
以色列野生燕麦物候及农艺性状与起源生态地理因素的相关性
2019-02-25 08:21:14赖弟利王佳俊EVIATARNevo
麦类作物学报 2019年1期
范 昱,赖弟利,王佳俊,何 凤,赵 钢,EVIATAR Nevo,严 俊
(1.成都大学,农业部杂粮加工重点实验室/药学与生物工程学院,四川成都 610106;2.海法大学进化研究所,以色列海法31905)
燕麦(AvenasativaL.)是禾本科燕麦属草本植物,广泛分布于亚洲、欧洲、北美洲北纬40°以北的地区,是世界上第七大栽培作物[1]。野生燕麦(2n=2x/4x/6x=14/28/42)作为栽培燕麦的近缘植物,大多数分布于中东和北非地区,过去常被视作杂草,也作为青贮饲料[1]。燕麦野生近缘种具有丰富的遗传多样性,具有抗秆锈病、黄矮病、冠锈菌等特性,籽粒含有高蛋白、β-葡聚糖、赖氨酸、膳食纤维及矿质营养等成分[1]。发掘和鉴定野生燕麦种质资源,可为燕麦优质、抗逆新品种的选育提供优异基因材料,同时将提高我国麦类育种水平与育种效率[2-4]。目前,国内外学者对野生燕麦的少数优异性状或基因进行了研究与利用,如Ladizinsky等[5]发现欧洲四倍体大燕麦(A.magnaL.)和墨菲燕麦(A.murphiLadiginsky)蛋白含量高达25%~30%;Cavan等[6]发现野燕麦抗除草剂基因可能通过花粉进行传递;Yu等[7]研究了六倍体野生燕麦中抗除草剂相关基因ACCase的遗传、转录和体外酶活性;耿广东等[8]采用5 srDNA间隔序列分析和 SRAP标记验证以色列野生二粒小麦与光稃野燕麦远缘杂种的真实性;张庆勤等[9-11]利用野生二粒小麦、斯卑尔脱小麦、节节麦实现了与野生燕麦的远缘杂交培育了高抗病等优异材料。普通栽培燕麦被认为起源于地中海,由野红燕麦(A.sterilisL.)演变而来[1]。地处地中海新月沃地的以色列及周边广泛分布有不同生态群体野生燕麦,是栽培燕麦最大和原始的多样性基因保存地,而目前对以色列野生燕麦的研究少见报道。
不同地区间生态地理环境因素的差异会造成物种农艺性状的多样性[11]。野生燕麦在长期适应生态地理环境中,通过自然选择形成了其特有的物候及农艺性状的遗传多样性,演化产生不同特征群体[12-14]。因此,本研究以源自以色列7个不同地区的70个基因型的野生燕麦物候及农艺性状与生态地理因素进行相关分析,探讨起源地生态地理因素对野生燕麦生长特性的影响与塑造,以便进一步研究野生燕麦的遗传进化规律,为野生燕麦优异资源的开发利用提供参考。
1 材料与方法
1.1 试验材料
本研究使用的7个野生燕麦群体由以色列海法大学进化研究所协助采自以色列不同地区自然环境。群体起源地生态地理数据见表1。
1.2 田间试验
野生燕麦7个群体70个基因型种植于四川省成都市金堂县五凤镇成都大学试验基地。该基地地处东经104°51′,北纬30°61′,海拔436.4 m,年均气温17.1 ℃,无霜期283 d,年降雨量875 mm,空气相对湿度65%~70%,土壤为沙壤土,肥力中等,pH 6.11,有机质含量3.57%,全氮含量2.15 g·kg-1,全磷含量0.87 g·kg-1,全钾含量14.8 g·kg-1,水解氮含量110.0 mg·kg-1,有效磷含量4.3 mg·kg-1,有效钾含量116.0 mg·kg-1,铜含量44.1 mg·kg-1,锌含量59.9 mg·kg-1,锰含量463 mg·kg-1,硒含量0.149 mg·kg-1,铁含量3.15%。
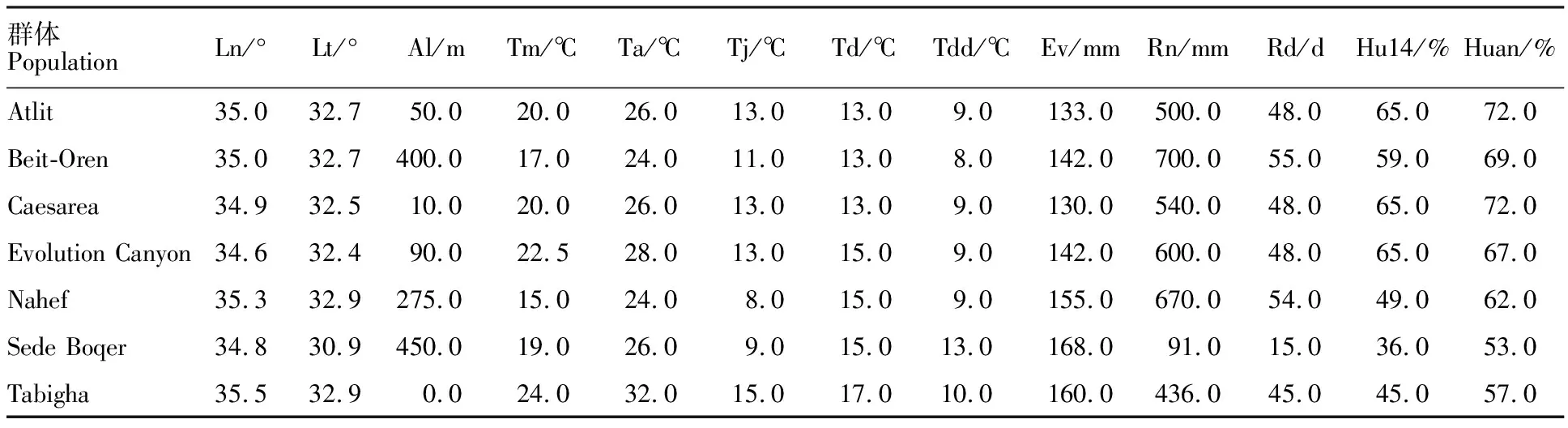
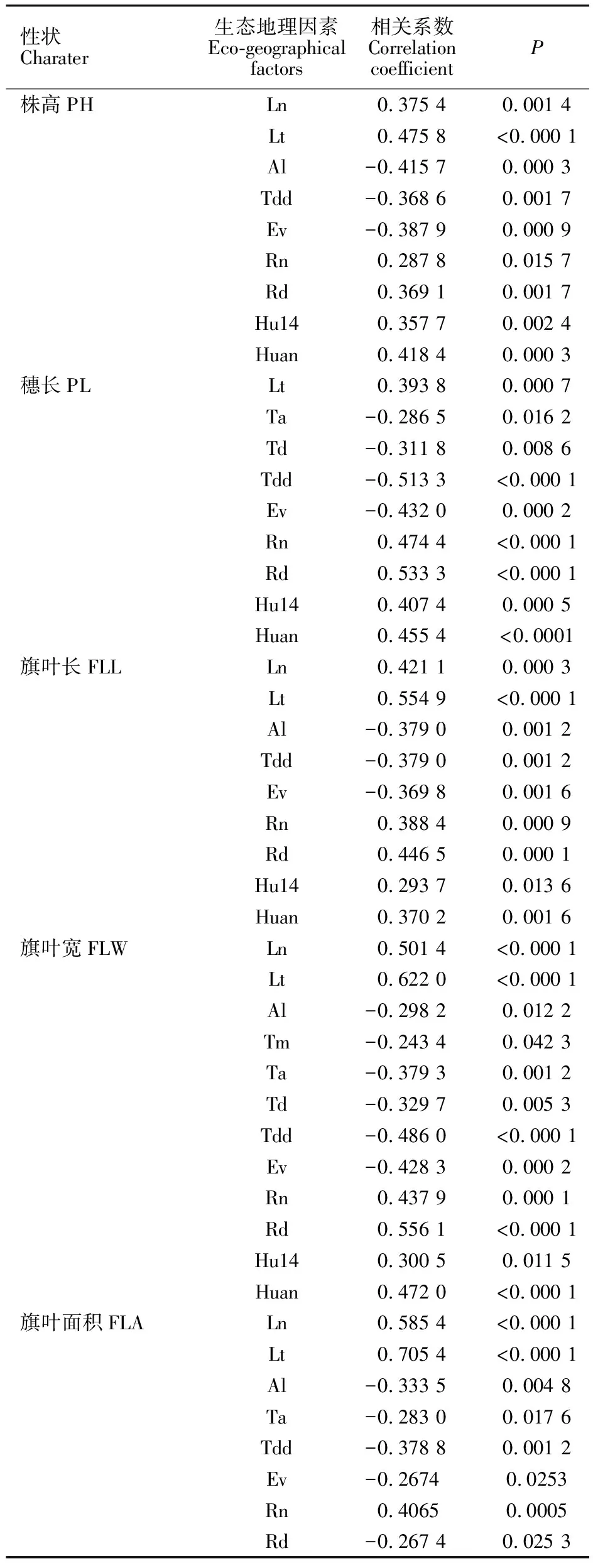
表1 7 个以色列野生燕麦群体起源地生态地理数据Table 1 Eco-geographical data for seven populations of wild oats in Israel
Ln:经度;Lt:纬度;Al:海拔; Tm:年平均温度;Ta:八月平均温度;Tj:一月平均温度;Td:季节温差;Tdd:昼夜温差;Ev:年蒸发量; Rn:年降雨量;Rd:年降雨天数;Hu14:14:00平均湿度;Huan:年平均湿度。下同。
Ln:Longitude; Lt:Latitude; Al:Altitude; Tm:The annual average temperature; Ta:The average temperature in August; Tj:The average temperature in January; Td:Seasonal temperature; Tdd:Temperature difference between day and night; Ev:Annual evaporation; Rn:Annual rainfall; Rd:On rainy days; Hu14:14:00 average humidity; Huan:Average annual humidity. The same in table 4 and 5.
2016年11月播种。从每个野生燕麦基因型中选出80粒大小均匀、饱满的种子,按随机区组试验设计进行穴播,每穴播种2粒,每行5穴,穴深10.0 cm,行长1.0 m,行宽20.0 cm,设置3个重复,并做保护行。待燕麦幼苗长出,每行定苗5株。全生育期集中隔离种植,2017年5-6月收获。其生长过程观察三叶期、分蘖期、拔节期、孕穗期、抽穗期、灌浆期和成熟期等物候期,测定株高、穗长、旗叶长、旗叶宽、旗叶面积、第一节间长、分蘖总数、有效分蘖数、生物产量、小穗千粒重、单株小穗数、种子千粒重等性状。
1.3 统计分析
用JMP 6.0软件(SAS Institute)进行方差(ANOVA)、斯皮尔曼秩(Spearmanp’s Rho)多元相关性分析,以及群体指标平均值差异显著性(P<0.05)Tukey-Kramer HSD检验;用R2.11构建燕麦相关性状间的皮尔森相关性矩阵(Pearson correlation matrix),并用Cytoscape 2.7.0软件对相关性矩阵进行网络分析(correlation-based network analysis,CNA)。
2 结果与分析
2.1 野生燕麦群体和基因型物候差异
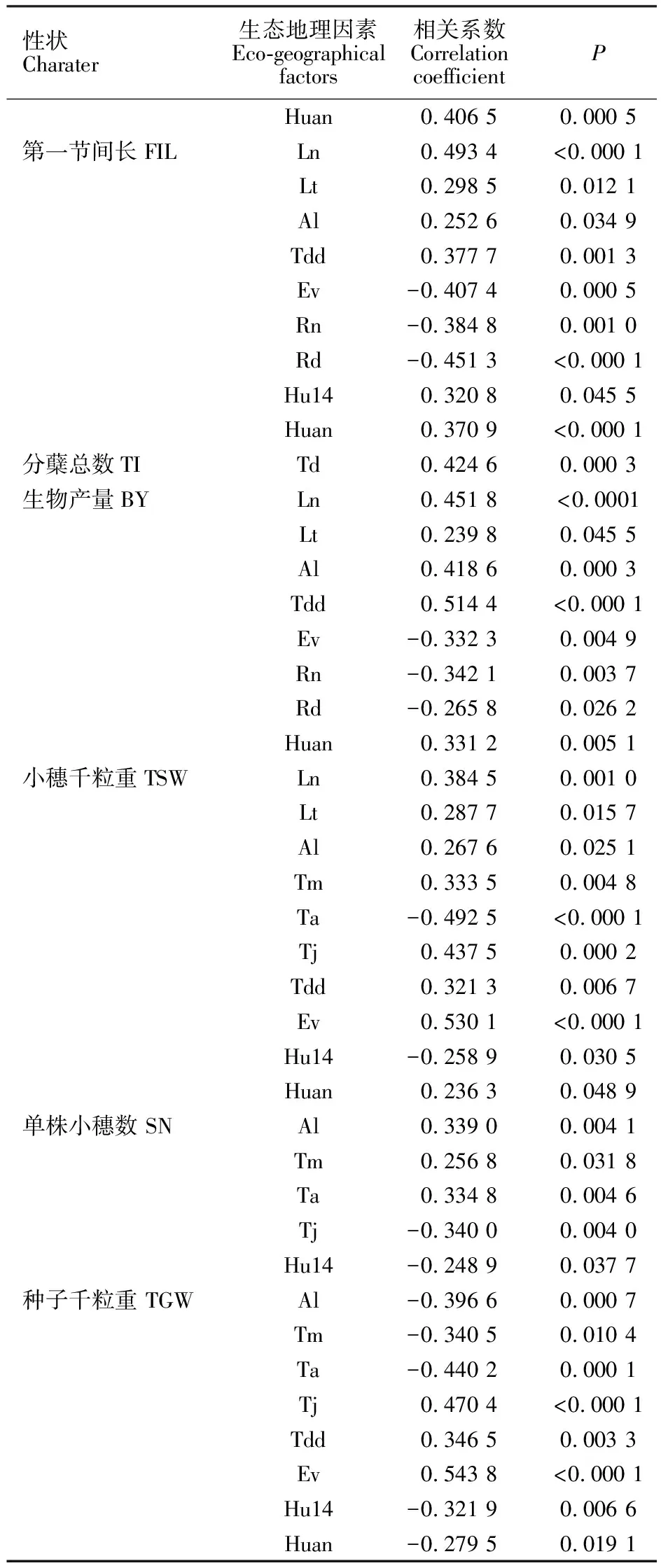
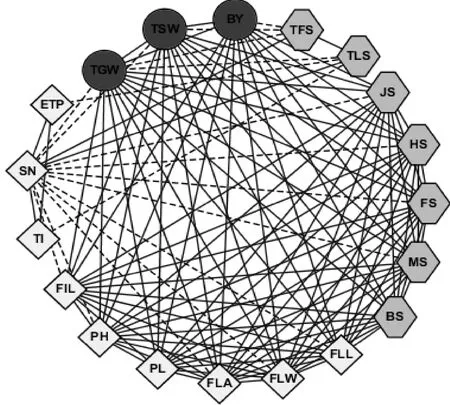
由表2可知,源自以色列7个地区(群体)野生燕麦物候期差异显著。三叶期最短的群体是Atlit,最长的是Caesarea;最早分蘖的是Evolution Canyon,最晚的是Sede Boqer;拔节期出现最早的是Sede Boqer,最晚的是Caesarea;孕穗期出现最早的是Sede Boqe,最晚的是Beit-Oren;抽穗期出现最早的是Sede Boqer,最晚的是Caesarea;群体灌浆期出现最早的是Sede Boqer,最晚的是Nahef;成熟期出现平均值表现为Sede Boqer 经Tukey HSD检验,在三叶期上,Caesarea群体显著晚于Atlit外,其余群体间均为同一水平;在分蘖期和抽穗期上,Sede Boqer与Evolution Canyon无明显差异,但仍保持了较低水平;在孕穗期、灌浆期和成熟期上,仅Sede Boqer均显著早于其他群体,而其他群体间差异均不明显。 经对7个野生燕麦群体的农艺性状进行方差分析,除分蘖总数及有效分蘖数(单株)外,株高、穗长、旗叶参数、第一节间长度、生物产量、小穗千粒重、单株小穗数和种子千粒重等性状间均存在显著差异。由表3可知,平均株高最大的群体是Atlit,最低的是Sede Boqer,其显著低于其余群体,而其余群体间无显著性差异。株高最大的基因型是来自Atilit 的Atl-04,达1 853.2 mm,最小的是来自Sede Boqer 的SB14-11,仅有694.0 mm。成熟时平均穗长最长的是Nahef,其显著高于穗短性状较为集中的Sede Boqer群体,其中最长的Nah-10为469.5 mm,最短的SB14-04仅202.0 mm。类似的,在旗叶长、宽和面积上,Nahef群体均显著高于Evolution Canyon和Sede Boqer群体,而来自Sede Boqer 的SB14-04的旗叶三性状均最低。第一节间长除Sede Boqer显著低于其他群体外,其他群体间均无显著性差异,其中最长的出现在Evolution Canyon中的ECg-16,达894.0 mm;最短的SB14-05,仅368.0 mm。单株生物产量上,除Sede Boqer群体显著低于其他群体外,其余群体间无显著差异,其中最大单株生物产量的基因型为Caesarea的C-06,高达268.1 g;最低生物产量为Sede Boqe的SB14-04,仅7.8 g。相反地,平均单株小穗数最高是Sede Boqer,显著高于其他群体,且其余群体间无显著性差异。其中单株小穗数最高的SB14-14,可达2 982.1;最少的是Evolution Canyon的ECg-03,为153.4。在所有的野生燕麦群体中,小穗千粒重和种子千粒重最大的均为Tabigha群体的Tg-09,分别为197.3和28.8 g;而Sede Boqer群体的种子普遍较小,SB14-22的小穗千粒重和种子千粒重最小,分别仅为4.3和1.5 g。 经斯皮尔曼秩相关性分析(表4),野燕麦三叶期仅与起源地海拔呈显著正相关,分蘖期仅与八月平均温度呈显著负相关;而拔节期、孕穗期、抽穗期以及灌浆期和成熟期等均与年降雨量、年降雨天数、14点平均湿度、年平均湿度呈显著正相关,与季节温差、昼夜温差、年蒸发量呈显著负相关。另外,孕穗期、抽穗期、灌浆期还与经度和纬度呈显著正相关,而与八月平均温度呈显著负相关。 表2 7个以色列野生燕麦群体物候期差异Table 2 Phenological difference of the seven populations of wild oats from Israe 同列平均值后,不同字母表示7个群体间差异显著(Tukey HSD检验,P<0.05)。表 3同。 Different letters after the means within the same columns indicate significant differences among the seven populations(P<0.05,by Tukey-Kramer HSD test). The same in table 3. 相关分析结果(表5)表明,农艺性状中,分蘖总数仅与季节温差呈显著正相关,其他性状均与经度、纬度、年降雨量、年降雨天数、14点平均湿度、年平均湿度呈显著正相关,与海拔、昼夜温差、年蒸发量呈显著负相关。类似的,穗长与纬度、年降雨量、年降雨天数、14点平均湿度、年平均湿度呈显著正相关,与八月平均温度、季节温差、昼夜温差、年蒸发量呈显著负相关,但与经度和海拔间均无显著的相关性。值得指出的是在旗叶参数中,旗叶宽和面积还与八月平均温度呈显著负相关。在产量因素中,小穗千粒重和种子千粒重均与年平均温度、一月平均温度、八月平均温度、14点平均湿度、年平均湿度呈正相关,而与海拔、昼夜温差、年蒸发量呈负相关;与之相反的是,单株小穗数与年平均温度、一月平均温度、八月平均温度、14点平均湿度、年平均湿度均呈负相关,与海拔呈正相关。 农艺性状和物候期之间的网络分析显示(图1),野生燕麦多个物候期和农艺性状相互间存在不同程度的显著相关性(P<0.05)。单株小穗数与分蘖总数、有效分蘖总数和三叶期、分蘖期呈显著正相关,与其余指标均呈显著负相关;分蘖总数与分蘖期、单株小穗数和有效分蘖总数呈显著正相关。有效分蘖总数和分蘖期呈显著负相关,与单株小穗数和分蘖总数呈显著正相关。种子千粒重与分蘖总数和有效分蘖数均无显著相关性,与三叶期和分蘖期呈显著负相关,与其余指标呈正相关。小穗千粒重与三叶期和单株小穗数呈显著负相关。生物产量与三叶期呈显著负相关,与分蘖总数和有效分蘖数无显著相关性。 表4野燕麦性状和生态地理因素的斯皮尔曼秩显著相关性 物候期Phenological period生态地理因素Eco-geographical factors 相关系数 Correlation coefficientP三叶期Trefoil stageAl 0.237 00.048 2分蘖期Tillering stageTa -0.254 20.033 7拔节期Jointing stageTd -0.295 40.013 0Tdd -0.558 7<0.000 1Ev -0.449 3<0.000 1Rn 0.568 9<0.000 1Rd 0.534 4<0.000 1Hu140.428 90.000 2Huan 0.419 50.000 3孕穗期Booting stageLn 0.268 50.024 6Lt 0.369 30.001 7Tm -0.290 00.014 9Ta -0.414 90.000 4Td-0.402 10.000 6Tdd-0.522 3<0.000 1Ev-0.392 00.000 8Rn0.475 5<0.000 1Rd0.542 9<0.000 1Hu140.273 00.022 2Huan0.424 80.000 2抽穗期Heading stageLt0.399 90.000 6Ta-0.310 00.009 0Td-0.347 70.003 5Tdd-0.561 6<0.000 1Ev-0.478 4<0.000 1Rn0.543 2<0.000 1Rd0.573 7<0.000 1Hu140.394 80.000 7Huan0.476 6<0.000 1灌浆期Filling stageLn0.293 50.013 7Lt0.448 00.000 1Ta-0.282 00.018 0Td-0.253 20.034 4Tdd-0.510 0<0.000 1Ev-0.392 10.000 8Rn0.512 2<0.000 1Rd0.546 2<0.000 1Hu140.338 50.004 2Huan0.399 90.000 6成熟期Maturity stageTd-0.392 70.000 8Tdd-0.374 80.001 4Ev-0.447 30.000 1Rn0.245 80.040 3Rd0.335 70.004 5Hu140.371 80.001 5Huan0.478 2<0.000 1 表5 野燕麦农艺性状和生态地理因素的斯皮尔曼秩显著相关性Table 5 Spearman’s Rhosignificant correlation between agronomic traits and birthplace eco-geographical factorsof wild oats (续表5Continuedtable5) 性状Charater生态地理因素Eco-geographical factors 相关系数 Correlation coefficientPHuan0.406 5 0.000 5 第一节间长FILLn0.493 4 <0.000 1Lt0.298 5 0.012 1 Al0.252 6 0.034 9 Tdd0.377 7 0.001 3 Ev-0.407 4 0.000 5 Rn-0.384 8 0.001 0 Rd-0.451 3 <0.000 1Hu140.320 8 0.045 5 Huan0.370 9 <0.000 1分蘖总数TITd0.424 6 0.000 3 生物产量BYLn0.451 8 <0.0001Lt0.239 8 0.045 5 Al0.418 6 0.000 3 Tdd0.514 4 <0.000 1Ev-0.332 3 0.004 9 Rn-0.342 1 0.003 7 Rd-0.265 8 0.026 2 Huan0.331 2 0.005 1 小穗千粒重TSWLn0.384 5 0.001 0 Lt0.287 7 0.015 7 Al0.267 6 0.025 1 Tm0.333 5 0.004 8 Ta-0.492 5 <0.000 1Tj 0.437 5 0.000 2 Tdd0.321 3 0.006 7 Ev0.530 1 <0.000 1Hu14-0.258 9 0.030 5 Huan0.236 3 0.048 9 单株小穗数 SNAl0.339 0 0.004 1 Tm0.256 8 0.031 8 Ta0.334 8 0.004 6 Tj -0.340 0 0.004 0 Hu14-0.248 9 0.037 7 种子千粒重 TGWAl-0.396 6 0.000 7 Tm-0.340 5 0.010 4 Ta-0.440 2 0.000 1 Tj 0.470 4 <0.000 1Tdd0.346 5 0.003 3 Ev0.543 8 <0.000 1Hu14-0.321 9 0.006 6 Huan-0.279 5 0.019 1 一般而言,作物长期生长在特定的环境条件下,因温度、雨量、地形、海拔、土壤和植被等因素差异,演化产生了不同特征群体与基因型,在自然选择压力的推动下,适应性进化并产生了丰富的遗传多样性[16-18]。本研究显示,以色列野生燕麦已分化形成了不同的生态类型,而这些生态群体的产生正是不同环境条件下野生燕麦长期适应其生境的结果。在以色列不同地区的7个野生燕麦群体中,来自南方内盖夫沙漠的Sede Boqer群体具有较短的全生育期,特别是其自拔节期起的物候期与北方的群体间产生极显著差异性(P<0.01)。北方野燕麦群体整体生育期较长,如群体Atlit、Beit-Oren,而北方群体相互间物候期和全生育期值相差不大,这说明物候期和生育期通常会受起源地地理因素的影响。但值得注意的是各群体在三叶期的时间上差异不显著(P>0.05),说明野生燕麦可能在萌发至三叶期的发育时间上存在一定的保守性。 图中所有相关性均达到显著水平(P<0.05) 。圆形和六边形代表野生燕麦的农艺性状,菱形为物候期,之间的连线代表性状之间的相关性,实线代表正相关,虚线代表负相关。图中各性状缩写见表2、3。 All correlations were significant at 0.05 level in network. The circles and hexagon represent the phenotypic characters,and the triangle represent the phenological. The edges between nodes indicate positive and negative correlations by solid and dotted lines,respectively. 图1野生燕麦群体的物候期与农艺性状数据相关性网络 Fig.1Correlationnetworksofwildoatsamongagronomictraitsandphenologicaldata 分析发现,起源地的水分、温度因子以及地理各因素单独或者共同对多数物候期和农艺性状起不同程度的作用。起源于经纬度较高、海拔较低、降雨较充沛、水分蒸发量少环境下的野生燕麦群体的生物产量及种子千粒重等较高,而海拔较高地区野生燕麦单株小穗数较高(表3、表4)。此外,野生燕麦的分蘖总数仅与环境季节温差有关,而与其他因子无关(表4)。与前人的报道一致[19-20],野生燕麦的旱生沙漠型相比北方湿生型,其籽粒更小,生长期较短,植株矮小,旗叶面积也稍小,灌浆期更短,但是种子数量更多。这是由于湿生型野生燕麦生长在水分充沛、光温适中的自然环境条件下,而旱生型生长环境高温少雨、土壤瘠薄,正是凭着粒小、粒多、成熟快并迅速掉粒于砂砾中的生态适应性反应,当面临极端干旱时,将大量种子存储于地下种子库,等待来年获得雨水后繁衍生息。 目前尚未见报道以色列野生燕麦种质资源在我国的引进情况。本研究结果表明,来自以色列七个不同地区野生燕麦70个基因型都能在成都平原地区自然气候条件下正常生长,说明以色列野生燕麦具有很强的适应性。由于成都平原与其起源地环境有很大的不同,势必会对物候及农艺性状产生一些影响,因而需要多年多环境试验,发掘野生燕麦稳定的优异性状及遗传基因。由于对地中海起源地野生燕麦的研究报道较少,不能做出更全面的比较。我国燕麦目录品种的株高一般为100.0~120.0 cm,最高的达到175.0 cm[15],而在本研究中野生燕麦高度范围在69.4~185.3 cm,说明较在编资源株高范围更宽。在我国西南平坝燕麦生态区,燕麦生育期一般为200.0~220.0 d,但在本研究中燕麦全生育期更短,仅为170.0~190.8 d。总之,野生燕麦作为理想的粮饲两用植物,其丰富的基因资源可用于温带禾谷类和牧草的改良,以及未来禾草类植物再生能源相关性状的生物学认识研究。本研究为进一步探究燕麦生长发育和遗传规律,以及种质资源创新提供了很好的参考依据。2.2 野生燕麦群体和基因型农艺性状差异
2.3 物候期和起源生态地理因素之间的相关性
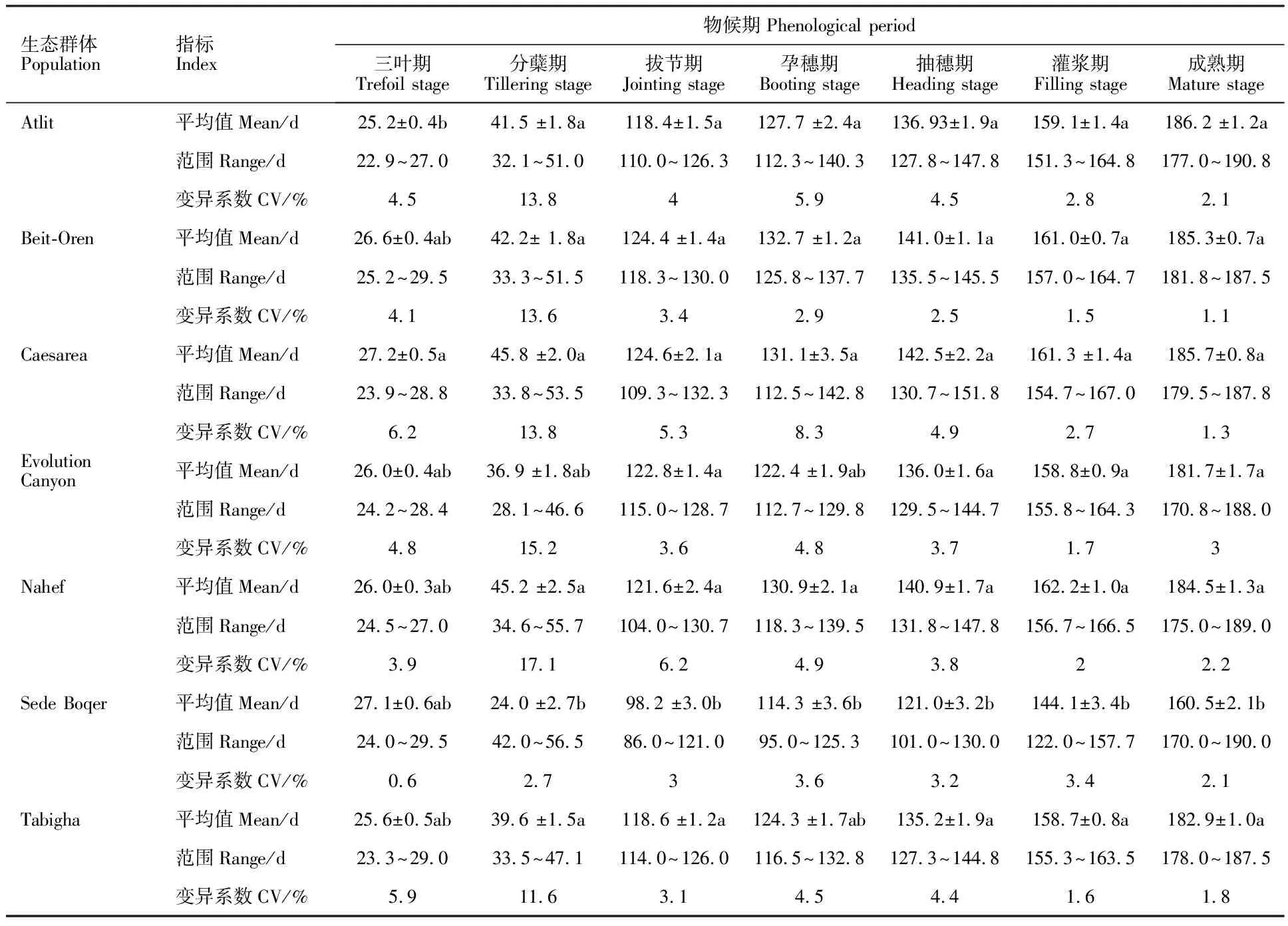
2.4 农艺性状和生态地理因素之间的相关性
2.5 物候期与农艺性状之间相关性
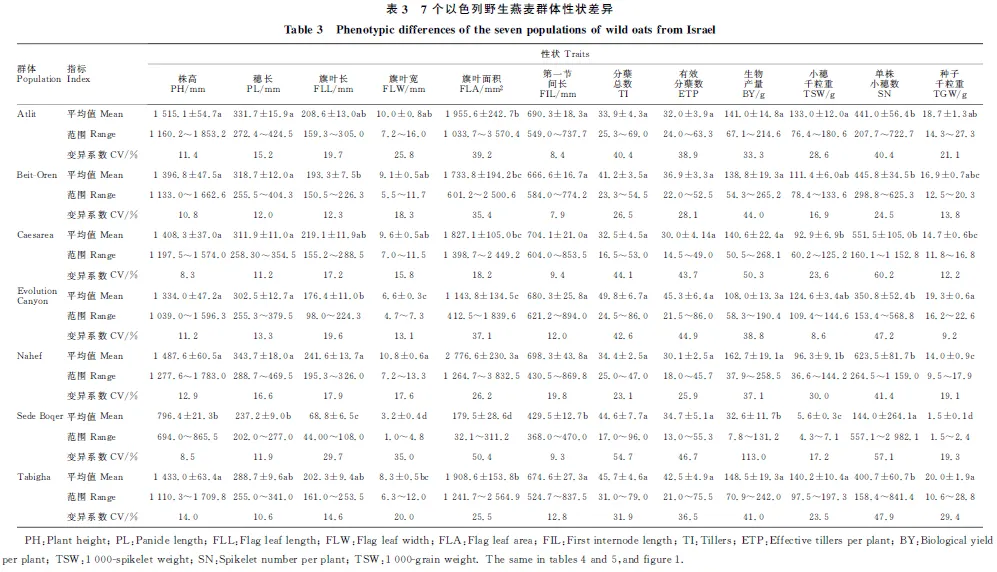
Table4Spearman’sRhosignificantcorrelationbetweenagronomictraitsandbirthplaceeco-geographicalfactorsofwildoats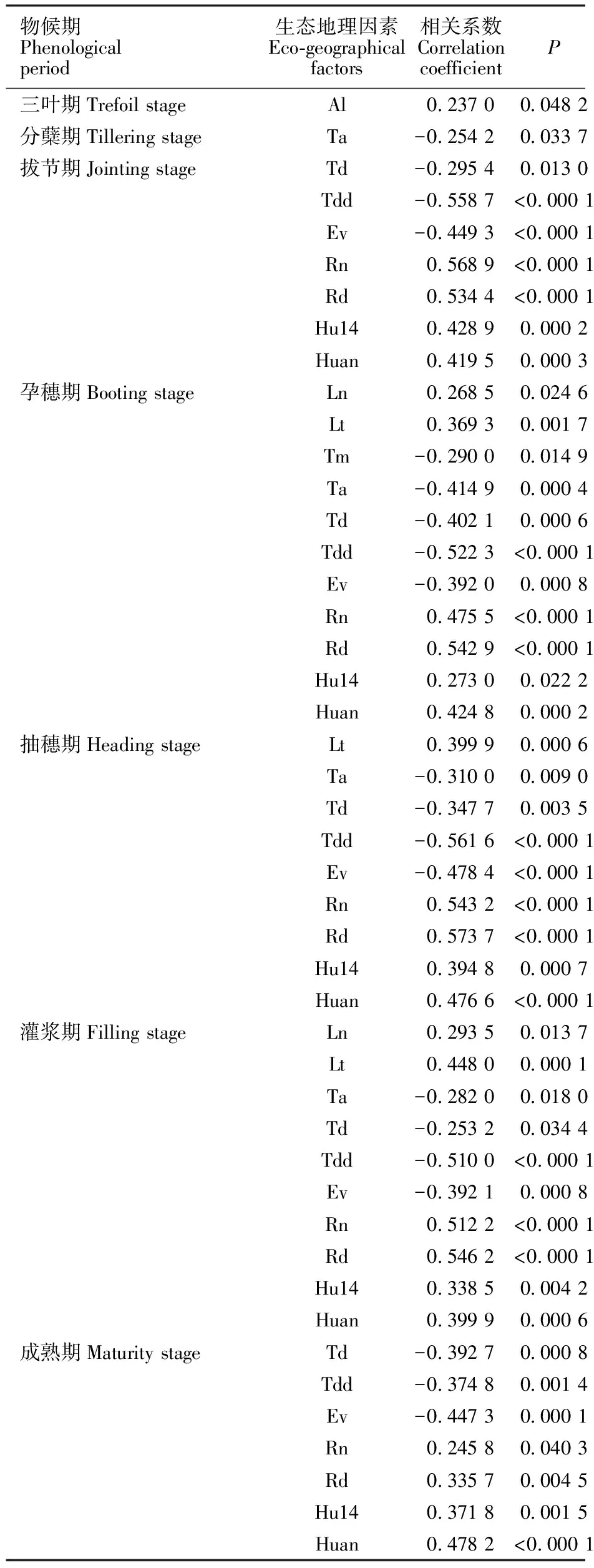
3 讨 论
猜你喜欢
环球时报(2023-03-21)2023-03-21 19:17:23
热带作物学报(2022年6期)2022-07-08 06:45:10
农业工程技术(2021年25期)2021-12-06 09:08:02
天津医科大学学报(2021年1期)2021-12-05 11:11:05
时代农机(2018年2期)2018-05-21 07:45:10
西南农业学报(2017年5期)2017-06-23 08:14:35
中国医学影像技术(2017年11期)2017-01-16 12:54:05
河北林业科技(2016年5期)2016-11-08 03:12:55
考试周刊(2016年63期)2016-08-15 14:33:26
西南农业学报(2016年5期)2016-05-17 05:42:22
