
红壤侵蚀区植被恢复对土壤呼吸及其温度敏感性的影响*
2019-02-26 03:30吕茂奎张康靖谢锦升
土壤学报 2019年1期
邓 翠 吕茂奎 曾 敏 张康靖 张 浩 谢锦升†
(1 福建师范大学地理科学学院,福州 350007)
(2 湿润亚热带山地生态国家重点实验室培育基地,福州 350007)
土壤呼吸是陆地生态系统碳循环的重要过程,是土壤向大气输出碳的主要途径,每年向大气输出83~108 Pg C[1],是化石燃料排放的10倍以上。随着温度不断升高,在全球变暖的背景下,土壤呼吸的强弱很大程度上决定了全球气候变化和碳循环变化的反馈关系,其微小的变化都能显著改变大气CO2浓度。土壤呼吸主要由微生物分解和根际呼吸组成,植被变化通过改变植物数量及群落组成,改变根系生物量及凋落物数量和质量而影响着土壤呼吸,另一方面,凋落物在土壤微生物的作用下形成土壤有机质而对微生物产生反馈作用,这些过程均会对土壤呼吸造成一定程度的影响[2-5],同时,植被覆盖也可通过改变土壤温度调节土壤呼吸[3]。土壤呼吸温度敏感性(Q10)反映了土壤呼吸随土壤温度的变化关系,一般情况下,随着土壤温度持续升高,土壤呼吸不再上升,Q10下降,造成土壤呼吸不变或下降,这可能是由于呼吸底物的有效性受限、加速土壤有机氮的矿化导致C/N失衡、水分限制和微生物群落结构的改变等造成的[5-7]。
我国是全球水土流失最严重的国家之一,水土流失总面积为484.7万km2,占国土总面积的51.1%[8]。福建长汀县是我国南方花岗岩地区最典型的水土流失区,存在大面积的侵蚀裸露地,由于缺少植被覆盖导致土壤温度显著提高。从20世纪80年代起,当地政府采取许多治理措施,水土流失得到有效控制,以大面积马尾松(Pinus massoniana)种植为主的植被得到不同程度的恢复,随着地力条件的改善,林下植被—芒萁(Dicranopteris dichotoma)不断入侵,成为红壤侵蚀区主要的林下水土保持植被。
前期对严重侵蚀红壤室内培养试验表明,温度越高矿化量越高,土壤温度10~20 ℃的 Q10为1.41,然而土壤温度20~30 ℃ Q10仅为1.06[9],表明严重侵蚀地土壤由于夏季高温而可能具有较低的Q10。植被恢复后有利于调节土壤温度,但相应的土壤呼吸的动态变化过程及其温度敏感性是如何变化的仍不清楚。因此,本研究选取典型红壤侵蚀区福建长汀河田镇未治理地和治理后的马尾松人工林为研究对象,通过野外土壤环境因子和土壤呼吸的定位观测,探讨植被恢复对侵蚀地土壤呼吸的影响,为进一步阐明红壤区侵蚀土壤的有机碳积累机制提供科学依据。
1 材料与方法
1.1 研究区概况
研究区福建省长汀县河田镇地处福建西南部(116°18′~116°31′E,25°33′~25°48′N),海拔300~500 m,属中亚热带季风气候区,雨热同期,根据长汀县多年平均气温降雨量观测,雨季3—8月(图1)。年均气温17.5~18.8℃,年均降雨量1 737 mm,年均蒸发量1 403 mm,年均无霜期260 d,平均日照时数1 925 h,≥ 10℃积温为4 100~4 650℃。土壤为花岗岩发育的红壤,抗蚀能力低。地带性植被(常绿阔叶林)破坏殆尽,现有植被主要以马尾松次生林和人工林为主。

图1 月均降水量和气温分布Fig. 1 Distributions of monthly mean rainfall and air temperature
1.2 研究方法
利用时空代换法,于河田镇选择3个不同恢复年限的马尾松林地组成植被恢复序列,各个林地在治理前均为土壤A层流失殆尽,B层裸露,本底条件基本相似。各试验地的基本情况如下:(1)Y0,未治理地:来油坑,坡度15°。土壤强度侵蚀,地表浅沟发育,立地条件极差,乔木主要以稀疏的马尾松为主,平均树高2.3 m,平均地径4.5 cm,植被覆盖度15%,林下少量芒萁零星分布,芒萁地上生物量39.13 g·m-2,马尾松年凋落量为85.07 g·m-2。(2)Y13,恢复13 a:石壁下,坡度8°。从2002年开始实施植被恢复措施,具体采用穴状整地,在坡面马尾松之间采挖穴,穴面宽×深×底宽为50 cm×40 cm×30 cm,每穴施有机肥0.25 kg,2 490穴·hm-2,每穴用类芦(Neyraudia reynaudiana)苗分兜栽植或播种宽叶雀稗(Paspalum wettsteinii Hackel),治理后任其自然生长,现林下植被以芒萁为主,盖度90%,芒萁地上生物量887.3 g·m-2,马尾松平均树高9.5 m,平均胸径10.4 cm,马尾松年凋落量为392.9 g·m-2。(3)Y31,恢复31 a:水东坊,坡度11°。从1984年开始实施植被恢复措施,具体采用水平撩壕整地,种植黑荆(Acacia mearnsii),之后通过封禁管理来减少人为干扰和破坏,使其自然恢复,但黑荆因几次寒流大部分死亡,目前为马尾松纯林,平均树高16.1 m,平均胸径14.7 cm。林下植被以芒萁为主,伴有少量灌木,芒萁地上生物量741.6 g·m-2,盖度85%,马尾松年凋落量为535.8 g·m-2。
在不同恢复年限的马尾松人工林(Y0、Y13、Y31)分别随机设置3块样地,每块样地随机布设20 m×20 m的标准样方,每个样方分别设置8个用于测量土壤呼吸的PVC环(内径20 cm、高10 cm),PVC环埋入土壤5 cm,测量期间PVC环位置保持不变。于2015年4月至2016年3月,采用LI-8100开路式土壤碳通量测量系统(LI-COR,Nebraska,USA)测定土壤呼吸速率。于每月中旬选择晴朗的一天在9:00~12:00测定土壤呼吸速率,每个测定点测定时间为2 min。同时,采用时域反射仪(TDR)(Model TDR300,Spectrum Company,Aurora,USA)测量附近0~12 cm的土壤体积含水量,每个测定点周围测量3次,取平均值;采用长杆电子温度探针(SK-250WP,Sato Keir-Yoki,Kanda,Japan)测定附近土壤5 cm深处的土壤温度。
在每个样地内布置5个80 cm×80 cm凋落物框收集马尾松凋落物,每月收集一次,称其总重,然后取部分凋落物样品于65 ℃烘箱烘干,称其干重,根据含水量计算总凋落物生物量。2015年8月,用内径2.5 cm土钻在每个样地按“S”型随机取10钻,并混匀土样,采集完的土样保鲜,剔除石砾、根系等杂物,过2 mm筛,用于微生物生物量碳的测定,另一部分土样风干,研磨过0.149 mm筛用于土壤有机碳、全氮和全磷的测定。土壤总有机碳和全氮含量用碳氮元素分析仪(Elementar Vario MAX,Gemany)测定。土壤微生物生物量碳用氯仿熏蒸法浸提,土壤全磷含量采用HClO4-H2SO4法提取,用流动分析仪(Skalar san++,Netherlands)测浸提液。根系生物量:在每个样地用内径5 cm土钻随机取6钻,放水中浸泡1 h,清洗干净,并挑出大于2 mm细根,于65 ℃下烘干并称其干重,本研究所采用的数据为10 cm土壤深度细根。样地表层土壤基本理化性质如下表1。

表1 表层土壤(0~10 cm)理化性质Table 1 Physical and chemical properties of the soil in the 0~10 cm soil layer
1.3 数据处理
土壤呼吸速率RS的计算公式:

式中,T表示土壤5 cm深处温度,a、b为待定参数。
土壤呼吸的温度敏感性(Q10)的计算公式:

式中,b为式(1)计算的常量。
所有数据运用SPSS 22.0软件进行分析,采用重复测量方差(repeated-measures ANOVA)检验未治理地、恢复13 a和恢复31 a马尾松人工林之间土壤呼吸速率、温度和含水量月动态差异,采用单因素方差分析(one-way ANOVA)比较不同恢复年限土壤理化性质、凋落物量、季节的土壤呼吸速率和温度敏感性Q10值的差异。
利用结构方程模型(SEM)建立土壤温度、土壤养分、微生物及凋落物等因子与土壤呼吸及其温度敏感性之间的关系[2,10-11],分析植被恢复过程中影响土壤呼吸及其温度敏感性的主要因子,比较各因子的相对贡献。用Amos 21.0操作。
2 结 果
2.1 植被恢复对土壤温度和含水量的影响
土壤温度与大气温度具有一致的月变化规律(图2),不同季节间差异显著(图3),不同恢复年限土壤温度最高值出现在8月份,最低值出现在1~2月份(图2)。未治理地、恢复13 a和恢复31 a土壤温度变化幅度分别为7.11~36.96 ℃、12.17~27.92 ℃、11.07~31.95 ℃,植被恢复后,土壤温度变化幅度减小,且年均土壤温度下降了1.68~2.04 ℃。春季(3—5月)、夏季(6—8月)和秋季(9—11月)中,3~10月恢复13 a和恢复31 a土壤温度低于未治理地(图3),下降幅度范围为1.44~9.05 ℃,冬季(12—2月)土壤温度高于未治理地,其中1、2月份达到显著水平(P<0.05),增加幅度为0.58~5.20℃。土壤含水量最高值出现在5月份,最低值出现在8月份(图2)。重复测量方差分析显示,植被恢复并未显著改变土壤含水量(P = 0.136)。

图2 不同恢复年限气温(a)、土壤温度(b)、土壤含水量(c)和土壤呼吸(d)月动态Fig. 2 Monthly dynamics of air temperature (a), soil temperature (b),soil moisture (c) and soil respiration (d)relative to restoration history
2.2 植被恢复对土壤呼吸的影响
未治理地土壤呼吸速率并未出现显著的季节变化,植被恢复后土壤呼吸速率季节差异显著(图3),土壤呼吸速率最高出现在夏季(6—8月),最低值出现在冬季(12—2月)(图2和图3)。植被恢复后年均土壤呼吸速率显著高于未治理地(P<0.05),其中恢复13 a和31 a的马尾松林年均土壤呼吸分别是未治理地的2.52倍和2.62倍,而恢复13 a和31 a的马尾松林年均土壤呼吸速率没有显著差异(图3)。
2.3 土壤呼吸及其温度敏感性与各因子的关系
未治理地和植被恢复后土壤呼吸速率与土壤温度呈显著的指数相关(图4),而与土壤含水量不相关(P>0.05)。未治理地土壤温度可以解释土壤呼吸速率变化的25.29%,而恢复13 a和恢复31 a土壤温度分别能解释土壤呼吸速率变化的66.53%和48.75%。在R=aebT模型参数中,a反映了土壤的基础呼吸速率,从图4可见,恢复13 a和恢复31 a的土壤基础呼吸速率分别为1.030 μmol·m-2·s-1和1.149 μmol·m-2·s-1,显著高于未治理地的(0.726 μmol·m-2·s-1)。单因素方差分析显示,植被恢复后显著增加了土壤呼吸的温度敏感性(Q10)(P<0.05),未治理地Q10为1.27,而恢复13 a和恢复31 a Q10增加至1.66和1.58。
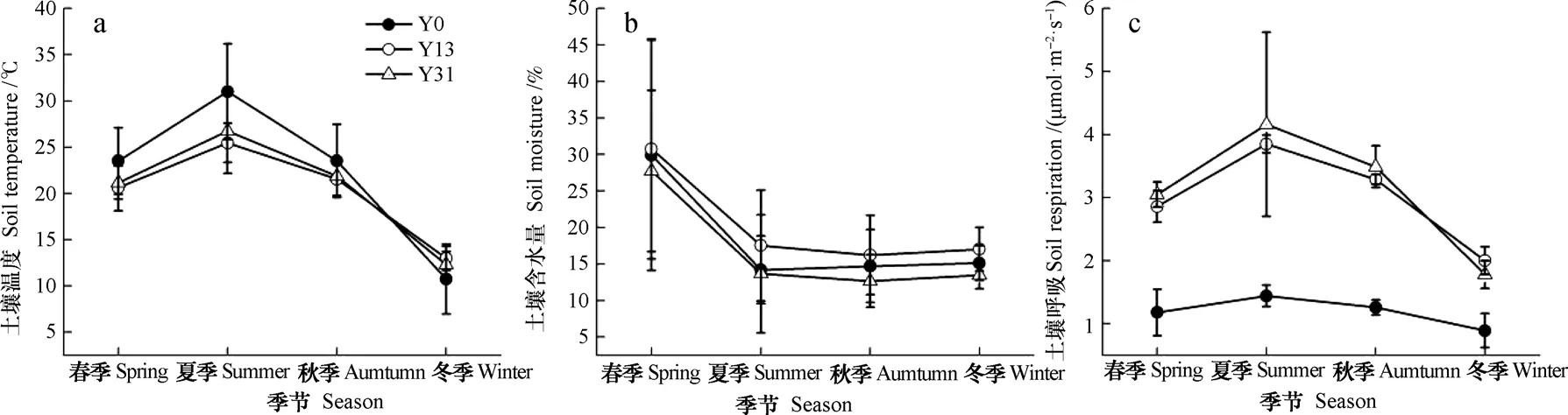
图3 不同恢复年限土壤温度(a)、土壤含水量(b)和土壤呼吸(c)季节比较Fig. 3 Comparison between seasons in soil temperature (a), soil moistureand (b) and soil respiration (c) relative to restoration history
SEM模型表明,模型对土壤呼吸和Q10分别解释了90%和82%。植被恢复所改变模型中的因子,凋落物量输入对土壤呼吸的响应系数最高(图5),能直接影响土壤呼吸,通过土壤碳氮比、微生物间接影响土壤呼吸。土壤呼吸和Q10与土壤温度呈负相关,凋落物量、土壤碳氮比和土壤微生物分别与土壤呼吸和Q10呈正相关(图5),其中,土壤微生物响应最高。
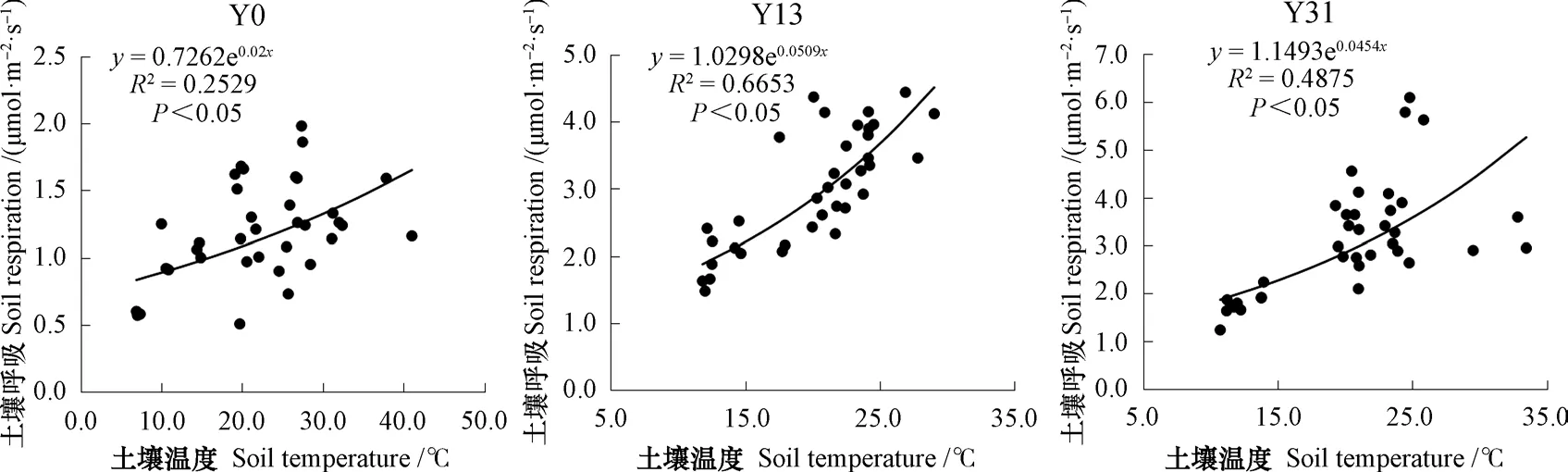
图4 不同恢复年限5 cm深度土壤温度与土壤呼吸的关系Fig. 4 Relationships between soil temperature of the 5 cm soil layer and soil respiration rate relative to restoration history
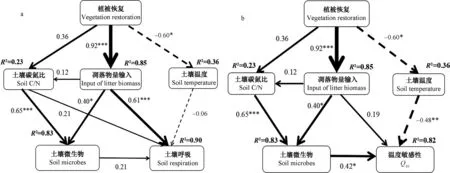
图5 结构方程模型模拟各因子对土壤呼吸及其温度敏感性的影响Fig. 5 Fitting of effects of various factors affecting soil respiration and its temperature sensitivity with the structural equation model
3 讨 论
3.1 植被恢复过程中土壤温度与土壤呼吸的关系
在亚热带地区正常年份,通常土壤呼吸与土壤温度存在较好的指数关系,与土壤含水量相关性较弱或不相关[12-14],这与本研究结果一致。未治理地土壤温度的季节变化幅度最大,但土壤呼吸速率在不同季节间并无显著差异,导致土壤温度与土壤呼吸的模型系数R2仅为0.25,低于亚热带大部分地区0.46~0.95范围[4,15-20],这意味着随着土壤温度升高,未治理地土壤呼吸难以增加,加剧了土壤呼吸影响因素的复杂性。而植被恢复后土壤呼吸季节差异显著,恢复13 a和恢复31 a的R2分别为0.67和0.49(表2),趋近于亚热带大部分森林生态系统水平,周文嘉等[21]也发现土壤温度解释了土壤呼吸变化的58.3%~90.2%。本研究中土壤温度与土壤呼吸的改善主要是由于植被恢复后,马尾松和芒萁盖度的增加,调节了土壤温度;另一方面,增加的生物量促进了土壤底物来源,如土壤有机碳、总氮等增加,因此,植被恢复有利于减缓未治理地逆境胁迫,增加土壤呼吸。
3.2 植被恢复对土壤呼吸的影响
土壤呼吸速率在植被恢复过程中不断增加,这与植被恢复改变了植被生物量、土壤有机碳数量和质量等有关(表1)。研究表明凋落物归还量、土壤有机碳和碳氮比分别与土壤呼吸显著相关[2-3,13-14],本研究中植被恢复能解释土壤呼吸90%(图5a)。凋落物量输入对土壤呼吸的影响最强,其变化作为植被恢复后生产力提高的直接体现,生态恢复促进了马尾松及林下植被数量的增加,提高了凋落物量输入。同时,模型中凋落物量输入与土壤微生物呈显著正相关,表明凋落物量的增加提高了微生物的底物来源,进而导致土壤呼吸增加。土壤碳氮比表示土壤有机碳质量的变化,能反映微生物利用的程度,本研究碳氮比与土壤呼吸正相关,未治理地碳源的匮乏限制了土壤呼吸,随着植被恢复土壤有机碳质量的提高,土壤可利用碳增加,刺激了微生物活性。土壤温度与土壤呼吸负相关,说明土壤温度对土壤呼吸的影响存在尺度依赖性,而根系生物量并未带入模型中,这是因为恢复31 a细根生物量低于恢复13 a,造成细根生物量与土壤呼吸关系较弱,这与相关研究并不一致[12-13]。值得注意的是,本研究中芒萁的数量在植被恢复过程中存在变化,未治理地芒萁生物量仅为39.13 g·m-2,恢复13 a和31 a后芒萁的盖度和生物量显著提高,其地上生物量分别为887.3 g·m-2和741.6 g·m-2。由于芒萁地下根系、凋落量输入改变了地下碳的质量和数量,进而影响了土壤呼吸的变化,这表明本研究中植被恢复对土壤呼吸的影响机制还需考虑芒萁的作用。
恢复13 a和恢复31 a的年均土壤呼吸速率分别为3.00 μmol·m-2·s-1和3.12 μmol·m-2·s-1,高于亚热带大部分人工林,但低于阔叶林[4,15-20]。排除系统误差,年均土壤呼吸速率的差异除了与不同林分类型、植被组成和观测频率等有关外,不同生长阶段、生产力和立地条件也影响着土壤呼吸的变化。恢复13 a和恢复31 a的土壤呼吸速率低于阔叶林可能与植物生理特征有关,阔叶林土壤呼吸速率一般较针叶林高10%左右[2],而马尾松林在该阶段(恢复13 a和31 a)土壤呼吸速率较高与其正处于速生阶段有关(有机碳含量低)[22-23]。恢复13 a和恢复31 a的土壤呼吸速率高于大部分地区的不同林龄阶段,但土壤有机碳、总氮和总磷供应低于大部分地区[4,15-20],这可能与本区域存在较大的养分胁迫有关。在养分胁迫可能会加剧微生物与植物之间的竞争,植物通过与微生物互利共生如菌根侵染与微生物从有机质中掠夺养分并释放大量胞外酶,加速土壤有机质矿化,而在可利用养分充足的情况下土壤微生物会减少木质素降解基因、酚氧化酶及过氧化物酶等难分解有机质信号的表达[24-26],减少土壤有机碳的分解[26]。
3.3 植被恢复对土壤呼吸温度敏感性的影响
未治理地土壤温度敏感性值(Q10)为1.27,而本研究区通过室内培养拟合的Q10范围为1.06~1.41[9],表明Q10存在季节变化。影响未治理地土壤呼吸的因素较为复杂,这可能与夏季高温加速土壤有机质消耗有关,土壤生物产生逆境适应,使得Q10接近1,而其他季节缓解了土壤呼吸对温度的响应,全年来看,未治理地Q10仍低于亚热
带大部分地区(表2)。植被恢复后Q10显著增加,变化范围为1.58~1.65,接近于Zheng等[27]整合的亚热带森林土壤(1.56~2.39)的范围。
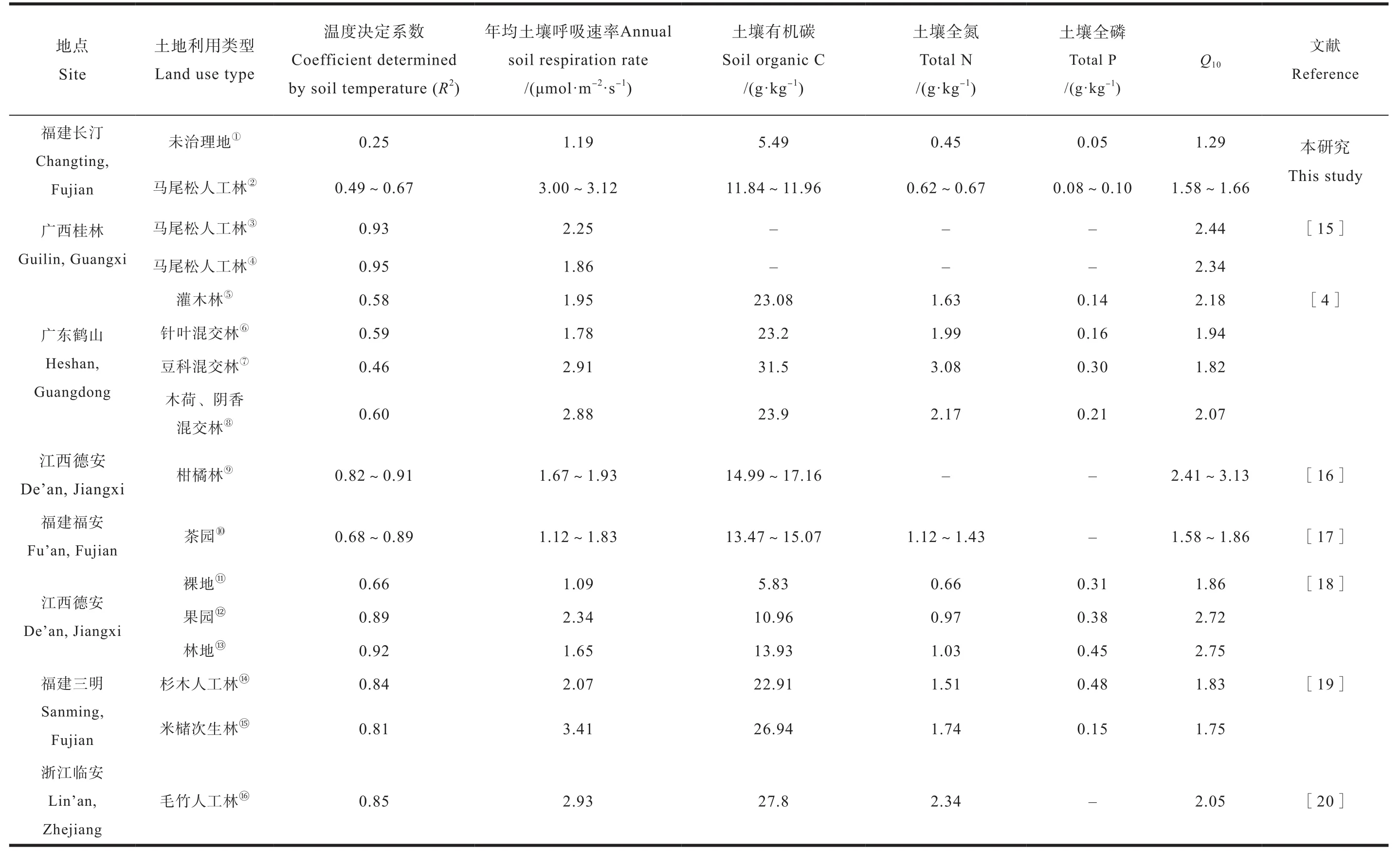
表2 亚热带季风气候区不同土地利用类型土壤呼吸及温度敏感性(Q10)的比较Table 2Soil respirationand its temperature sensitivity(Q10)in the subtropical monsoonclimate zone relative to land use
研究表明,Q10受土壤温度、根系生物量、微生物生物量、土壤有机碳数量与质量等因素的影响[6-7,28-29],而土壤呼吸温度敏感性模型显示,土壤温度、凋落物、土壤微生物和碳氮比均对Q10有重要影响(图5b),其中土壤温度与Q10呈负相关,说明土壤温度越高,Q10越低,这与前人结果研究一致[6-7,9]。未治理地可能加速土壤生物对底物的消耗,造成土壤有机碳降低,而植被恢复通过降低土壤温度,同时凋落物量又增加促进了底物来源,减缓了由于底物供应不足而导致的土壤呼吸下降,这对于增加土壤微生物丰度和改变微生物群落结构具有重要意义。土壤碳氮比通过影响微生物而间接影响Q10,这可能是因为未治理地土壤有机质、碳氮比较低影响着微生物细胞的构建。植被恢复过程中Q10并未随着恢复年限的增加而增加,这可能与活性碳存在较大的关系,土壤活性有机碳在恢复7~11 a显著提高,而在27~30 a有所下降[30-31]。此外,本研究区芒萁的盖度和数量并未随着恢复年限的增加而增加,这是否与芒萁覆盖下土壤微环境和土壤有机碳的变化有关还有待进一步研究。
4 结 论
植被恢复显著增加了土壤呼吸,与林地生产力提高有关;在季节尺度上,未治理地土壤呼吸无显著季节差异,土壤温度仅能解释土壤呼吸的25.3%,而植被恢复土壤呼吸的季节差异显著,土壤温度则解释了土壤呼吸速率的48.8%~66.5%,表明植被恢复对于增加Q10,缓解不利的土壤生物环境具有重要意义。
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
西北林学院学报(2022年5期)2022-10-04
农业工程学报(2022年8期)2022-08-08
北京大学学报(自然科学版)(2022年3期)2022-06-17
土壤(2022年1期)2022-03-16
新农业(2021年12期)2021-11-29
棉花学报(2020年3期)2020-08-08
农家科技中旬版(2020年10期)2020-07-12
绿色科技(2019年5期)2019-11-29
浙江农林大学学报(2019年6期)2019-11-13
