
月季F1代群体表型性状变异分析
2019-02-14 09:11:16周利君万会花潘会堂张启翔
植物研究 2019年1期
周利君 于 超 常 笑 万会花 罗 乐 潘会堂 张启翔
(花卉种质资源创新与分子育种北京市重点实验室,国家花卉工程技术研究中心,城乡生态环境北京实验室,北京林业大学园林学院,北京 100083)
月季(Rosaspp.)是蔷薇科(Rosaceae)蔷薇属(RosaL.)的一个重要类群[1]。通过蔷薇属内种间反复杂交和长期选育,形成了色彩绚丽的现代月季杂交品种群[2],被誉为“花中皇后”。同时,作为我国十大传统名花之一,月季栽培历史悠久、来源广泛、种类丰富、适应性强、栽培养护简单、生产繁殖相对容易,园林应用形式多样且商品用途众多[3]。基于其高观赏价值和可观的经济价值,月季一直是全世界的生产商和研究者所关注的对象[4]。中国是蔷薇属植物的分布中心,拥有栽培历史悠久的古老月季品种资源,是进行月季品种改良的重要遗传资源[5],将其和遗传背景复杂的现代月季优良品种杂交,有利于利用杂种优势进行种质创新,培育具有自主知识产权的月季新品种[6~7]。
植物大多数观赏性状都是由微效多基因控制的数量性状,在一个自然群体或者同一杂交后代群体的不同个体中往往表现出连续的数量差异,易受环境影响,不易明确分组归类[8],需要应用统计学方法对整个群体进行的系统测定和分析[9]。由于蔷薇属观赏植物遗传组成上高度杂合和复杂、染色体倍性水平差异较大、种子高度木质化及后熟造成萌发困难,通过常规育种手段对其观赏性和抗性相关的数量性状研究较为困难[10]。以构建高密度遗传图谱为基础进行数量性状位点QTLs(Quantitative Trait Locus)定位成为数量性状遗传研究的常用方法,目标性状相关遗传信息及等位基因的跟踪和选择可以通过构建图谱得以实现[11]。
目前,蔷薇属植物的花瓣数量、花期、花香、花色、花径、生长势、皮刺、株形和抗白粉病等多个农艺性状进行了QTLs定位,但是目前直接对二维性状花冠面积以及细部性状花瓣长宽等研究报道较少。另一方面,植株叶片的形态特征,如叶长、叶宽不仅影响植株的观赏效果,还与植株光合产物积累相关,也与花朵的表现形态也有一定关联性[12];本研究利用统计学方法对月季杂交群体的表型性状进行测定分析,了解和掌握该群体花部7个具体性状以及叶片2个表型性状的遗传变异规律及相关性,以期为月季表型性状的QTLs定位提供理论基础和参考数据,对于蔷薇属植物优良基因的挖掘及辅助选择育种具有实际意义[13]。
1 材料与方法
1.1 试验材料与试验地概况
2011年4~6月,以1株四倍体古老月季品种‘云蒸霞蔚’(Rosa‘Yunzheng Xiawei’)为母本,收集另1株四倍体现代月季品种‘太阳城’(R.‘Sun City’)的花粉进行杂交,采收成熟果实于2012年3月进行播种,获得333株F1代实生苗群体。该群体露地种植于北京市昌平区国家花卉工程技术研究中心苗圃,年平均日照时数2 684 h,年平均气温11.8℃,年平均降水量550.3 mm,个体栽植点的温度、光照、水分、土壤等环境因子基本相同,生长健壮、表现稳定。随机选择184株杂交F1代群体和杂交亲本作为研究对象(表1)。

表1 试验材料主要信息
1.2 试验方法
该群体2个集中花期(2016年9月、2017年5月)内,测定2株亲本及184株子代的花部和叶片性状(表2)。
1.2.1 花部性状的测量方法
每个单株上选3朵开放程度到图1所示第五个阶段的花朵,分别进行如下性状的测量:
(1)花朵直径:测量如图2所示,过辐射对称中心测量花朵直径,每朵花测量3次。
(2)花冠面积:采用图片采集法[14]。测量时,将直尺与花俯视角度的图像位于同一平面(与地表平行)拍照,后期利用Adobe Photoshop 2015进行处理。根据直尺上的标记确定参照面积RA(Reference substance Area)=1 cm2,记录参照面积所占用的像素RP(Reference substance Pixel),使用磁性套索工具,选择待测花冠面积,记录待测面积所占用的像素,记为FP(Flower Pixel),依据待测面积/参照面积=待测面积像素/参照面积像素的比例关系,即FA=RA×FP÷RP,计算待测花冠面积FA(Flower Area)=FP÷RP(单位:cm2)。
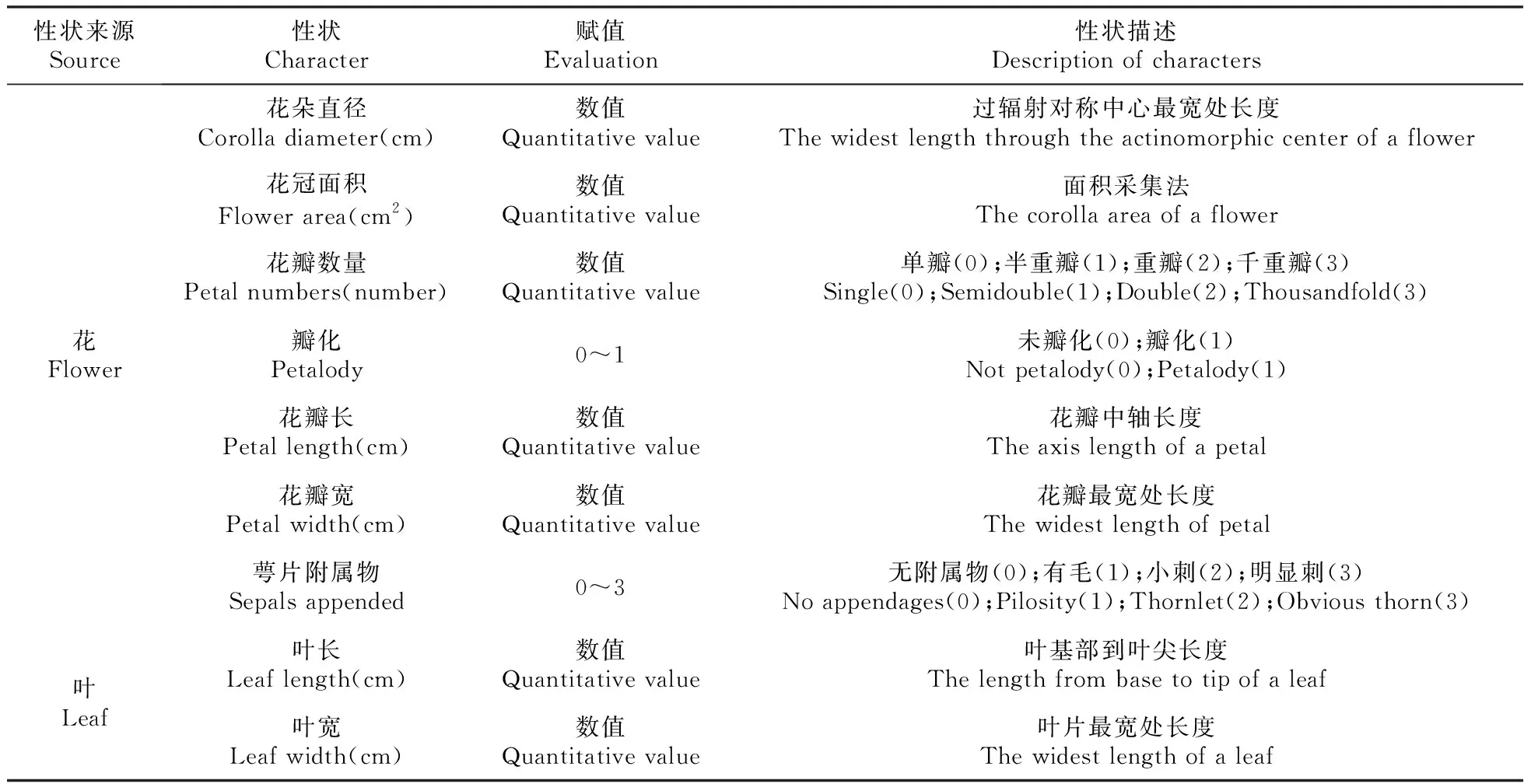
表2 性状选取与测定标准
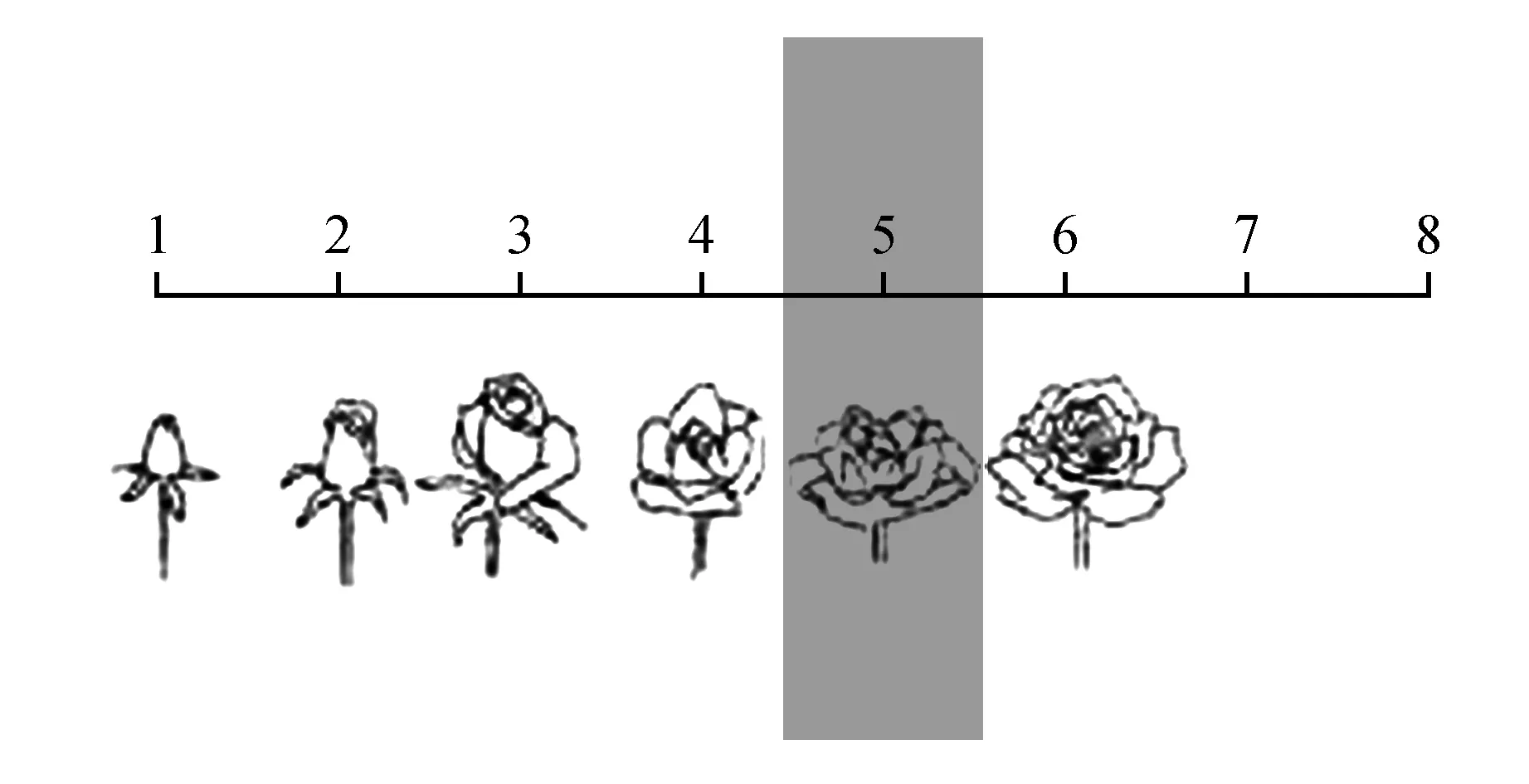
图1 测量取材示意图Fig.1 A method diagram of the measurement

图2 花朵直径测量示意图Fig.2 A schematic diagram for measuring corolla diameters

图3 花瓣长、宽测量示意图Fig.3 A schematic diagram for measuring the length and width of petals
(3)花瓣数量和瓣化情况:对花瓣数量进行直接统计,重瓣性高的子代待花瓣完全打开后进行计数,雄蕊瓣化特别明显的计入其中。参照国家林业局于2010年发布的《蔷薇属植物新品种特异性、一致性、稳定性测试指南》,采用统计与筛选的方法将花瓣数量的平均值按照单瓣花(<10)、半重瓣花(10~20)、重瓣花(20~50)及千重瓣(>50)4类进行统计,分别赋值为0、1、2和3;未瓣化和瓣化赋值为0和1。
(4)花瓣长和花瓣宽:分别选取3朵花各1枚花瓣(重瓣花选取从外至内第2~3轮的平展花瓣,单瓣花直接用该轮花瓣)进行如图3的测量,花瓣中轴线为花瓣长,与中轴垂直的花瓣最宽处为花瓣宽。
(5)萼片附属物:观测萼片的形态,统计并记录是否存在明显的附属物(将无附属物、有毛、小刺、明显刺分别赋值0、1、2和3)。
1.2.2 叶片性状的测量方法
参照国家林业局于2010年发布的《蔷薇属植物新品种特异性、一致性、稳定性测试指南》,使用游标卡尺对每个单株盛花期时花枝下第3侧枝顶叶进行测量,重复测量3枚叶片,叶长以叶基到叶尖的距离为准,叶宽以叶片最宽处为准。
1.3 数据分析
将测量和计算所得数据通过Excel 2013、SPSS 24.0等软件进行统计分析。通过平均值、标准差的计算得出变异系数(CV=标准差/平均值),用以衡量表型性状在群体内的变异水平。通过描述性统计、遗传分析以及相关性分析等得出该F1群体9个表型性状的变异特征。
2 结果与分析
2.1 月季表型性状的变异分析
利用SPSS 24.0对花朵直径、花冠面积、花瓣数量、花瓣长、花瓣宽、花瓣长宽比、叶片长、叶片宽以及叶片长宽比9个性状进行描述性统计分析(表3)。结果显示,整个群体的变异系数在7.33%~68.08%,证明群体内表型差异幅度大。花瓣数量标准差为19.29,变异系数达到68.08%,证明群体间在这个性状上的差异显著。花冠面积变异系数达到31.58%,证明群体里花冠大小差异明显;花瓣长、花瓣宽、叶长和叶宽的变异系数明显大于花瓣长宽比和叶片长宽比,证明该群体花瓣和叶片在大小上有所差异,但花瓣形态和叶型较为稳定。
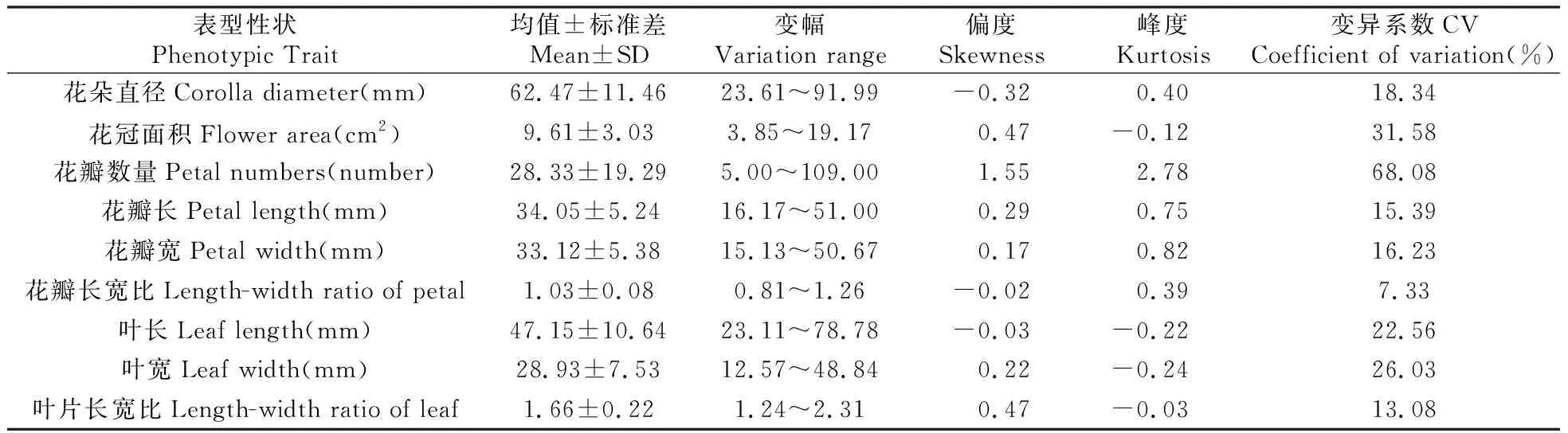
表3 F1表型性状统计
对F1代群体表型性状进行频数分布分析(图4),9个表型性状都呈现较为突出的单峰分布,接近正态分布,符合数量性状的特点,其中花朵直径、花瓣长、花瓣宽以及叶长和叶宽的偏度绝对值较接近0,证明这几个表型性状分布较为均匀;花冠面积、花瓣长宽比、叶长、叶宽以及叶片长宽比的峰度绝对值较小,证明这几个性状与正态曲线拟合性更好;花瓣数量的偏度以及峰度绝对值大于1,呈现出明显正偏性和极端性。
2.2 月季表型性状遗传分析
对F1群体9个表型性状与亲本进行比较分析(表4),F1代群体中花瓣数量和叶片长宽比的均值明显小于中亲值,其余7个性状的均值与中亲值的比值接近100%。从总体上看,杂交后代分离广泛,各个性状均出现小于低亲和大于高亲的超亲个体。其中花冠面积在双亲之间的子代比例达到92.93%;花朵直径、花瓣数量、花瓣长、花瓣宽、叶长、叶宽六个性状在双亲之间的子代约占整体80.00%,花朵直径小于低亲的个体约为大于高亲的2倍,叶宽大于高亲的个体远多于小于低亲的个体;花瓣长宽比介于双亲之间的个体只占整体10.33%,大于高亲的个体达到65.76%,呈现花瓣形状更狭长的趋势;叶片长宽比介于双亲的个体也较少,73.91%个体小于低亲,呈现杂交后代叶片形状更圆的趋势。
2.3 月季表型性状频率分析
2.3.1 雄蕊瓣化
母本‘云蒸霞蔚’和父本‘太阳城’均存在不同程度的雄蕊瓣化,在184株F1代群体中,71%子代发生雄蕊瓣化,29%子代未发生雄蕊瓣化,即瓣化:未瓣化约为3:1(图5)。由此可见,瓣化在杂交后代中的发生较为普遍,但仍存在约1/4的单株不同于亲本,未发生雄蕊的瓣化,表明在瓣化性状上,杂交后代存在较为明显的差异性。其中重瓣花约占整个群体1/2;半重瓣花约达到1/4;千重瓣花和单瓣花比例较少,各占整个群体的1/8,即四个阶段的比例为单瓣花∶半重瓣花∶重瓣花∶千重瓣花=4∶2∶1∶1(图6)。
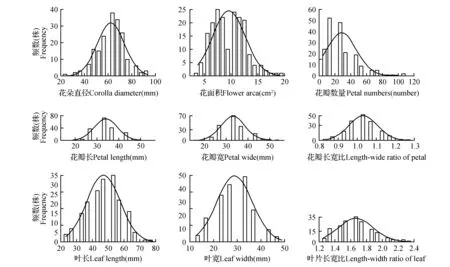
图4 F1代群体表型性状频数分布图Fig.4 Histogram of frequency distribution about the phenotypic traits of F1 population

表型性状Phenotypic trait亲本ParentsF1代群体与亲本比较Comparison among F1 population and parents(%)♀♂中亲值PMid-parents均值/中亲值X/P小于低亲Less than the low parent双亲之间Between the parents大于高亲Greater than the high parent花朵直径Corolla diameter(mm)75.6752.7664.2197.2919.5770.1110.33花冠面积Flower area(cm2)14.494.619.55100.631.6392.935.43花瓣数量Petal numbers(number)11.0053.0032.0088.5313.0476.6310.33花瓣长Petal length(mm)41.1225.6233.37102.033.8087.508.70花瓣宽Petal width(mm)41.1025.9733.5498.766.5284.788.70花瓣长宽比Length-width ratio of petal1.000.990.99103.6723.9110.3365.76叶长Leaf length(mm)60.5332.4646.50101.4110.8778.8010.33叶宽Leaf width(mm)33.7614.5624.16119.741.6375.5422.83叶片长宽比Length-width ratio of leaf1.792.232.0182.5473.9124.461.63
2.3.2 萼片附属物
母本和父本在萼片上附着有毛,184株F1代群体中,萼片附属物这一性状发生了分离,其中与亲本一样附着有毛的株数为106株,占杂交群体的一半以上;其次是萼片上无附属物,株数为35株,约占群体的1/5;另外还有15.76%萼片附着有大刺,7.61%萼片附着有小刺,4种性状分离比为有毛∶无附属物∶大刺∶小刺=15∶5∶4∶2(表5)。
表5F1代萼片附属物分布分析
Table5ThedistributionofsepalsappendagesinF1population
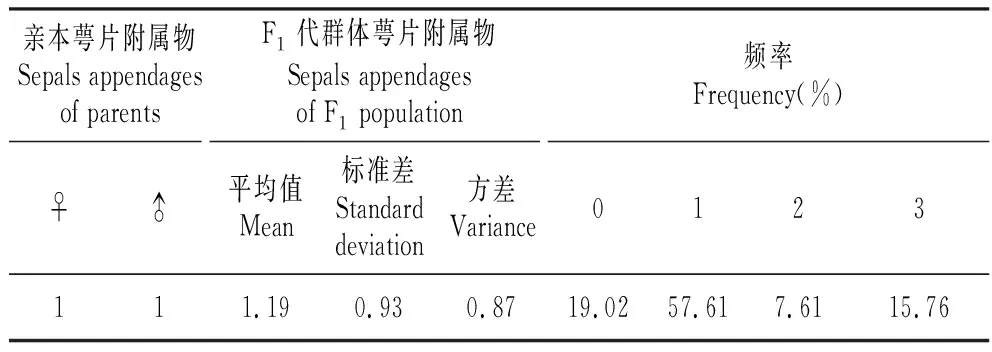
亲本萼片附属物Sepals appendages of parentsF1代群体萼片附属物Sepals appendages of F1 population频率Frequency(%)♀♂平均值Mean标准差Standard deviation方差Variance0123111.190.930.8719.0257.617.6115.76
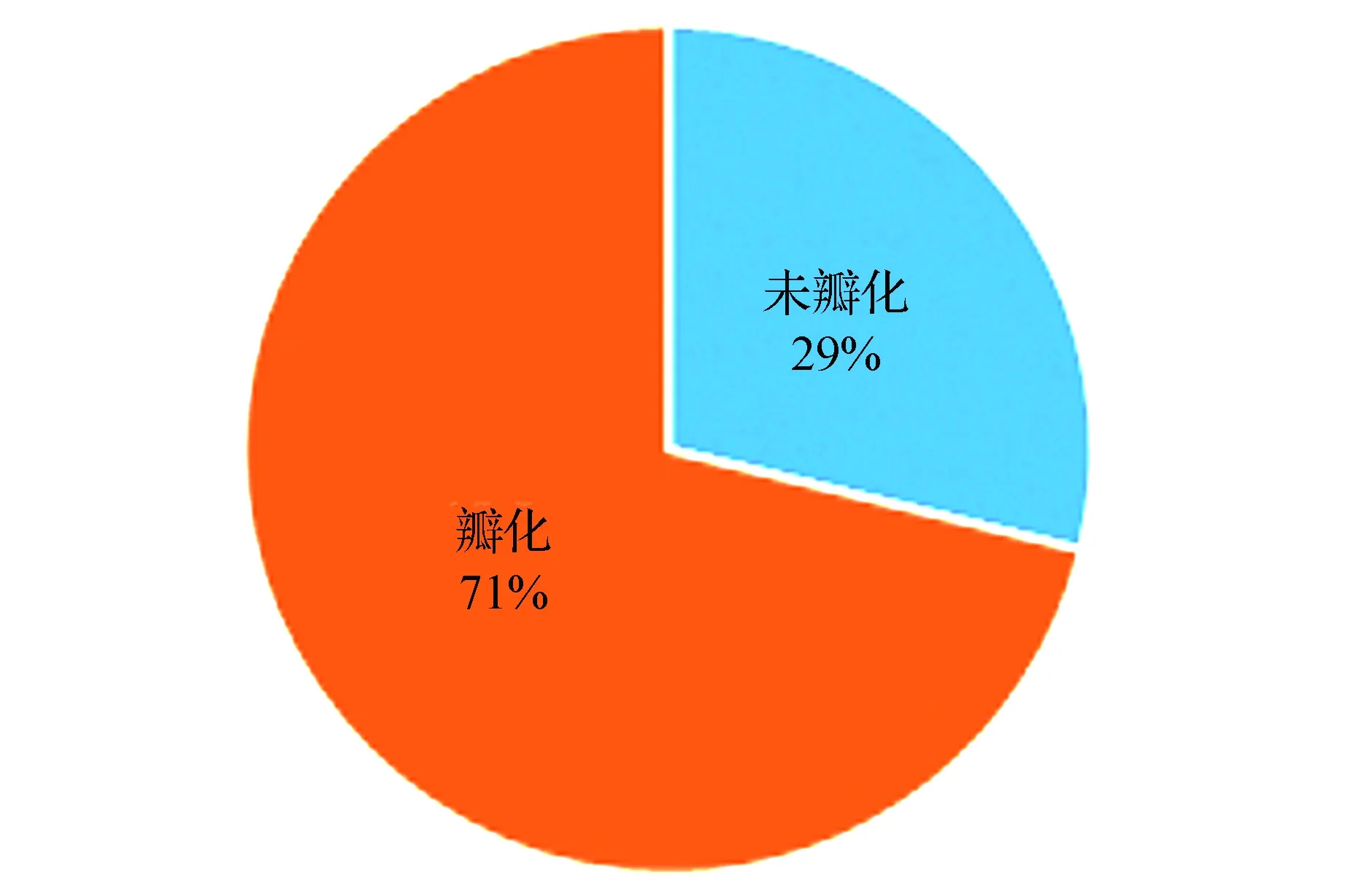
图5 F1代瓣化情况Fig.5 The petalody of F1 population

表型性状Phenotypic trait花瓣宽Petal width花瓣长宽比length-width ratio of petal花瓣数量Petal numbers花朵直径Corolla diameter花冠面积Flower area叶长Leaf length叶宽Leaf width叶片长宽比Length-width ratio of leaf花瓣长Petal length0.889**0.095-0.0420.774**0.0610.147*0.182*-0.111花瓣宽Petal width-0.363**0.0270.636**0.0350.1140.112-0.039花瓣长宽比Length-width ratio of petal-0.1290.181*0.0330.0350.113-0.143花瓣数量Petal numbers-0.0140.080-0.113-0.1090.015花朵直径Corolla diameter0.0910.1100.185*-0.166*花冠面积Flower area-0.083-0.044-0.076叶长Leaf length0.865**0.001叶宽Leaf width-0.487**
**P<0.01;*P<0.05
2.4 花部与叶部性状相关性分析
分析花部7个性状和叶片3个性状之间的相关性(表6)。花瓣数量、花冠面积与其他性状的相关性较低,花朵直径分别与花瓣长和宽呈显著正相关关系,证明花瓣长、花瓣宽影响花朵直径,但未表现出对花瓣数量以及花冠面积的影响。花瓣宽与花瓣长呈显著正相关,与花瓣长宽比呈显著负相关。叶宽与叶长呈显著正相关,与叶片长宽比呈显著负相关,与花瓣情况类似,证明叶片和花瓣的长宽比受其宽度影响更大;花部与叶部各个性状相关性不显著,只有花瓣长分别与叶长和叶宽存在弱的正相关关系以及花朵直径与叶宽和叶片长宽比存在弱相关性。
3 讨论
观赏植物的表型性状由其基因型和环境共同作用[15]。植物的大部分性状都是数量性状,掌握数量性状的遗传规律是育种工作的基础[16]。通过杂交可以使后代性状发生广泛分离[9],建立一个分离群体是开展数量性状位点的定位分析的重要方式之一。对生长于同一环境中的杂交群体的表型性状进行系统测定和分析,对于研究观赏性状的遗传规律、探索该性状表现型与基因型的关联性以及开展后续目标育种具有重要意义[17]。李宗艳等[18]通过对黄牡丹12个形态指标进行观测,发现花部形态变异较叶部大,且萼片的变异系数最大。本研究184株F1代群体中,各性状变异系数在7.33%~68.08%,证明群体内表型差异大,遗传多样性丰富。其中花瓣数量和花冠面积变异系数较高,但是花瓣形态变异较叶部小。花瓣和叶片长宽比的变异程度明显小于花瓣和叶片各自长宽,证明花瓣和叶片大小受环境影响较大,但形状较为稳定。对各性状进行遗传分析,杂交后代大部分性状介于亲本之间,但是也均出现小于低亲和大于高亲的个体,体现出杂交优势。
贺丹等[19]对尾叶紫薇(Lagerstroemiacaudata)与紫薇(L.indica) F1代群体主要表型性状的研究表明叶长、叶宽与叶面积呈极显著正相关,认为性状的相互关联是由控制该性状的QTLs相互关联造成或者由某个QTL的一因多效所引起。周俐宏等[20]对现代月季表型性状进行相关性分析并和SSR标记进行关联分析,成功的找到了与现代月季6 个表型性状相关联的SSR标记。其中叶长、叶宽、花径两两间呈极显著相关。由于月季花部表型直接影响观赏效果,本研究除了叶长、叶宽外,选取花部5个代表性状进行相关性分析,发现花朵直径分别与花瓣长和宽显著正相关,与叶宽存在弱的正相关关系。花瓣和叶片各自的长、宽和长宽比三个性状之间的相关性相似,证明长宽比受其宽度影响更大。性状间相关性的研究可以辅助提前选择[21],有利于缩短育种时间,加快育种进程。但是,本研究发现,花朵直径和花冠面积未表现出相关性。另外,花朵直径在群体中的变异系数为18.34%,花冠面积在群体中的变异系数高达31.58%,同样是描述花冠俯视面大小的两个指标,变异系数相差很大,证明花朵直径所包含的花部的信息少于花冠面积,所以呈现花朵直径的变异系数低于花冠面积的结果。由此推测本研究中所测的叶长和叶宽所代表的叶部信息有限,叶面积等其他叶部性状的测量可以使叶部信息更完善[22]。目前,由于一维指标测量起来更加容易,研究者对月季花冠大小倾向于选择一维的花朵直径、花瓣长、花瓣宽等性状作为指标;相关研究表明,回归方程法可以满足叶面积连续测定的需要,数据精度较高[23]。同样的方式也可以应用到其他植物花冠面积、叶面积的测量当中[24],建立花瓣长、花瓣宽、花朵直径、花瓣数量等单因子或多因子的花冠面积回归方程以及叶长、叶宽的叶面积回归方程,可以更准确代表花部信息,便于探索各性状遗传和相关显著性等关系。
自然界有丰富多彩的重瓣观赏植物,其起源大致包括积累起源、苞片起源、雌雄蕊起源、台阁起源、重复起源和花序起源6种方式[25]。本研究的F1群体表现出了明显的雄蕊瓣化现象,验证了月季重瓣性来源于雄蕊瓣化。Serrato[26]对万寿菊(Tageteserecta)瓣性研究也表明,单瓣花的后代是单瓣或复瓣;重瓣花的后代中单瓣、复瓣和重瓣均有分布。黄秀强[27]等用单瓣美洲黄莲(Nelumbonucifera)与单瓣莲杂交,其种间杂种仍为单瓣,与重瓣莲品种杂交,其种间杂种为半重瓣或重瓣,据此认为莲属花型并非简单的孟德尔遗传,而是由多基因控制。于超[1]检测到了月季杂交群体7个与花瓣数量相关联的QTLs,证明花瓣数量这一性状为数量性状,并可能存在主效基因控制。上述情况也在本研究中得到验证,该群体在花瓣数量这一性状上分离程度高,呈正态分布,证明月季的重瓣性表现为由多基因控制的数量性状,通过多代杂交选育,后代往往可能出现超过亲代花瓣数量的品种,此发现对于指导月季重瓣花育种和探索花发育的遗传调控因子具有参考价值。
猜你喜欢
金沙江文艺(2022年3期)2022-03-25 02:06:34
新疆农业科学(2020年1期)2020-02-14 03:22:46
亚热带植物科学(2019年2期)2019-07-22 13:16:12
第二课堂(小学版)(2019年4期)2019-05-13 21:54:18
江苏农业学报(2018年5期)2018-09-10 22:36:44
大灰狼(2018年3期)2018-06-11 15:28:50
江苏农业科学(2016年10期)2017-02-05 14:45:09
中国三峡(2016年5期)2017-01-15 13:58:53
家家乐·上半月(2013年6期)2013-04-29 00:44:03
