
2012—2017年海州湾潮间带大型底栖动物群落结构及变化
2019-01-19 01:51季相星于爱琛
中国环境监测 2018年6期
季相星,李 军,姜 毅,于爱琛
连云港市环境监测中心站,江苏 连云港 222001
海州湾位于中国黄海中部,为典型的开敞式海湾。近年来,许多学者已对中国大部分海域潮间带底栖生物开展了较系统的研究[1-3],但对海州湾潮间带的研究却较少[4-6],特别是跨年度的研究更是鲜有报道。目前人类在海州湾潮间带广泛开展了养殖活动,这势必对其大型底栖动物群落结构造成影响。笔者对2012—2017年海州湾潮间带大型底栖动物的群落结构及其年度变化进行了研究,分析了人类养殖活动对其群落结构的影响,为该海域生态系统的深入研究打下基础,为科学养殖和生物多样性保护提供参考。
1 实验部分
1.1 采样点设置
研究断面设置于原海州湾旅游度假区潮间带,位于江苏省连云港海头镇,地理坐标为119°12′30″E、34°55′57″N,按高潮带、中潮带和低潮带依次设置3个采样点(图1)。于2012—2016年每年的9—10月(秋季)进行一次采样;为研究其周年变化,另于2017年2、5、8月各进行一次采样,共计采样8次。
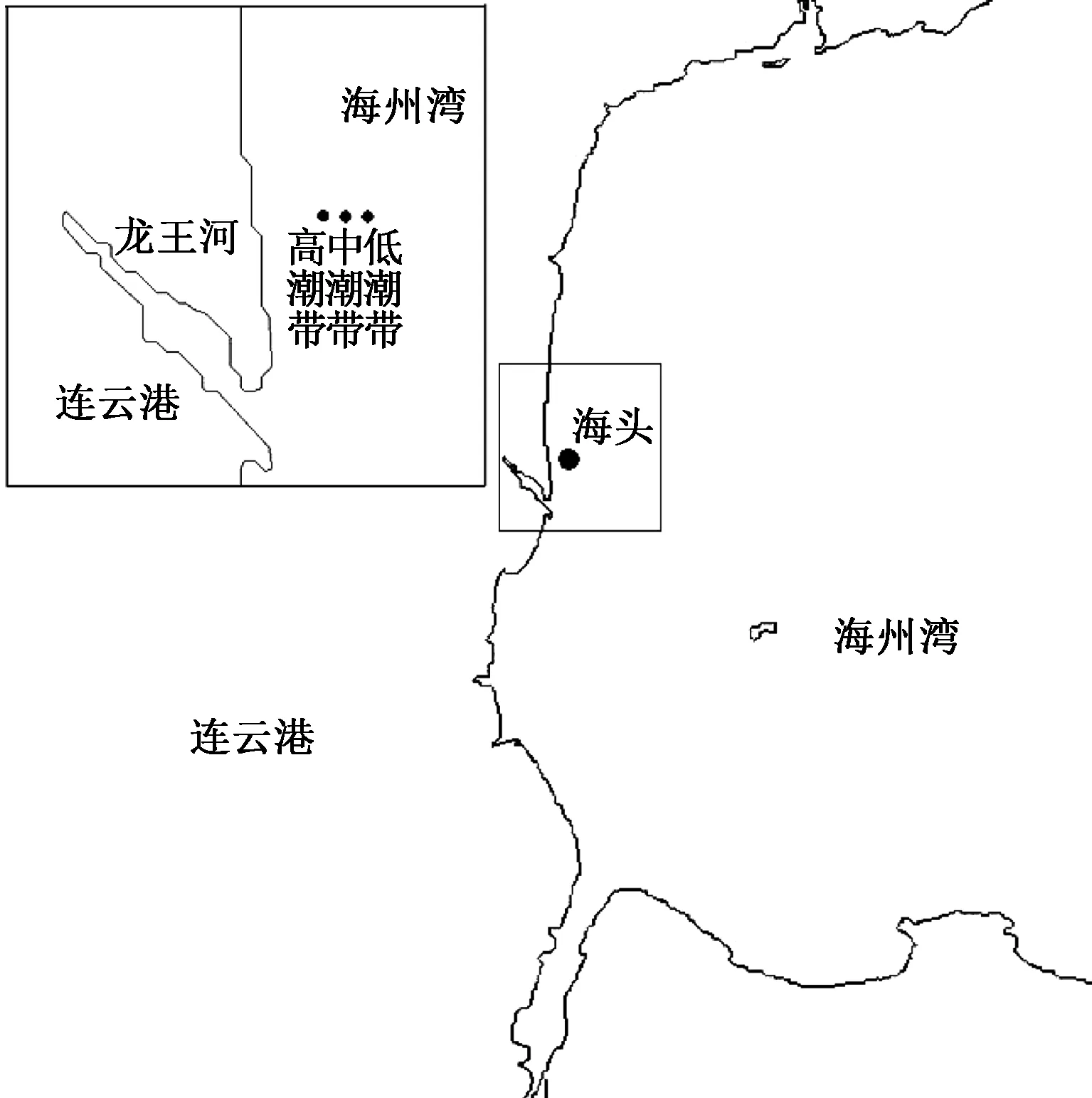
图1 海州湾潮间带大型底栖动物调查采样点Fig.1 Location of sampling stations of macrobenthos in Haizhou Bay
1.2 样品采集和处理
生物样品的采集和处理按照《海洋监测规范》[7]进行。使用GPS精确定位,在退大潮时到采样点处采样。采用25 cm×25 cm取样框随机取样,每次取3个平行样。取样后当场用0.5 mm的网筛进行筛选,将所留物加等体积10%甲醛溶液固定后带回实验室,置于常温保存。实验室使用0.5 mm网筛分选样品,在体视显微镜下进行样品种类鉴定及计数。挑选出的所有大型底栖动物标本用感量为0.000 1 g的电子分析天平称生物量湿重。
1.3 数据分析
1.3.1 优势种的确定
采用相对重要性指数(Index of Relative Importance,IRI)[8]确定优势种。取研究中各物种的平均丰度及平均生物量计算其IRI值,同时分
别计算每次调查的IRI值,计算公式如下:
IRI=(W+N)×F
(1)
式中:W为每种生物量占总生物量的百分比,N为每种丰度占总丰度的百分比,F为该种出现的频率。
1.3.2 多样性指数的计算
采用Shannon-Wiener多样性指数(H′)[9]、Margalef丰富度指数(d)[10]以及Pielou’s均匀度指数(J′)[11]进行多样性分析。根据《近岸海域环境监测规范》[12]对H′指数的划分进行评价。
公式如下:
(2)
d=(S-1)/log2N
(3)
J′=H′/log2S
(4)
式中:Pi为第i种的个体数与总个体数的比值,N为总个体数,S为总种数。
1.3.3 聚类分析
群落结构的分析主要利用PRIMER 5.0软件包进行。采用组平均法CLUSTER聚类划分海州湾潮间带大型底栖动物群落的种类组成,并使用SIMPER分析了解表征群落的物种(特征种)[13]。
2 结果
2.1 大型底栖动物种类组成
2012—2017年在海州湾潮间带共发现120种大型底栖动物,包括44种环节动物多毛类,21种软体动物,46种甲壳动物,4种刺胞动物,2种腕足动物,纽形动物、扁形动物及半索动物各1种。
图2显示了2012—2017年海州湾潮间带大型底栖动物总种数及类群组成。
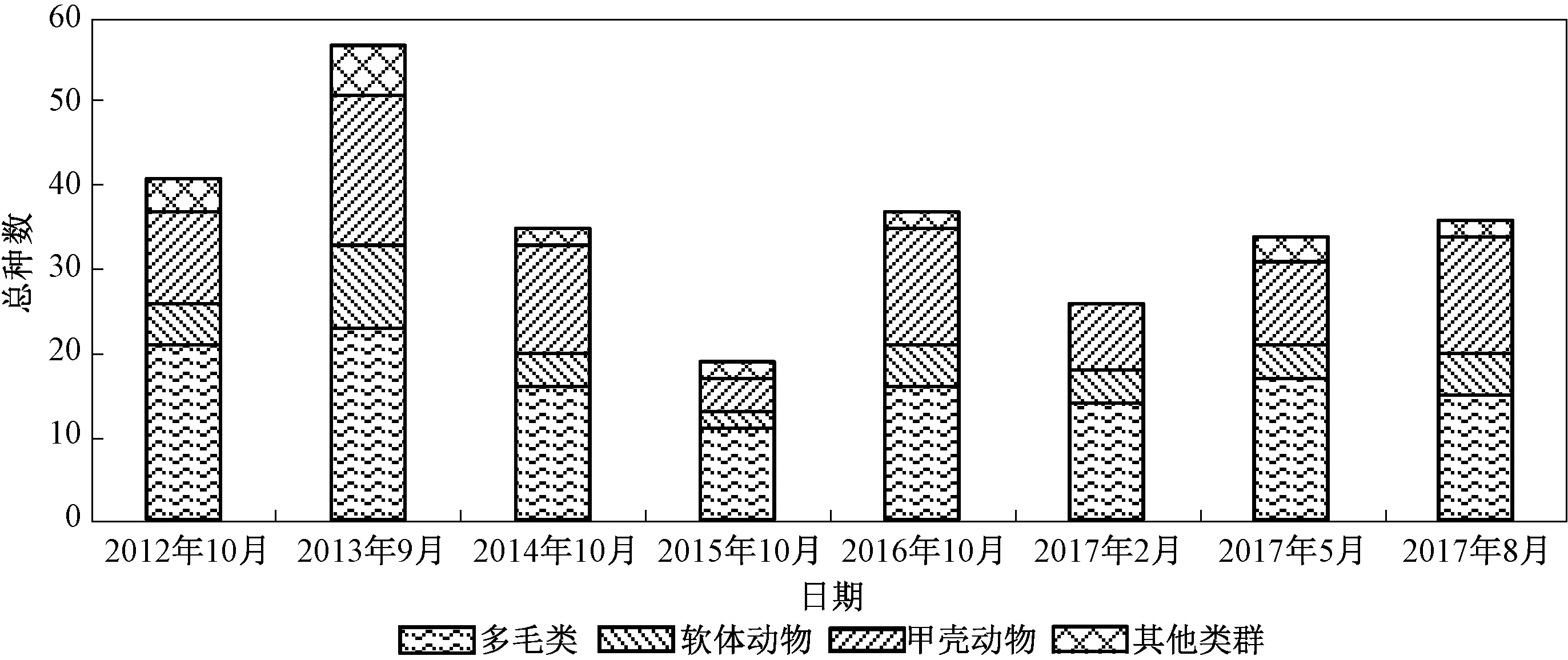
图2 2012—2017年海州湾潮间带大型底栖动物总种数及类群组成Fig.2 Species number and composition of macrobenthos in the intertidal zone of Haizhou Bay from 2012 to 2017
如图2所示,2013年调查发现物种最多(57种);2015年调查发现物种最少(19种);2017年2月调查发现物种略高于2015年(26种);其余年份调查发现物种数量相近。
多毛类和甲壳动物是海州湾潮间带大型底栖动物最主要的类群。如图2所示,在每次调查中多毛类均为第一大类群,甲壳动物为第二大类群;而将历年数据综合统计后却发现甲壳动物为第一大类群,多毛类为第二大类群。这说明每次调查发现的多毛类重复物种较多,而每次调查的甲壳动物物种的变化较大。
2.2 大型底栖动物丰度变化
2012—2017年海州湾潮间带大型底栖动物丰度范围为506.7~17 864.0 ind./m2,总平均丰度为3 495.9 ind./m2。其中多毛类总平均丰度为413.7 ind./m2,占总平均丰度的11.83%;软体动物为2 453.3 ind./m2,占70.18%,为丰度第一大类群;甲壳动物为603.4 ind./m2,占17.26%;其他类群共25.2 ind./m2,占0.72%。
图3显示了2012—2017年海州湾潮间带大型底栖动物丰度变化。
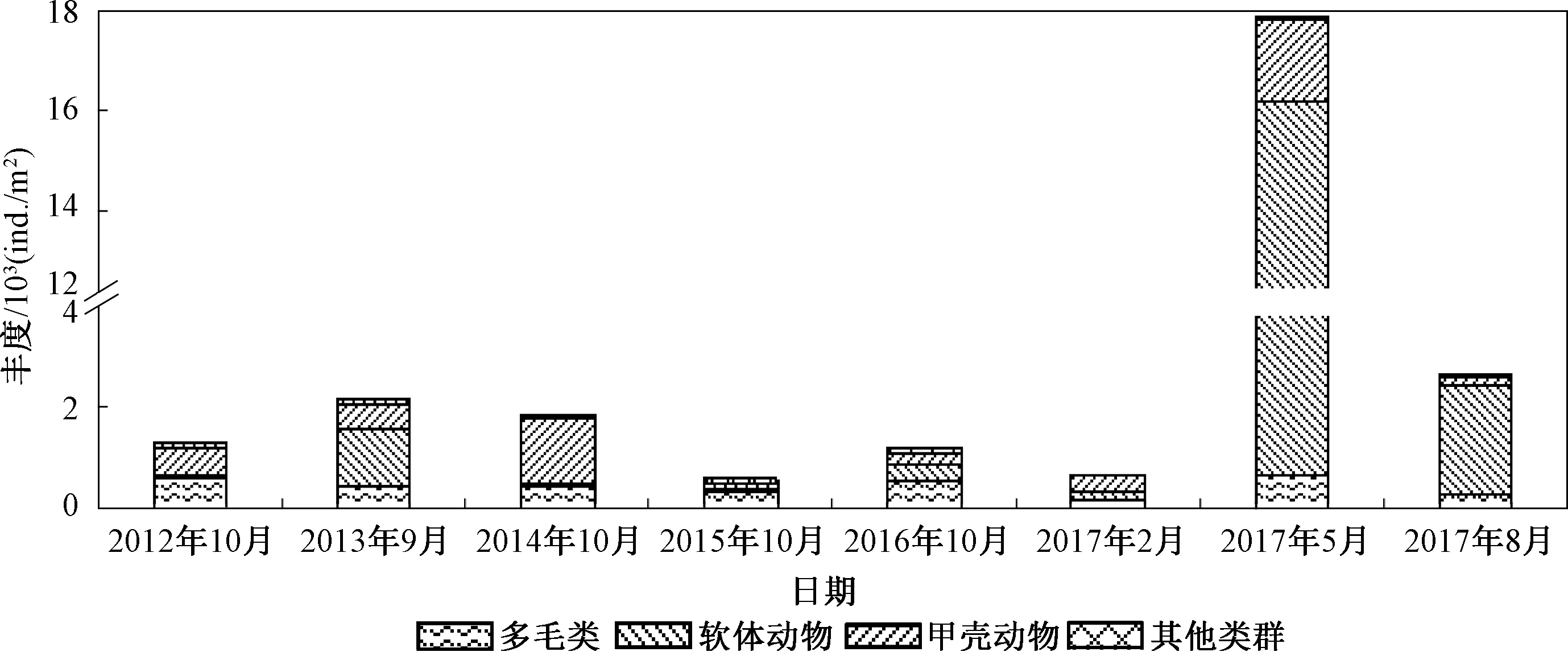
图3 2012—2017年海州湾潮间带大型底栖动物丰度Fig.3 Abundance of macrobenthos in the intertidal zone of Haizhou Bay from 2012 to 2017
如图3所示,同物种数一样,2015年的丰度最低(506.7 ind./m2);2017年2月的丰度仅高于2015年(650.7 ind./m2)。
近5年大型底栖动物的丰度变化较剧烈,特别是2017年5月的丰度远高于其他调查时段。此次调查发现了大量的光滑河蓝蛤(Potamocorbulalaevis),其丰度为15 544 ind./m2,占该次调查丰度的87.01%。根据现场调查,该潮间带如今是光滑河蓝蛤的养殖区,5月正是其快速生长的季节。同时,该次调查甲壳动物中国周眼钩虾(Perioculodesmeridichinensis)的丰度也较高(1 194.7 ind./m2)。
除2017年5月外,2013、2014年以及2017年8月调查丰度也较高。2017年8月同5月一样,其高丰度也是由光滑河蓝蛤引起的(2 141.3 ind./m2),其丰度占该次调查总丰度的81.44%。2013年的高丰度主要是由光滑狭口螺(Stenothyraglabar)引起的(1 036.4 ind./m2),其丰度占总丰度的48.91%。2014年中国周眼钩虾的丰度为874.7 ind./m2,对总丰度的贡献率达到了48.38%。
2.3 大型底栖动物生物量变化
2012—2017年海州湾潮间带大型底栖动物生物量范围为1.58~489.17 g/m2,总平均生物量为197.26 g/m2。其中多毛类总平均生物量为15.34 g/m2,占总平均生物量的7.78%;软体动物为161.20 g/m2,占81.72%,为生物量第一大类群;甲壳动物为14.24 g/m2,占7.22%;其他类群共6.47 g/m2,占3.28%。
图4显示了2012—2017年海州湾潮间带大型底栖动物生物量变化。
同丰度变化一样,2015年生物量最低(1.58 g/m2),2017年5月的生物量最高(489.17 g/m2)。如上所述,2017年5月调查发现了大量的光滑河蓝蛤,这也导致其在该次研究中的生物量最高。同时,2016年10月—2017年8月调查生物量都较高,且均是由光滑河蓝蛤所引起的。
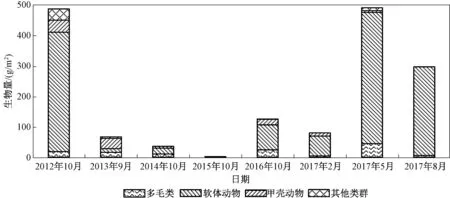
图4 2012—2017年海州湾潮间带大型底栖动物生物量Fig.4 Biomass of macrobenthos in the intertidal zone of Haizhou Bay from 2012 to 2017
对比图3可见,2012年的丰度并不高,但其生物量较高(485.22 g/m2),仅次于2017年5月,该次调查在中潮带发现了四角蛤蜊(Mactraveneriformis),为典型的低丰度高生物量物种,其在中潮带生物量为1 127.11 g/m2,这是该次调查丰度不高而生物量高的主要原因。
2.4 海州湾潮间带大型底栖动物优势种
表1显示了该次研究中IRI值居前十位的种。

表1 IRI值居前十位的种Table 1 The top 10 species of IRI
如表1所示,光滑河蓝蛤的IRI最高,且远高于其他物种。根据研究统计结果,光滑河蓝蛤在2015年之前并不在IRI前十种之列,而在2015年开始出现,且IRI远高于其他物种。
光滑河蓝蛤、智利巢沙蚕、光滑狭口螺、长吻沙蚕和长叶索沙蚕属高丰度高生物量物种;中阿曼吉虫、中国周眼钩虾、三叶针尾涟虫和多鳃齿吻沙蚕属典型的高丰度低生物量物种;四角蛤蜊为典型的低丰度高生物量物种。
除四角蛤蜊仅出现于2012年的调查中外,其余物种均多次出现于IRI前十种之列,它们对海州湾潮间带大型底栖动物群落起着较重要的作用。四角蛤蜊虽仅出现于2012年的调查中,但其IRI较高,其消失引起了群落组成的变化,对海州湾潮间带大型底栖动物群落结果产生了较大的影响。
2.5 生物多样性指数
图5显示了2012—2017年生物多样性指数变化趋势。
如图5所示,H′指数、d指数及J′指数均呈下降、上升再下降的趋势,而整体均呈现出下降的趋势。根据H′指数评价,可将2012、2013、2016年海州湾潮间带环境评价为“优良”,2014、2015年和2017年2月评价为“一般”,而2017年5、8月评价为“差”。
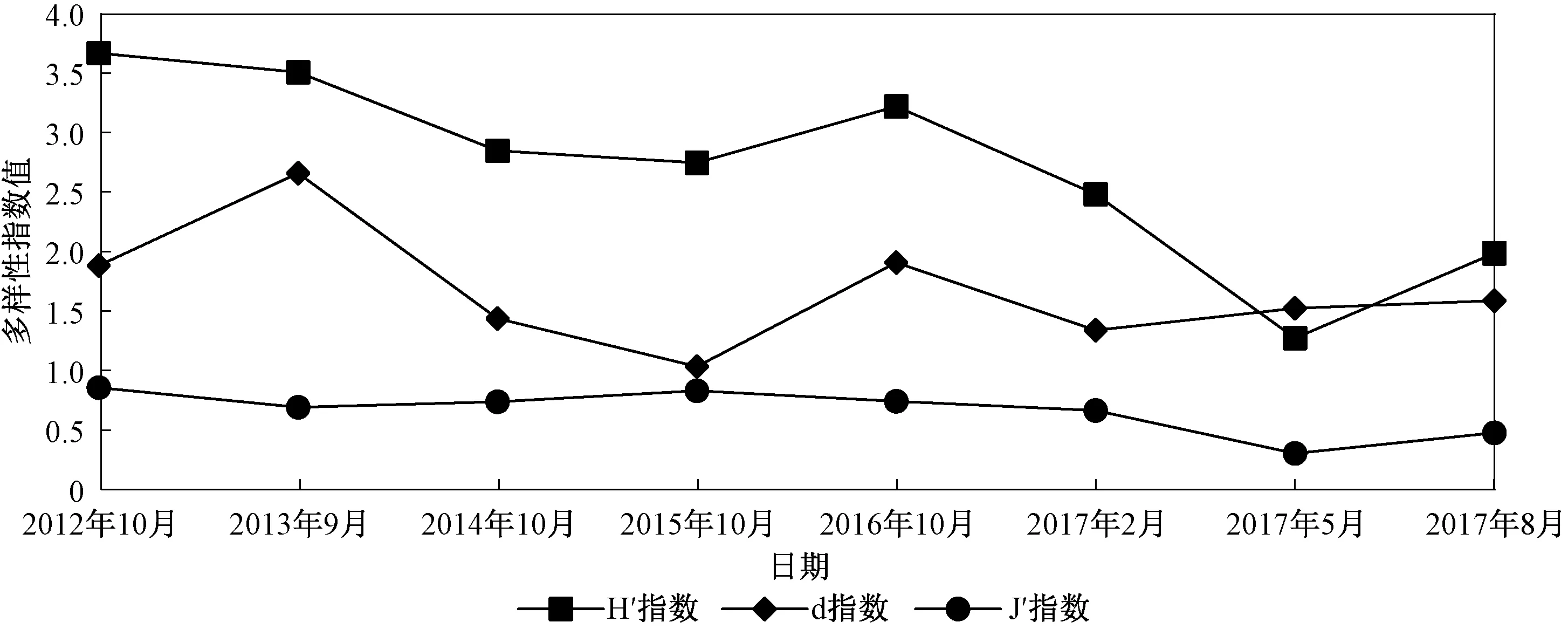
图5 2012—2017年海州湾潮间带大型底栖动物多样性指数变化趋势Fig.5 Diversity index of macrobenthos in the intertidal zone of Haizhou Bay from 2012 to 2017
2.6 聚类分析
图6显示了2012—2017年海州湾潮间带大型底栖动物种类组成聚类分析结果,同时使用SIMPER分析各组的特征种。

图6 2012—2017年海州湾潮间带大型底栖动物种类组成聚类分析Fig.6 The results of cluster analysis of macrobenthos in the intertidal zone of Haizhou Bay from 2012 to 2017
如图6所示,在40%的相似性上,可将8次调查分为4组,2012、2013年分别单独为一组,无特征种。2014年和2017年5月为一组,该组平均相似性仅为14.83%,特征种为中国周眼钩虾(贡献率为59.96%)、三叶针尾涟虫(贡献率为13.16%)和中阿曼吉虫(贡献率为8.41%),累计贡献率为81.54%。主要是因中国周眼钩虾的贡献而将其划为同一组。值得注意的是,对2017年5月群落结构影响极大的光滑河蓝蛤在此并未体现出来。2015、2016年和2017年2、8月为一组,该组平均相似性为29.08%,特征种为光滑河蓝蛤(贡献率为22.80%)、中国周眼钩虾(贡献率为17.39%)、中阿曼吉虫(贡献率为16.32%)、长吻沙蚕(贡献率为13.59%)、智利巢沙蚕(贡献率为6.02%)及三叶针尾涟虫(贡献率为5.16%),累计贡献率为81.28%,这些特征种均出现于IRI前十位之列。
3 讨论
3.1 人类养殖活动对大型底栖动物群落的影响
结合走访调查,并参考高爱根等[4]的研究结果,可大体总结出人类养殖活动在海州湾潮间带的变迁历史。在2012年之前,该潮间带是四角蛤蜊的养殖区,随后,四角蛤蜊的养殖逐渐减少,从2013年至今的7次调查均未再发现四角蛤蜊;2013—2014年该潮间带处于养殖空档期,大型底栖动物在此自由生存与竞争,从2013、2014年第一优势种分别为光滑狭口螺和中国周眼钩虾可看出这一现象的存在;2015年至今,渔民大量养殖光滑河蓝蛤,其成为绝对优势种;同时,由于2015年为恢复养殖的第一年,渔民对潮间带进行了“翻耕”,导致该年度发现的物种及其丰度、生物量均远低于其他调查年份。
可见人类养殖活动对海州湾潮间带大型底栖动物群落结构产生了及其重要的影响。当养殖四角蛤蜊时,四角蛤蜊便是第一优势种;当停止养殖时,各物种自由竞争,优势种随竞争而更迭;当为养殖做准备而“翻耕”时,群落的种类数、丰度、生物量都远低于其他时间;当养殖光滑河蓝蛤时,其在各季节都成为绝对优势种。同时从H′指数的变化可见,光滑河蓝蛤的大量养殖也导致了大型底栖动物多样性的降低。
3.2 海州湾潮间带光滑河蓝蛤的生长周期
根据魏利平等[14]和孙晋廷[15]的研究,光滑河蓝蛤的繁殖盛期在每年9月中旬至10月中旬,而刘吉明等[16]的研究则认为光滑河蓝蛤每年有2次产卵高峰期,分别为5月初和9月底。根据研究调查结果进行分析,海州湾潮间带光滑河蓝蛤的繁殖盛期在9—10月。
表2显示了2016—2017年调查光滑河蓝蛤的丰度、生物量及个体重量情况。
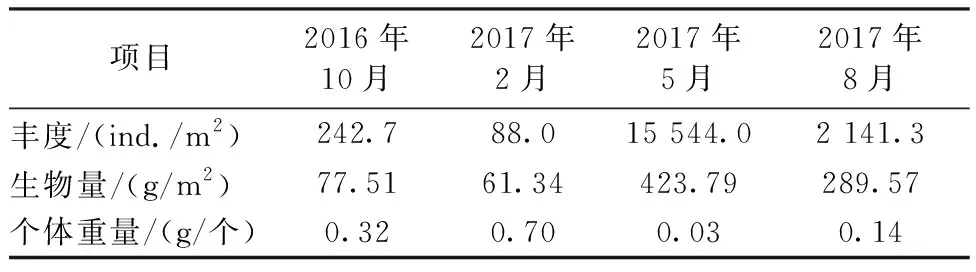
表2 2016—2017年调查光滑河蓝蛤的丰度和生物量Table 2 The abundance and biomass of P. laevis from 2016 to 2017
结合表2和当地的养殖活动,可得出海州湾潮间带光滑河蓝蛤的生长周期。渔民在当年8月进行采捕,但会留下一定数量亲贝繁殖后代;9—10月光滑河蓝蛤大量繁殖;随着温度降低,丰度也随之降低,但个体重量在增加,这时上一年繁殖的光滑河蓝蛤个体还小,大型底栖动物调查还无法采集到;随着第二年温度的升高,大量的光滑河蓝蛤幼体快速生长,这便是2017年5月其丰度极高的原因。
4 结论
此次调查在海州湾潮间带共发现大型底栖动物120种,其总平均丰度为3 495.9 ind./m2,总平均生物量为197.26 g/m2,近年丰度及生物量的波动均较大。光滑河蓝蛤为第一优势种,其大量养殖对大型底栖动物的群落结构造成了重要的影响,同时也导致了多样性的降低。
猜你喜欢
中国水产(2021年1期)2021-03-01
海洋信息技术与应用(2020年4期)2021-01-18
今日农业(2020年19期)2020-12-14
军事运筹与系统工程(2020年2期)2020-11-16
海洋通报(2020年3期)2020-10-20
军事运筹与系统工程(2018年3期)2018-03-26
中国科技信息(2015年2期)2015-11-16
中国科技信息(2015年18期)2015-11-10
植物营养与肥料学报(2014年1期)2014-03-11
中国烟草学报(2012年3期)2012-04-10
