
Effects of potassium deficiency on photosynthesis, chloroplast ultrastructure, ROS, and antioxidant activities in maize (Zea mays L.)
2019-02-14 03:12DUQiZHAOXinhuaXlALeJlANGChunjiWANGXiaoguangHANYiWANGJingYUHaiqiu
DU Qi, ZHAO Xin-hua, XlA Le, JlANG Chun-ji, WANG Xiao-guang, HAN Yi, WANG Jing, YU Hai-qiu
1 College of Agronomy, Shenyang Agricultural University, Shenyang 110866, P.R.China
2 Bureau of Liaoning Provincial Seed Management, Shenyang 110034, P.R.China
Abstract Potassium (K) deficiency significantly decreases photosynthesis due to leaf chlorosis induced by accumulation of reactive oxygen species (ROS). But, the physiological mechanism for adjusting antioxidative defense system to protect leaf function in maize (Zea mays L.) is unknown. In the present study, four maize inbred lines (K-tolerant, 90-21-3 and 099; K-sensitive,D937 and 835) were used to analyze leaf photosynthesis, anatomical structure, chloroplast ultrastructure, ROS, and antioxidant activities. The results showed that the chlorophyll content, net photosynthetic rate (Pn), stomatal conductance(Gs), photochemical quenching (qP), and electron transport rate of PSII (ETR) in 90-21-3 and 099 were higher than those in D937 and 835 under K deficiency treatment. Parameters of leaf anatomical structure in D937 that were significantly changed under K deficiency treatment include smaller thickness of leaf, lower epidermis cells, and vascular bundle area,whereas the vascular bundle area, xylem vessel number, and area in 90-21-3 were significantly larger or higher. D937 also had seriously damaged chloroplasts and PSII reaction centers along with increased superoxide anion (O2·) and hydrogen peroxide (H2O2). Activities of antioxidants, like superoxide dismutase (SOD), catalase (CAT), and ascorbate peroxidase(APX), were significantly stimulated in 90-21-3 resulting in lower levels of O2-· and H2O2. These results indicated that the K-tolerant maize promoted antioxidant enzyme activities to maintain ROS homeostasis and suffered less oxidative damage on the photosynthetic apparatus, thereby maintaining regular photosynthesis under K deficiency stress.
Keywords: potassium deficiency, maize, photosynthesis, chloroplast ultrastructure, ROS and antioxidant
1. lntroduction
Potassium (K) is an essential element for plant growth and development, because it is involved in photosynthesis,enzyme activation, electrical neutralization, transport of metabolites, osmoregulation, and control of turgor pressure in living plant cells (Wang and Wu 2017). However,compared with nitrogen (N) and phosphorus (P), K is typically applied at a lower level during fertilization. An ongoing downward trend of negative K balance of about 60 kg ha–1yr–1appeared in intensive agricultural production areas starting in the late 1990s, and currently, more than 75% of soil in China is deficient in available K (Huet al.2016). Maize (Zea maysL.) is an important grain because it is an important source of feed, biofuel, and forage crop worldwide. A series of problems have emerged due to K deficiency in maize production systems, such as leaf chlorosis and reduced photosynthesis, causing subsequent inhibition of growth and development (Luet al. 2016).
Photosynthetic activity is known to decrease under K deficiency. Early chlorophyll degradation is very noticeable based on brown scorching and curling of leaf tips and margins as well as chlorosis (yellowing) under K deficiency(Tianet al. 2008; Römheld and Kirkby 2010). Transport of water and nutrients were found to be restricted by damaged anatomical structures from roots to shoots (Mattielloet al.2015; Sunet al. 2015), and the process of photosynthetic carbon dioxide uptake was impeded under K deficiency(Bednarzet al. 1998). However, changes in morphology and structure of chloroplasts were significantly related to chlorophyll content and photosynthetic capacity (Tianet al.2008). Under abiotic stress, dramatically low chlorophyll content, poor chloroplast ultrastructure, and restricted saccharine translocation inhibited the photosynthetic rate in plant leaves (Zhaoet al. 2001). Additionally, Cakmak(2005) concluded that leaf chlorosis caused by K-deficiency was related to oxidative degradation of chlorophyll by excess production of reactive oxygen species (ROS), a response to K deficiency. Stable morphology and structure of chloroplasts is an advantage contributing to tolerance in plants under K deficiency stress (Jiaet al. 2007; Shaoet al.2016). Ruanet al. (2015) also suggested low-K tolerant wheat could adjust development of anatomical structures to adapt to K-starvation through gene expression. However, a comprehensive understanding of how changes in anatomical structure and chloroplast ultrastructure affect photosynthesis is lacking under K deficiency in maize.
Under nutrient deficiency stress, oxidative stress occurs when the equilibrium between the production of ROS and antioxidant defense is disrupted (Shinet al. 2005). ROS is toxic to plant cells at high concentrations and thus excess oxidants must be scavenged to avoid deleterious effects(del Río 2015). Disturbance in electron transport during photosynthesis and respiration is a major source of abnormal levels of ROS (Apel and Hirt 2004). Evidence indicates that K deprivation stimulates ethylene production and results in further increases in ROS production (Shin and Schachtman 2004; Kimet al. 2010). Excessive accumulation of ROS will inevitably disturb physiological function in cells under long-term K deficiency. Plants possess ef ficient systems to scavenge ROS, which provides protection from destructive oxidative reactions. Antioxidative enzymes, such as superoxide dismutase (SOD) (Li C Let al. 2011), catalase(CAT), peroxidase (POX), or ascorbate peroxidase (APX)(Mittleret al. 2004; Qiet al. 2017), are regarded as the most important defense mechanisms for ROS. Enzymes that are associated with antioxidants, including ascorbic acid and glutathione, are highly ef ficient for detoxifyingand H2O2in cells (You and Chan 2015; Veronicaet al. 2017). Studies have also suggested that stronger photosynthetic ability is linked to higher antioxidant enzymes under K deficiency (Jiaet al. 2007; Chenet al. 2008).
Understanding the underlying physiological mechanisms in K-tolerant genotypes during photosynthesis is bene ficial for selecting K-tolerant parents in a traditional breeding program. In our previous study, we found that leaf senescence of 90-21-3 and 099, K-tolerant inbred lines,were significantly slower than that in D937 and 835,K-sensitive inbred lines. In addition, the photosynthetic rate in 90-21-3 was higher than that in D937 under low K stress at a later growth stage, which prolonged the functional period of leaves (Wanget al. 2012). However, we did not identify the cause or causes of internal physiological changes in leaves under K deficiency between different maize genotypes.Therefore, the aims of this study were to: (1) determine what regulates photosynthesis; (2) explain the variations in anatomical structure and chloroplast ultrastructure; and (3)identify the relationship of ROS and antioxidant activities between different tolerant genotype maize lines under K deficiency stress.
2. Materials and methods
2.1. Plant materials and treatments
Two groups of maize inbred lines, 90-21-3 (K-tolerant) and D937 (K-sensitive), 099 (K-tolerant) and 835 (K-sensitive),were compared from more than 2 000 lines cultivated for over 10 years in a K-deficient field (available K was less than 55 mg kg–1) (Caoet al. 2007), in which K fertilizer has not been supplied since 2000. The study was conducted in the long-term K fertilizer anchor pool (41°82´N, 123°56´E)of Shenyang Agricultural University in Liaoning Province,China, on 5 May 2016. The original K-deficient soil was obtained from Liaozhong County in Liaoning Province.The plough layer (20 cm) nutrient compositions were 11.87 g kg–1organic matter, 50.40 mg kg–1available K,98.50 mg kg–1alkali-hydrolysable N, 15.03 mg kg–1available P, and pH 7.3. Plants were fertilized with 150 kg ha–1urea(N, 46%) and 150 kg ha–1phosphorus diamine fertilizer(P2O5, 46%) as base manure before sowing, and 315 kg ha–1urea was replenished for nitrogen supply at the booting stage. According to K content in soil, sulfuric acid potassium fertilizer (K2O, 50%) was regulated the available K content to 130 mg kg–1in controls, while none applied in treated pools. Four rows of each inbred line were arranged on each experiment plot, with spacing of 0.50 m between rows, 0.30 m in a row, and row lengths of 3.5 m.
2.2. Pigment content determination
Chlorophyll was extracted according to Li X Tet al.(2011).A total of 0.5 g of fresh leaves was cut into small pieces and extracted using 80% acetone at –4°C for 48 h until they were blanched. The absorbance of the supernatant was recorded at 645 and 663 nm using a UV-spectrophotometer(UV-1800; Hitachi, Japan).
2.3. Leaf anatomical structure
Each 5 mm×5 mm leaf was cut in formalin-aceto-alcohol(FAA) solution containing 38% formaldehyde:glacial acetic acid:70% alcohol (1:1:18, v/v) for 24 h at 4°C to retain the natural leaf structure according to Mattielloet al. (2015), with some modifications. The fixed leaf pieces were dehydrated in ascending series of ethanol (70, 85, 95, and 100%), then cleared in xylene and embedded in paraf fin with the Epon 812 resin (Sigma, St. Louis, MO, USA), and finally cut into 10 mm sections using an Ultra-Thin Semiautomatic Microtome (Ultracut-UC7; Leica, Germany). Sections were dewaxed and dyed in 1% safranine O water solution and 0.5% Fast Green solution (95% alcohol dissolved), then observed and photographed at 100× using a microscope(ZEISS Axio Scope A1, Germany). The thickness of leaf and upper and lower epidermis cells, the vascular bundle (Kranz)area, and xylem vessel number and area, were randomly determined and analyzed by ZEN Blue Lite Software(n=25).
2.4. Chloroplast morphology and ultrastructure
To avoid structural differences in parts of leaves, pieces of leaves (1 mm×2 mm) from the margin of the middle eye leaves were cut into sodium phosphate buffer (PBS,0.1 mol L–1, pH 6.8), containing 2.5% glutaraldehyde (v/v)and 2% paraformaldehyde. Air was removed from the bottle in order to soak leaves completely in buffer solution according to Chenet al.(2004), with some modifications.The pieces were fixed at 4°C for 24 h, followed by three rinses of 15 min each with PBS, and post- fixed in 1% osmic acid (OsO4) for 1.5 h. The fixed samples were dehydrated for 15 min in an ascending series of ethanol dilutions with 50, 70, 80, 90, and 100% (three times). Then they were washed with tert-Butanol-ethanol solution (1:1, v/v) and tert-Butanol for 15 min, and finally embedded in Epon 812 resin (Sigma, USA). The sections were cut with an LKB-V ultramicrotome (LKB, Sweden) and stained with uranium acetate-lead citrate before being examined under a transmission electron microscope (HT7700; Hitachi,Japan) at 100 kV.
2.5. Gas exchange and chlorophyll fluorescence measurement
Gas exchange parameters were measured using a CIRAS-2 Portable Open- flow Gas Exchange System with an internal red/white LED light source (PP Systems, USA). The leaf chamber was set at an air flow rate of 100 mL min-1, 90%relative humidity, CO2concentration of (390±5) μmol mol-1,and photon flux density (PFD) of 1 500 μmol m-2s-1. Five eye leaves were used to measure net photosynthetic rate(Pn), transpiration rate (Tr), stomatal conductance (Gs), and intercellular CO2concentration (Ci).
Chlorophyll fluorescence parameters including minimal fluorescence (Fo), the maximal fluorescence (Fm), the maximum quantum ef ficiency of PSII photochemistry(Fv/Fm), actual photochemical ef ficiency of PSII (ΦPSII),photochemical quenching (qP), electron transport rate of PSII (ETR), and non-photochemical quenching (NPQ, NPQ=Fm/Fm´–1) were measured with an FMS-2 pulse modulated fluorometer (Hansatech, UK) on the same leaves adapted for 20 min in total darkness.
2.6. Oxidant production and lipid peroxidation
The H2O2level was detected using a method based on Tsaiet al.(2004). H2O2was extracted by homogenizing with 5 mL of 50 mmol L–1phosphate buffer (pH 6.8) including 1 mmol L–1hydroxylamine hydrochloride. The homogenate was centrifuged at 6 000×g for 25 min at 4°C. To determine the H2O2level, 3 mL of extracted solution was mixed with 1 mL of 0.1% titanium chloride in 20% (v/v) H2SO4, and then the mixture was centrifuged at 6 000×g for 15 min.The intensity of supernatant solution (yellow color) was measured at 410 nm. H2O2level was calculated with a known concentration and expressed as micromole per gram of fresh weight.
Malondialdehyde (MDA) content was determined with thiobarbituric acid as described in Miret al. (2015). A total of 0.5 g leaf tissues were homogenized with 5 mL of 5%trichloroacetic acid (TCA) and centrifuged for 20 min at 5 000×g. The supernatant solution (1.5 mL) was mixed with a solution containing 2.5 mL of 5% TCA and 0.5%thiobarbituric acid, then heated for 30 min at 100°C, and subsequently placed in an ice bath for cooling. The reaction mixture was centrifuged at 5 000×g for 10 min, and the resulting supernatant was used for determination of MDA content by recording the absorbance at 532 and 600 nm.
2.7. Enzyme extraction and assays
To extract antioxidant enzymes, 0.5 g of leaf tissue was homogenized in precooled 50 mmol L–1phosphate buffer(pH 7.8) with 0.1 mmol L–1EDTA and 2% (w/v) insoluble polyvinyl pyrrolidone. Then the homogenate was centrifuged at 10 000×g for 20 min at 4°C. The supernatant solution was used for enzyme assays, including SOD, CAT, POX,and APX (Chenet al. 2015).
SOD activity was measured as described by Netoet al.(2006). The reaction mixture (3 mL) contained 750 mmol NBT, 20 mmol ribo flavin, 130 mmol methionine, 100 mmol EDTA, 50 mmol L–1sodium phosphate buffer (pH 7.8),and 50 mL enzyme solution. The reaction mixture was illuminated at 4 000 lx for 20 min, while the control group was kept in darkness. The SOD activity was recorded at 560 nm and defined as the amount of SOD required to produce a 50% inhibition of reduction of nitrobluetetrazolium(NBT).
POX and CAT activities were measured following Tianet al.(2003). The reaction mixture (3 mL) consisted of 0.25%(v/v) guaiacol and 3 mmol L–1hydrogen peroxide in 100 mmol L–1sodium phosphate buffer (pH 6.0). 1 U of POX activity was defined as the increase in absorbance of 0.1 min–1at 470 nm g–1FW. CAT activity was detected in a 3-mL 50 mmol L–1sodium phosphate buffer (pH 7.0) containing 3 mmol L–1H2O2. 1 U was defined as the decrease in absorbance of 0.01 min–1at 240 nm g–1FW.
APX activity was measured according to Akcayet al.(2010). The reaction mixture (3 mL) consisted of 50 mmol L–1sodium phosphate buffer (pH 7.0), 0.25 mmol L–1ascorbate,1.0 mmol L–1H2O2, and 100 mL of the enzyme extract. 1 U of enzyme activity was defined as a decrease in absorbance of 0.01 min–1at 290 nm.
2.8. Statistical analysis
The experimental design was a completely randomized block design with three replicates (n=3) for each treatment. All data were subjected to an analysis of variance in SPSS18.0(SPSS Inc., Chicago, IL, USA), and the significance of difference between mean values were compared using the least significant difference (LSD) test at a 0.05 level of probability. All tables and figures were made using Excel 2013.
3. Results
3.1. Chlorophyll content
Under K deficiency, Chla, Chl (a+b), and Chla/bin 90-21-3 and 099 were slightly different than controls, whereas these values in D937 and 835 were significantly decreased at tasseling and flowering stages compared to controls(Table 1). Chlaand Chl (a+b) of D937 were significantly decreased by 13.99 and 9.03% in comparison to the control,17.92 and 14.65%, respectively, in 835. Moreover, Chla/bwas significantly decreased by 8.85 and 16.31% respectively due to the decreased Chlain D937 and 835. Chlbwas stable under K deficiency stress. Chlorophyll parameters of 90-21-3 and 099 were all significantly higher than those in D937 and 835 under K deficiency treatment.
3.2. Leaf anatomical structure
The leaf anatomical structure in 90-21-3 and D937 were clearly visible in Fig. 1, including upper epidermis cells,lower epidermis cells, vascular bundles and xylem vessels.Compared with the controls, the thickness of leaf was significantly decreased by 5.21% in D937 under K deficiency treatment, but was not significantly different in 90-21-3(Table 2). Both varieties were decreased in upper epidermis thickness and mesophyll cells, with significant decreases of 15.34 and 7.5%, respectively, in D937. There was nodifference in distance between bundles. The Kranz area and xylem vessel area were not significantly different from controls under K deficiency although they were slightly decreased by 5.54 and 12.90%, respectively, in 90-21-3,and by 8.09 and 13.57%, respectively, in D937. The vascular bundle area, and xylem vessel number and area in 90-21-3 were significantly higher than those in D937 under K deficiency.

Table 1 Chlorophyll content in 90-21-3, D937, 099, and 835 maize inbred lines grown under control and potassium deficient conditions

Fig. 1 Leaf anatomical structure in 90-21-3 and D937 grown under control and potassium deficient conditions. A–D, the picture of leaf anatomical structure of 90-21-3+K, 90-21-3-K, D937+K, and D937-K at 100×, respectively. UC, upper epidermis cells; LC,lower epidermis cells; Kr, Kranz; Ve, vessel; St, stoma; ET, epidermal tomentum; Bc, bulliform cells.

Table 2 Leaf anatomical structure in 90-21-3 and D937 grown under control and potassium deficient conditions
3.3. Chloroplast morphology and ultrastructure
Chloroplast morphologyNormal chloroplast morphology and structure in mesophyll cells (MCs) are necessary for ensuring normal leaf photosynthesis of maize. In this study, chloroplast from both lines was irregular, and chloroplast number in MCs was reduced accordingly under K deficiency (Table 3). Compared with controls, the number of chloroplasts in MCs was decreased by 8.91 and 20.61%in 90-21-3 and D937, respectively. In addition, edema was evident in some chloroplasts under K deficiency in 90-21-3 and D937. The external surfaces of chloroplasts were transformed from long and oval to elliptical or almost circular.Accordingly, the values of length/width (L/W) of chloroplasts were significantly decreased by 15.42% in 90-21-3, and by 32.64% in D937 under K deficiency due to the decreased length and increased width of chloroplasts. Compared with the controls, the length in D937 was significantly decreased by 13.41% and the width was significantly expanded by 28.22%, while in 90-21-3 the values were significantly decreased by 4.2 and 13.4%, respectively.
Chloroplast ultrastructureThe ultrastructure of chloroplasts was clearly visible in controls, with grana and stroma lamellae embedded in a stromal matrix and bounded by a double-membrane envelope for both genotypes(Fig. 2-A, B, G, and H). However, a pronounced difference in 90-21-3 and D937 was observed under K deficiency treatments. The external envelope and boundary, and the thylakoid systems were well-preserved in 90-21-3 (Fig. 2-D and E), although plastoglobuli were increased and swollen under K deficiency. In contrast, the ultrastructure of D937was seriously destroyed under K deficiency stress. The chloroplast was irregular, with substantially gathered and swollen plastoglobuli, and the lamellae structure was loose(Fig. 2-J and K). In these seriously damaged cells, the chloroplast envelopes were broken, the structure of lamellae was not normal, and most of them were dispersed in the cytoplasm for the K-sensitive genotype, with loose and vacuolated structure. The numbers of grana and grana
lamellae were significantly reduced in D937 by 30.04 and 33.49%, respectively, and by 10.03 and 9.32%, respectively,in 90-21-3 (Table 3). Mitochondria in 90-21-3 and D937 were decreased in size compared to the controls (Fig. 2-F and L).The membrane structure of mitochondria in D937 was blurry,and was more seriously damaged than that in 90-21-3.

Table 3 Chloroplast morphology and ultrastructure in 90-21-3 and D937 grown under control and potassium deficient conditions
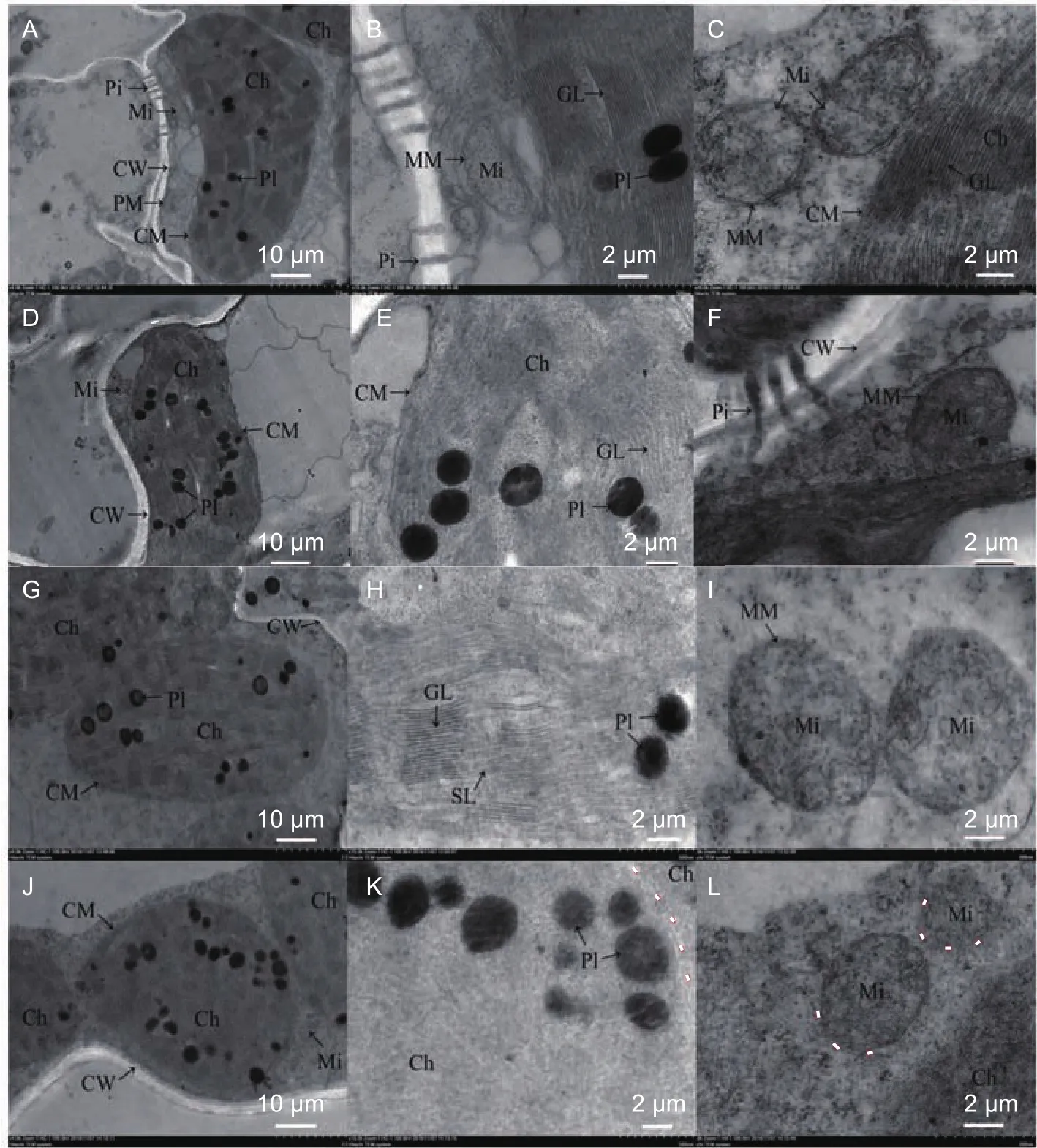
Fig. 2 Chloroplast morphology and ultrastructure in 90-21-3 and D937 grown under control and potassium deficient conditions. A,the complete picture of chloroplast of control 90-21-3 (×4 000). B and C, ultrastructure of chloroplast (×15 000) and mitochondria(×20 000), respectively, of control 90-21-3. D, E, and F, the complete picture of chloroplasts (×4 000, ×15 000) and mitochondria(×20 000), respectively, of 90-21-3 under treatment. G, H, and I, chloroplast (×4 000, ×15 000) and mitochondria (×20 000),respectively, of control D937. J, K, and L, chloroplast (×4 000, ×15 000) and mitochondria (×20 000), respectively, of D937 under treatment. Pi, piasmodesma; Mi, mitochondria; CW, cell wall; PM, plasma membrane; CM, chloroplast membrane; MM, mitochondrial membrane; GL, grana lamella; SL, stromal lamellae; PI, plastoglobuli; Ch, chloroplast. Broken line, boundary.
3.4. Gas exchange parameters
Varieties of gas exchange parameters of four inbred lines were affected by K deficiency stress (Table 4).Pn,Gs, andTrwere decreased andCiwas increased at the tasseling and flowering stages compared to the controls. In 90-21-3,Pn,Gs, andTrslightly decreased by 4.50, 8.76, and 10.74%,respectively, andCiincreased by 9.45% at the tasseling and flowering stages. In D937,Pn,Gs, andTrsignificantly decreased by 13.03, 20.00, and 28.08%, respectively,andCisignificantly increased by 19.09%. Compared with the controls, there was no significant difference inPn,Gs,andTrin 099 treated with K deficiency stress, while in 835 significantly decreased by 17.73, 35.00, and 31.90%. TheCiwas slightly increased in 099, and significantly increased by 24.97% in 835.
3.5. Chlorophyll fluorescence characteristics
Compared with the controls, parameters includingFm,Fv/Fm,ΦPSII,qP, and ETR, were more seriously decreased in D937 and 835 under K deficiency stress. The parameters significantly decreased by 11.53, 17.50, 14.29, 35.94, and 32.81% in D937, respectively, whileFoand NPQ increased by 10.25 and 30%, respectively (Table 5). And the first five parameters in 835 were significantly decreased by 14.07,28.25, 25.24, 23.13, and 25.24%, respectively, and the last two were increased by 6.29 and 24.88%, respectively.In 90-21-3 and 099, the first five parameters were slightly decreased under K deficiency stress, and the NPQ significantly increased by 18.60 and 34.52%, respectively.
3.6. ROS and MDA

Table 4 Gas exchange parameters in 90-21-3, D937, 099, and 835 grown under control and potassium deficient conditions
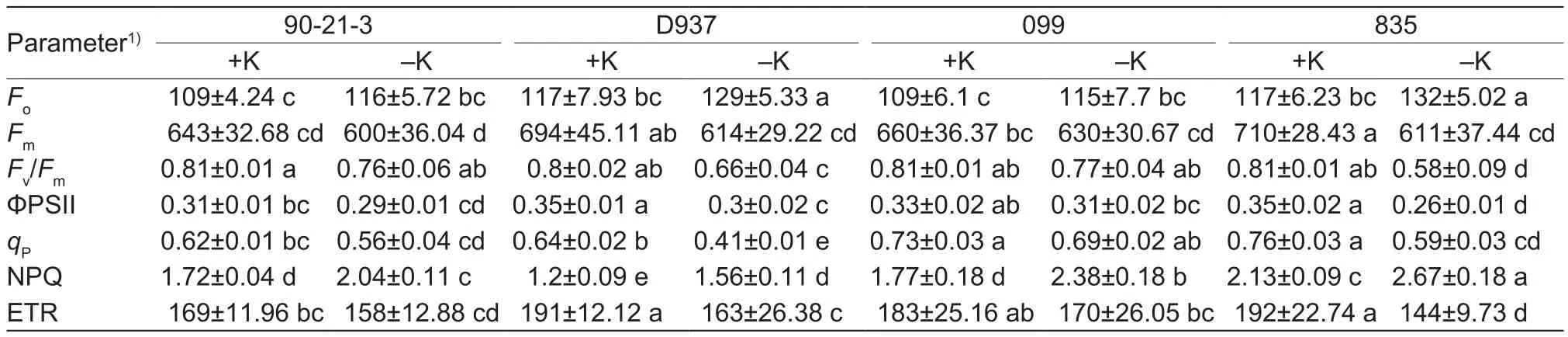
Table 5 Chlorophyll fluorescence in 90-21-3, D937, 099, and 835 grown under control and potassium deficient conditions
3.7. Antioxidant enzyme activity
Leaf antioxidant enzyme activity was stimulated in both lines over long-term exposure to K deficiency stress at tasseling and flowering stages (Fig. 4). Compared with the controls, SOD activity was significantly elevated by15.51% in 90-21-3; SOD activity was not significantly different between D937 and control (Fig. 4-A). POX activity was significantly increased in 90-21-3 and D937 at tasseling and flowering stages, by 43.41 and 32.18%, respectively, compared to the controls (Fig. 4-B). A significant increase of 69.48%in CAT activity was observed in the leaf of 90-21-3 under K deficiency stress, but there was no difference in D937(Fig. 4-C). APX activity increased in 90-21-3 and D937 in response to K deficiency. And it was significantly increased in 90-21-3 by 30.31%, and by 12.53% in D937 (Fig. 4-D).
4. Discussion
4.1. Photosynthetic parameters

Fig. 3 Accumulation of reactive oxygen systems (ROS) and malondialdehyde (MDA) in 90-21-3 and D937 grown under control and potassium deficient conditions. A–C are differences of superoxide anion hydrogen peroxide (H2O2), and MDA contents,respectively in both inbred lines at the tasseling and flowering stages. The bars are the standard errors of the mean; the letters represent significant differences at the P<0.05 level.

Fig. 4 Antioxidant enzyme activity in 90-21-3 and D937 grown under control and potassium deficient conditions. A–D are differences in superoxide dismutase (SOD), peroxidase (POX), catalase (CAT), and ascorbate peroxidase (APX) activities, respectively, in both inbred lines at the tasseling and flowering stages. The bars are the standard errors of the mean; the letters represent significant differences at the P<0.05 level.
Photosynthesis, the basis of plant growth, development and yield, includes light harvesting, PSII photochemistry,and CO2assimilation. Previous studies have shown that nitrogen, sulfur, and iron deficiencies could directly block the synthesis of protein complexes in photosynthetic reactions(Kalajiet al. 2014). LeafPndecreased under K deficiency stress in soybean, cotton, and maize, resulting in suppressed electron transfer energy, photosynthetic enzyme activity, and nitrogen metabolism enzyme activity (Quet al.2011; Wanget al. 2012, 2015; Huet al. 2017). Chlorophyll is the main photosynthetic pigment in plants. Chlorophyll synthesis is stimulated by K application, but stopped by increasing ethylene and abscisic acid under K deficiency(Lawansonet al. 1977). In the present study, decreases inPnandGs, and increases inCiwere more severe under K deficiency stress in D937 and 835 than in 90-21-3 and 099.However, K-tolerant maize, 90-21-3 and 099, had higherPnandGsthan D937 and 835 under K deficiency. In addition, K deficiency reduced chlorophyll in D937 and 835, especially Chla, but did not affect chlorophyll in K-tolerant 90-21-3 and 099. A similar result was reported in cotton (Huet al.2016). These data indicate that K-tolerant maize, 90-21-3 and 099, could maintain normal photosynthesis with higher chlorophyll by adjusting the stomata conductance to avoid increases in the intercellular CO2concentration.
Chlorophyll fluorescence parameters could provide precise information about the state of the photosynthetic apparatus and especially of PSII (Sunet al. 2015).Fv/Fmis a major indicator of photoinhibition or injury in the PSII complexes (Bednarzet al. 1998; Li X Tet al. 2011).qPis used to measure the utilization rate of energy absorbed by antenna pigment of PSII for photosynthesis, and also re flects the opening state of the PSII reaction center (Bilger and Björkman 1990). In the present study, theFv/Fm,ΦPSII,qP, and ETRwere significantly decreased in D937 and 835, but there were minimal changes in 90-21-3 and 099.The decreasedFv/Fmin D937 and 835 under K deficiency was due to the reduction ofFm,which indicated damage to the reaction centers of PSII, hindering electron transport.The decreased ETRdirectly indicated the disorganized electron transport chain, which resulted in free radicals,and damage of chloroplast membranes and changes in chloroplast structure
4.2. Leaf anatomical structure and chloroplast structure
Normal leaf anatomical structure and chloroplast structure are particularly bene ficial for photosynthesis (Liet al.2017). Damaged anatomical structure could in fluence the transport of water and nutrients from roots to shoots, and eventually impede the process of photosynthesis under nutrient deficiency (Mattielloet al. 2015; Sunet al. 2015).However, low-K-tolerant wheat could adjust its anatomical structure development to alleviate negative effects of K-starvation (Ruanet al. 2015). In this study, leaf thickness,mesophyll cells, lower epidermis cells, and Kranz area were significantly decreased in D937 under K deficiency stress.Although the Kranz area and xylem vessel area were also decreased by K deficiency, 90-21-3 had significantly higher area than D937, and thus retained higher transport capacity for water and nutrients from roots to shoots (Mattielloet al.2015). In addition, morphological and structural changes in chloroplasts were important for the decrease of chlorophyll content and photosynthetic capacity (Shaoet al. 2014).Dysfunction of chloroplasts was caused by degraded envelope, irregular chloroplast morphology and decreased grana and thylakoid lamellae due to abiotic stress (Xuet al.2008; Renet al. 2016). Zhaoet al. (2001) indicated that the function of chloroplasts was affected in cotton leaves under K deficiency stress, resulting in an increase in starch grains and plastoglobuli and decreased grana in chloroplasts. In this study, chloroplasts in 90-21-3 leaves had more welldefined membranes and thylakoids with smaller numbers of plastoglobuli, whereas chloroplasts in D937 were destroyed with deliquescent and fuzzy membranes and thylakoids. As a result, the photosynthetic process was decreased and chlorophyll content and chlorophyll fluorescence parameters were reduced, eventually resulting in the degradation of leaf photosynthetic capacity.
4.3. ROS and antioxidant activities
ROS was rapidly generated when crops were exposed to abiotic stress, especially in chloroplasts (Xuet al. 2006;Ahanger and Agarwal 2017). ROS led to disordered regular cell structure and function by damaging nucleic acids,oxidizing proteins, and causing lipid peroxidation (Foyer and Noctor 2005). Long-time K deficiency resulted in severe membrane lipid peroxidation due to high ROS production and accumulation in tomato (Hernandezet al. 2012). In this study,and H2O2were increased by K deficiency, with more accumulation in D937 than in 90-21-3. MDA contents indicated more severe membrane lipid peroxidation in D937 than that in 90-21-3 under K deficiency. By scavenging the abnormal ROS, enzymatic antioxidant systems were considered as primary regulators for scavenging and eliminating ROS in the plant cells. First, SOD could specially catalyze the conversion ofinto H2O2, acting as the first line of defense against the potential toxicity of superoxide radicals. Then, other antioxidant enzymes catalyzed the subsequent breakdown of H2O2to water (H2O) and oxygen(O2) (Garrattet al. 2002). Higher SOD enzyme activity in salt-tolerant rice was important for eliminating ROS as salt concentration increased (Chawlaet al. 2013). In our other study, we found higher SOD enzyme activity and lower MDA in roots of 90-21-3 under K deficiency stress (Zhaoet al.2016). CAT is a principal enzyme for H2O2scavenging under salt and P deficiency (Netoet al. 2006; Chenet al. 2015). In the present study, K deficiency stimulated antioxidant activity in 90-21-3 and D937. The increased activity of SOD in the K-tolerant line, 90-21-3, played essential roles in scavenging or eliminatingunder K deficiency. Also, CAT and APX in 90-21-3 were higher than those in the K-sensitive line, D937,under K deficiency. H2O2concentration in 90-21-3 and D937 leaves was considerably higher than that in the controls with increased CAT, POX, and APX activities. However,higher CAT and APX activities and lower MDA was observed in 90-21-3, although POX activity was increased in both lines. This result suggested that CAT and APX enzymes played a major role in scavenging H2O2in leaves under K deficiency stress. These results indicated that K-tolerant maize, 90-21-3, was able to effectively stimulate activity of SOD enzymes and coordinate expression of CAT, POX,and APX, alleviating membrane lipid peroxidation under K deficiency stress.
5. Conclusion
Under K deficiency stress, the K-tolerant maize, 90-21-3 and 099, maintained higher chlorophyll content, and had stable morphology and chloroplast structure, all of which led to higher photosynthetic ability compared to D937 and 835.In contrast, the reaction centers of PSII and electron transfer in D937 and 835 were inhibited under K deficiency stress,which resulted in severe membrane lipid peroxidation due to accumulation ofand H2O2. Compared with K-sensitive maize, antioxidant enzyme activity in K-tolerant maize was increased, which promoted scavenging of ROS under K deficiency stress and alleviated oxidative stress, allowing normal photosynthesis.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (31771725 and 31301259) and the National Key Technology R&D Program of China(2013BAD07B03).
 Journal of Integrative Agriculture2019年2期
Journal of Integrative Agriculture2019年2期
- Journal of Integrative Agriculture的其它文章
- Modelling and mapping soil erosion potential in China
- Updating conventional soil maps by mining soil–environment relationships from individual soil polygons
- Spatial variability of soil total nitrogen, phosphorus and potassium in Renshou County of Sichuan Basin, China
- Spatial variability of soil bulk density and its controlling factors in an agricultural intensive area of Chengdu Plain, Southwest China
- An integrated method of selecting environmental covariates for predictive soil depth mapping
- Remotely sensed estimation and mapping of soil moisture by eliminating the effect of vegetation cover