44期)2019-09-10 22:55:44
红领巾·萌芽(2017年2期)2017-03-09 15:25:16
江苏丝绸(2014年3期)2014-08-15 00:49:06
杂文选刊(2014年5期)2014-04-24 03:59:18
