
低纬高原梯田两系杂交稻制种生态适应性分析
2019-01-18 02:50陈忆昆董阳均刘晓利辜琼瑶李华慧卢义宣罗君尧奎丽梅
西南农业学报 2018年12期
涂 建,陈忆昆,董阳均,刘晓利,黄 平,辜琼瑶,李华慧,卢义宣,杨 久, 张 云,张 笑,罗君尧,奎丽梅*
(1.云南省农业科学院粮食作物研究所,云南 昆明 650205;2.云南金瑞种业有限公司,云南 昆明 650317;3.水富县种子管理站,云南 水富 657800)
【研究意义】水稻两系法育种是继三系法育种之后的一次技术革新[1-2],与三系法育种相比较,其优点在于简化了育种程序,核不育系可一系两用,不需要保持系,突破了三系法杂交水稻优势利用的恢保关系限制,而且核不育系的遗传简单,获得更多的强优势组合几率更大。此外两系法育种可选育出具有正常细胞质的核不育系,容易实现杂交水稻细胞质多样化,消除当前三系杂交稻存在的野败细胞质单一的潜在威胁,可解除野败胞质对产量的负面影响,对稻米品质、产量及抗性的改良等方面表现出明显优势。经过30余年的攻关,两系杂交水稻育种取得了重大进展,截止2010年年底,全国育成两系杂交稻组合427个通过国家或省级品种审定,两系杂交水稻在全国16个省推广种植,应用面积多次位居第一,目前两系杂交水稻推广面积占全国杂交水稻总面积的20 %左右,在水稻生产和保障粮食安全中发挥了重要作用[3]。在两系杂交水稻的大面积推广中,两系杂交稻制种技术作为其产业化的一项集成转化技术,即基于三系又不等同于三系,与三系制种相比较,两系法制种由于核不育系育性在生态环境改变的情况下因光温条件不同其育性易产生波动,不育系育性恢复自交结实,在制种的安全性上存在很大风险[4-7],因此,两系法杂交稻种子的安全生产问题已成为制约两系杂交水稻发展的瓶颈。【前人研究进展】国内有对两系杂交稻安全制种基地和时段选择方面的研究和报道[8-12],但有关研究大部分局限于某些地域,与低纬高原地区两系杂交稻安全制种研究的相关报道较少。【本研究切入点】通过对不同籼型和粳型两用核不育系在低纬高原梯田不同海拔生态条件下的育性表达规律研究,分析两用核不育系品种、海拔、播期与不育率的关系,并进行了不育系育性稳定性分析。【拟解决的关键问题】为明确低纬高原梯田两系杂交稻适宜的制种区域和播种期,为两系杂交稻在低纬高原地区安全制种生产提供有价值的数据支持。
1 材料与方法
1.1 试验材料
籼型核不育系:云软201S、2301S、蜀光357S、蜀光612S;粳型核不育系:N5088S、N95076S、云粳202S。
1.2 方法
1.2.1 试验点选择 试验点选在云南省水富县向家坝镇的上坝(海拔300 m)、香泉(海拔500 m)、永安(海拔700 m)3个农业社,地形属山区梯形田块。随着梯形田块的层叠上升,海拔逐渐升高。
1.2.2 试验设计 2013-2016年,在3个试验点共设8个播期,每期间隔10 d,自3月25日至6月3日止;全部材料统一在300 m海拔试点处播种育秧,4~5叶时同期材料同步移栽到3个试验点的大田,每个试验点每期每个材料栽插50苗,单苗移栽,栽插规格株行距为16.7 cm×20.0 cm。进入始穗期每2 d套1次隔离袋,套袋分布于整个群体的主穗和分蘖穗之中,每个试验点每期材料套袋数量不低于50个。成熟后,采集套袋穗考种,分析各核不育系在在不同海拔下的自交结实率,并作同一海拔不同材料、不同播期间的横向比较以及同一播期不同海拔间的纵向比较。本试验所涉方差、相关、回归等统计分析,均使用SPSS 12.0软件。
2 结果与分析
2.1 核不育系在不同生态条件下的育性表达
2.1.1 不同生态条件下分期播种的育性转换规律分析 7个核不育系在不同生态条件下经过4年的分期播种试验,各核不育系的自交不育率统计如表1所示。同一品种在同一水平条件下育性转换规律具有自身特性,当环境因素改变,其育性转换表现出一定规律性,海拔上升,不育率在下降,而播期推迟,不育率变化因不育系品种不同出现差异。
续表1 Continued table 1

不育系Varieties of rice PTGMS海拔(m)Altitude播期(月/日,%)Sowing dates(Month/Day)平均自交不育率(%)The average rate of self-crossing sterility3/254/44/144/245/45/145/246/3平均值(%)Average50099.35 99.71 99.21 99.35 99.15 97.59 97.49 92.35 99.76 70088.36 93.98 97.39 97.35 96.72 92.58 95.32 99.95 95.21 平均值Average94.93 96.02 98.58 98.57 98.49 96.60 96.98 93.78 96.74 N5088S30099.96 99.93 99.85 99.89 99.82 97.10 97.31 89.04 97.86 50099.90 99.89 99.62 99.43 98.40 97.71 82.39 93.71 96.38 70099.24 98.81 98.47 97.92 98.57 94.84 87.22 98.19 96.66 平均值Average99.70 99.54 99.31 99.08 98.93 96.55 88.97 93.65 96.97 N95076S30099.87 99.96 99.92 99.89 99.70 99.47 98.26 96.60 99.21 50099.91 99.71 99.74 99.70 98.55 96.83 92.78 89.17 97.05 70099.28 99.35 99.48 98.75 97.77 92.92 83.66 75.06 93.28 平均值Average99.69 99.67 99.71 99.45 98.67 96.41 91.56 86.94 96.51 云粳202S30099.94 99.99 99.99 100 99.98 99.87 99.26 99.72 99.84 50099.92 99.83 99.89 99.78 99.54 98.86 97.38 94.68 98.73 70099.86 99.61 99.69 99.11 99.09 97.73 94.55 90.79 97.55 平均值Average99.90 99.81 99.86 99.63 99.53 98.82 97.06 95.06 98.71
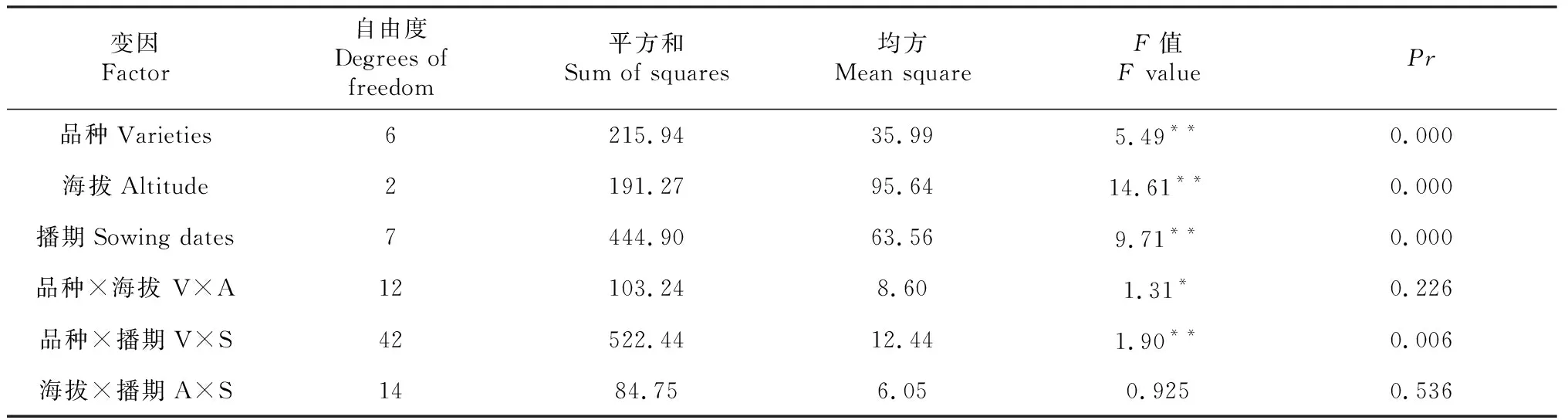
表2 2013-2016年不同生态下不育率方差分析(F值)
注:*和**分别表示在(0,05 Note:* and ** indicated significant difference at 0.05 or 0.01 level, respectively. The same as below. 2.1.2 所有核不育系的育性转化规律具有时空性 通过方差分析(表2)表明,各品种套袋自交不育率因育性基因表达不一,在品种间、海拔间、播期间均表现出极显著差异,核不育系自交不育率受多种因素的影响。在品种间与海拔间互作及海拔间与播期间互作差异不显著,说明不同的核不育系在海拔间、播期间的自交不育率表现是相一致的,核不育系的育性表达具有严格的空间性,而品种间与播期间差异达极显著,说明核不育系自交不育率受品种因素和播期的主导,其育性表达具有严格的时间限制。因此不同材料育性转换除受品种自身特性主导外,不同的环境因子均左右着育性的变化。 2.1.3 海拔升高,不育率降低,播期推迟,不育率受品种因素影响出现差异 以3月25日作为基点单位0,单位为天,对核不育系自交不育率作回归分析见表3,结果表明各参试材料不育率与海拔均呈负相关,其中除了蜀光612S、N5088S不显著相关外,其余均达显著负相关。除了蜀光612S、N5088S的不育率受海拔影响较小,各不育系的不育率受海拔影响较大,随着海拔的升高,不育率明显降低;在不育率与播期间,云软201S、2301S不育率与播期间呈正相关关系,但相关不显著;其余不育率与播期间呈负相关关系,除了蜀光612S外均达显著性关系,云软201S、2301S、蜀光612S不育率受播期影响较小,其余核不育系随着播期的推迟,不育率明显下降,云软201S、2301S随着播期推迟,不育率呈上升态势。 表3 不育率与不同因子的相关性分析 表4 不育率与海拔、播期之间相关性分析 注:Y,不育率;X1,海拔;X2,播期。 Notes:Ymeant percentage of sterility;X1meant altitude;X2meant sowing dates. 以300 m海拔作为基点单位0,海拔每上升100 m为1个单位,和以3月25日作为基点单位0,每推迟1个播期为10 d,分析海拔、播期二因素与不育率之间的回归关系。如表4所示:海拔每升高100 m,云软201S不育率下降0.85个百分点,2301S不育率下降0.08个百分点,蜀光612S不育率下降0.45个百分点,不育系云软201S、2301S、蜀光612S不育率受播期因素影响不大,而与海拔因素相关,除蜀光612S相关不显著外,云软201S、2301S达显著相关;N5088S不育率与播期因素显著相关,而受海拔因素影响不大,每推迟1个播期,不育率下降0.12个百分点;不育系蜀光357S、N95076S、云粳202S不育率均受海拔、播期的影响达极显著相关,当海拔每上升100 m,其不育率分别下降0.72个百分点、1.48个百分点和0.57个百分点,当每推迟1个播期,其不育率分别下降0.04个百分点、0.17个百分点和0.06个百分点;随着海拔的上升,除N5088S不育率不受影响外,其余核不育系不育率均在下降,其中不育率受海拔影响最大为N95076S,当海拔每上升100 m,其不育率下降1.48个百分点,影响较小的是2301S为0.08个百分点。而随着播期的推迟,蜀光357S不育率每推迟1个播期,其不育率下降0.04个百分点,N95076S下降0.17个百分点,云粳202S下降0.06个百分点,其余核不育系受播期影响不大。 选择安全生态区域制种是受核不育系自身育性表达规律所决定。在两系制种生产过程中,当核不育系不育率低于99.5 %以下时,所生产的杂交种种子纯度将承担较大风险,根据不同生态条件下分期播种的不育率如表1可知,云软201S适宜制种区域为海拔300 m,播期为4月4日之后;2301S适宜制种区域为海拔500 m以下,播期为3月25日至6月3日,当海拔上升到700 m时,可以选择到适宜制种的播期时段;蜀光357S适宜制种区域控制在海拔300 m较为安全;蜀光612S在供试点没有适宜的制种区域;N5088S适宜制种区域应控制在海拔500 m以下,海拔升高播期相应提前,最迟不超过5月4日;N5088S适宜制种区域为海拔500 m以下,播期为5月4日以前,当海拔升高,播期相应提前,当海拔升高到500 m,播期应提前到4月14日之前;N95076S不育率在海拔间差异变幅较大,其适宜制种区域为海拔500 m以下,播期为5月4日以前,海拔升高,播期提前,海拔升高到500 m时,播期提前到4月24日以前;云粳202S在供试区域均有适宜的安全播期,海拔300 m全程播种均为安全播期,当海拔上升到500 m时,安全播期提前到5月4日以前,当海拔上升到700 m时,安全播期提前到4月14日。 自1973年石明松发现两用核不育系农垦58S以来[13]。人们对核不育系的研究从未间断,目前一致认为,核不育系受自身遗传因素的影响,每个核不育系在不同年份和不同地点的不育期长短也不一致,由于核不育基因在不同遗传背景下受到表达条件的改变,从而决定着各不育系的利用价值及其适应地区。而为了两系制种需要,一般核不育系在制种生态区要求有30 d以上的不育期[14-15],,抽穗后的育性表现为稳定的不育。本试验选择在垂直距离10 km内进行,光长对核不育系的影响可以视为在同一纬度下而不受影响,温度对其影响来源于海拔差异。随着海拔的升高,温度在下降,当温度低于核不育系育性转换临界起点温度时,核不育系将从不育转为可育。目前,生产上所用的大部分核不育系的临界起点温度为21~23 ℃,根据气象学资料,海拔每上升100 m,温度下降0.6 ℃,对应2013-2016年当地温度记录分析,在海拔300 m试验点,当育性转换敏感期进入8月下旬,育性将引起波动;在海拔500 m试验点,当育性转换敏感期越过8月中旬,育性将引起波动;在海拔700 m试验点,当育性转换敏感期越过8月上旬,育性将引起波动;实际上同期播期材料在生育进程上随着海拔的升高相应推后,在所有供试材料中,5月14日之后播种,越过海拔500 m点,8月下旬及之后是其育性转换敏感期将对育性转换会产生波动。 两系法杂交水稻制种与三系法制种相比,二者在技术层面上有不少相似之处,两系对种子纯度的监控除与三系相同外,本身又有其独特性,纯度风险主要来源于不育系自身的光温特性。外界环境温度在不同地域、不同年份、不同季节有很大差异且年际间波动较大,不育系在育性表达上显示出不稳定状态,这是由核不育系自身特性所决定的,因而对两系杂交制种的保纯技术要求较高。种子纯度能否过关,关系到杂种优势的利用与推广,实施两系杂交种子纯度监控是制种过程中非常重要的技术环节[16]。因此选择适宜的生态区域进行制种,重点要根据核不育系育性转换特性,从时间和空间上对制种区域进行生态条件研究[17],除了研究适宜的水、肥条件及安全的抽穗扬花期外,最为重要的是不育系育性敏感期的气候条件必须保证不育系完全不育以保证制种的纯度,即把核不育系的育性转换敏感时期控制为败育状态,这样才能保证种子的纯度无核不育系自交结实种的混杂。当然,影响杂交稻种子纯度的因素很多,诸如人工混杂、机械混杂等人为可控因素外,核不育系自身遗传漂变及环境的不可控性在两系杂交稻制种过程中对纯度的影响尤为重要,只有适时对核不育系进行核心提纯复壮外,掌握制种中育性敏感安全期内的温度状况,才能从源头上对两系杂交稻种子纯度提供有效保证。 两系杂交稻制种,是综合环境与人为双重因素的集成技术,它是在适宜的生态条件下通过人为优化方案实行异交栽培方式[18]来获取极限产量。高产制种技术是追求合理配置下的高产栽培模式,其中对父母本特征特性的了解,可操作的技术方法摸索和集成技术的灵活运用,包括对父母本花期相遇的播差期安排、花期预测与调控、高产异交群体结构的构建、异交态势的改良、父母本花时动态与人工辅助授粉等关键技术环节。因此,提高制种单产不仅是降低种子成本、加快科技成果转化、实现种子产业化的有效途径,而且是实现社会效益、经济效益和生态效益的有利保证。 水富县作为云南两系杂交稻制种基地[19-20]其独特的生态地理环境为两系杂交稻制种提供了很好的光温条件,典型的丘陵梯田结构为高产群体提供了良好的光合环境,但是丘陵状梯田的分布在局部区域也形成了不同的小气候环境,对两系杂交稻制种带来一定的制约因素。多年来,通过对当地生态条件的深入研究,掌握了部分核不育系在此生态条件下的育性转换规律,已经形成了一套低纬高原梯田两系杂交稻高产制种技术体系。通过种子入库数核算,高产户最高产量达7.34 t/hm2。安全高产技术集成为云南低纬高原梯田两系制种基地的选择及建立奠定了坚实基础。 低纬高原两系杂交稻制种,首先应该考虑选择育性转换起点温度低的核不育系;其次,制种区域控制在海拔500 m以下、播期5月14日之前较为安全。当海拔高于500 m时,播期相应提前,把核不育系育性转换敏感期控制在8月中旬以前。当海拔高于700 m时,不适宜两系杂交稻制种。

2.2 核不育系的育性稳定性分析
2.3 安全制种区域及适宜播期时段的选择
3 讨 论
3.1 核不育系自身育性转换特性决定了对适宜制种生态区域的选择
3.2 选择适宜生态区域进行两系制种是保证种子纯度的首要因素
3.3 成熟的制种技术体系是低纬高原梯田两系制种的有力保障
4 结 论
猜你喜欢
作物学报(2022年9期)2022-07-18
粉末冶金技术(2021年3期)2021-07-28
作物研究(2021年2期)2021-04-26
今日农业(2020年17期)2020-12-15
今日农业(2020年15期)2020-12-15
云南农业(2020年4期)2020-05-16
农村百事通(2019年5期)2019-03-25
中学生数理化·高二版(2016年6期)2016-05-14
人间(2015年22期)2016-01-04
同位素(2014年2期)2014-04-16
