
水鸟栖息地影响因素及修复技术研究综述
2019-01-10 01:08:12陈庆
湿地科学与管理 2018年4期
陈庆
(深圳市福田红树林生态公园,深圳市红树林湿地保护基金会,广东 深圳 518000)
近几十年来,由于气候变化和人为活动的加剧,全球面临着严重的自然湿地损失及退化。国家林业局第二次全国湿地资源调查结果显示全国湿地总面积5 360.26万hm2,湿地率5.58%,受保护湿地面积2 324.32万hm2,占湿地总面积的43.51%,主要受到污染、围垦、基建占用、过度放牧、过度捕捞和采集、外来物种入侵等威胁因子的影响。基于湿地国际的数据库,中国260种水鸟中,其中84种水鸟(占32.3%)的种群下降,125种水鸟(占48.1%)的种群趋势未知。38种水鸟(占15.1%)处于受威胁状态:6种极危(CR);16种濒危(ER);16种易危(VU)。受威胁水鸟中有32种受到栖息地丧失的严重威胁(Wang et al,2018)。
当自然湿地质量下降或不适合水鸟栖息时,人工湿地成为对自然湿地丧失的补充(Ma et al,2004)。水鸟可利用各种类型的人工湿地,如稻田(Elphick et al,1998)、 鱼 塘(He et al,2016)、盐田(Velasquez,1992)、城市人工景观水面(Traut et al,2003)。
2016年我国水产养殖面积达8.34×106hm2,其中海水养殖2.17×106hm2,淡水养殖6.18×106hm2;海水养殖产值3 140.39亿元,淡水养殖产值5 813.18亿元(2017中国渔业统计年鉴)。从20世纪80年代至2010年,我国沿海各省市的养殖塘面积都在明显增加,广东省的养殖塘面积相对增幅最大 ;黄河三角洲沿海新增的养殖池,主要是由滩涂、盐沼和耕地等转化而来(姚云长等,2016)。在天然湿地面积及质量下降的背景下,如何通过有效的人工湿地,例如沿海养殖塘管理,为水鸟提供高质量的栖息地并维持其种群,维持养殖塘的可持续利用是一个至关重要的问题。
1 栖息地尺度下影响水鸟利用湿地的因素
1.1 湿地面积大小的影响
湿地面积直接影响物种的多样性,湿地通常有不同的空间形态,湿地面积的变化影响到可利用栖息地的变化,从而改变利用该湿地的鸟类组成成分(Paracuello,2006)。一般而言,大面积的湿地具有更为丰富的水鸟,这通常是由于大面积的湿地能提供更为多样的微生境,形成不同的生态位(Zou et al,2017)。“一块大的湿地VS多块小湿地 ”(single-large or several-small,SLOSS)的争论由来已久(Soulé et al,1986)。鸟类的丰度在鸟类占用湿地的能力中起着重要的作用(sampling hypothesis,抽样假设)。当湿地面积减少时,水鸟种类的减少模式是有序的,与每一物种的丰度存在关系(Paracuello et al,2004)。改善小块湿地生境异质性的管理政策,有利于水鸟丰度的维持,这也意味着小块湿地面积必须足够大,能满足这些物种的最低要求。
1.2 湿地水文的影响
湿地水文是影响水鸟栖息地适宜性的重要因素,主要体现在水深、水面面积及水位变化。由于水鸟自身形态特征的限制,如腿长、喙长、颈长等,水深直接决定了水鸟可觅食栖息地。涉禽通常偏好浅水区,而潜水的鸟类需要更深的水位。不同类群的水鸟对水深的需求不同,这对于多物种的管理而言是一个挑战,湿地管理者需要依据湿地鸟类的主要类群进行有效管理。
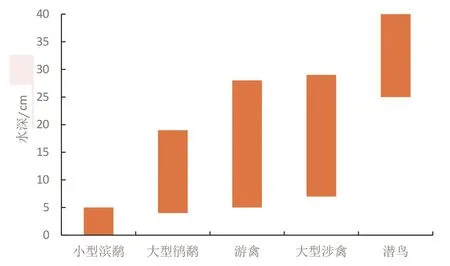
图1 不同类型水鸟觅食适宜水深(引自Ma et al,2010)Fig.1 Variation of water depths at foraging sites among waterbird groups
从图1可以看出,小型滨鹬类在不到5 cm深的水中觅食;大型鸻鹬(如黑尾塍鹬)在水深达15 cm的水中觅食;大型涉禽(如苍鹭、白鹭和鹮类)和游禽(如绿头鸭)在水中觅食深度达30 cm。潜水鸟类(如鸬鹚和䴙䴘)需要最低水深大于25 cm,甚至可在几米深的水中活动。
水位波动可能是由于季节性的洪水、潮汐、农业灌溉或排水引起。水位波动会引起水深和水面面积的变化,适应不同水深的鸟类利用也会发生相应的变化(Canepuccia et al,2007)。例如鄱阳湖越冬候鸟栖息地面积受到水位波动的影响,这种关系是非线性的,对于鸟类而言有一个适宜的水位变动范围(Wang et al, 2013)。潮汐是影响水鸟在滩涂栖息觅食的一个重要因素,水动力带来丰富的食物资源,退潮后的滩涂是水鸟重要的觅食场所,潮水上涨时水鸟随着水线移动,逐步转移到滨海湿地临近的盐田、农田、鱼塘等,潮汐周期性影响着水鸟对滩涂的利用。长期的水位变化反映了水鸟栖息地的适宜性,而栖息地的利用在短期内会受到水位季节性和年际变化的影响。虽然水位波动可能对水鸟栖息地的利用产生一些影响,但水位波动对湿地生产力的长期维持至关重要(Markham, 1982)。
1.3 湿地植被的影响
植被能为植食性的水鸟提供种子、块茎,例如在印度,白鹤Grus leucogeranus主要以香附子Cyperus rotundus块茎为食,在鄱阳湖也主要取食苦草Vallisneria spp.块茎(吴建东等, 2013)。此外,作为初级生产者的植物,能为无脊椎动物提供能量来源,而这些无脊椎动物也可能成为水鸟的食物来源。部分鸟类会在湿地植被中筑巢繁殖,如震旦鸦雀Paradoxornis heudei通常选择在茎秆较为粗壮的芦苇上筑巢,而草鹭Ardea purpurea则会选择将巢筑在芦苇茎秆细密的地方(李东来等,2015)。树岛也能为水鸟提供栖木(Bancroft et al, 2002),如美国大沼泽的白鹮Threskiornis melanocephalus,香港米埔的鸬鹚Phalacrocorax carbo。湿地边缘的挺水植物可以提高隐蔽性,形成水鸟主要休息区域与周边环境的隔离,降低人为活动的影响。
高密度的植被覆盖并非总是对水鸟有益。崇明东滩发现裸地面积比例是影响越冬水鸟种类分布的最主要因子:尽管游禽在地形变异较大、植被面积比例较低的区域数量较多,但在休息时游禽更偏好于裸地面积比例较高的区域;涉禽休息时偏好于地形变异较大的区域(金欣等, 2013)。美国南部、北部的“半沼泽”也发现在植被水面比为50:50时最利于水鸟的越冬繁殖(Smith et al, 2004)。美国内斯加州雨水盆地(RWB)也发现有中度(50%)植被覆盖的湿地游水鸭类数量最大,且此时鸟类的物种丰度最高(Webb et al, 2010)。鸟类利用湿地可能与植被的结构和覆盖模式有更紧密的联系,而不是植物群落的物种构成(Fairbairn et al,2001),但具体植被覆盖比例依然与利用地域的鸟类及其利用方式有关(Fujioka et al, 2001)。
1.4 食物可获得性的影响
水鸟存在不同的食物偏好:鹤类喜好以种子为食;鸭类大多为杂食性;小型鸻鹬类多取食无脊椎动物 ;鸬鹚多以鱼类为生。受栖息地影响,鸟类可能会改变其食谱,食物大小也会影响鸟类取食策略(Choi et al, 2015)。
食物的可获得性受限于鸟类自身的形态特征,如腿长、喙长等(Barbosa et al,1999);也与栖息地自身食源丰富度,食物的质量、营养和适口性有关,此外还需要考虑到食物的可摄食时间、鸟类处理食物的能力和能量摄食需求。沿海岸线的潮间带一直是水鸟长距离迁徙重要的栖息地,在非繁殖季节,它们倾向于选择具有高生产力的地区(Hurlbert,2010)。由于无脊椎动物在平均水位线附近最为丰富,相应水鸟大部分也分布在此范围(Baldwin et al,1994)。潮汐对食物可获得性的影响可能会限制水鸟在潮间带的食物获取,增加鸟类对替代喂养地点的依赖。
1.5 人为干扰的影响
人为干扰可能直接影响鸟类的种群结构与动态。捕杀直接杀死了鸟类个体,会改变配偶和家庭结构,从而影响到鸟类的子代,导致鸟类种群数量及结构变化(Madsen et al, 1995)。一些沿海养殖塘采用张挂围网的方式防止水鸟取食,大量的死鸟出现在单细丝大网孔(>5 cm)的网中(Nemtzov et al, 2003)。人类取食几乎造成了黄胸鹀Emberiza aureola的灭绝。
人为干扰也可干扰鸟类栖息地,间接影响其取食、繁殖、迁徙(颜凤等,2017;Steven et al,2011)。在人为活动频繁的码头游船区,骨顶鸡用于取食的时间分配明显减少(哈丽亚等,2014);繁殖期骨顶鸡躲避行为明显增加而孵蛋行为时间明显减少(程鲲等,2018)。围填海改变了土地利用类型,造成生境变化。中国渤海地区人口密集,天津新区和曹妃甸的开发围垦使得北迁候鸟,如红腹滨鹬Calidris canutus只能集聚在狭小的区域,遗鸥Larus relictus转移到未开发的唐山地区(Yang et al, 2011)。
水鸟种群对不同类型干扰的响应不一,电动交通工具通常较步行具有更短的逃跑启动距离(McLeod et al,2013)。不同鸟类对干扰的敏感程度也不同(Blumstein et al,2003),一般而言,体型较大的鸟类通常具有较大的启动距离(Rodgers et al,2002),美国莫诺莫伊国家野生动物保护区红腹滨鹬Calidris canutus的启动距离约为18 m,半蹼滨鹬Calidris pusilla约11 m(Koch et al,2014);加纳莫尔国家公园(Mole National Park)苍鹭Ardea cinerea的最远警戒距离为120.8 m,逃跑启动距离为32.7 m(Aikins et al, 2018)。对许多湿地而言,涉及多物种的管理,参考人类利用情况、各地各物种湿地利用高峰及逃跑启动距离,可以为缓冲距离的设定提供一定指导。此外,干扰的频度强度、集群大小等都会对水鸟产生不同的影响(Yasue, 2005)。
2 养殖塘鸟类栖息地修复技术
2.1 鸟类栖息地修复策略
湿地创造和修复项目取得成功的基本要求是:了解湿地功能;给系统的时间;并考虑到自然的自我设计能力(Mitsch et al, 1996)。湿地修复基于众多的生态学理论:岛屿生物地理理论、生态位、种群、营养级。自然栖息地为湿地修复提供了良好的参照系,在修复过程中必须考虑环境条件和生命历史特征,尤其是湿地水文对恢复湿地生物多样性与功能至关重要(Zedler, 2000)。
鸟类的迁徙具有季节性,对湿地生态系统的冬季管理而言,管理者需要为不同的水鸟群体(从小型鸻鹬类水鸟到潜水的鸟类)提供栖息地。中国的滨海湿地保护对于东亚-澳大利西亚迁徙路线的候鸟保护具有不可替代的重要作用,人为活动造成了候鸟栖息地的丧失(雷光春等, 2017),适当改造沿海养殖塘为水鸟提供了可利用的生境(华宁等,2009)。
2.2 养殖塘水鸟栖息地修复技术
养殖塘鸟类栖息地恢复技术主要是异质性生境单元的构建和食源供给两大方面,依主要保护对象的特性,按照鸟类栖息、繁殖和觅食活动的生境需求,分别进行微地形改造、底质改造、水位控制和补充食源地配置。微地形改造主要是通过挖掘或回填等方式,构建不同水深环境,以提高生境的异质性。水域、裸滩、植被的比例由目标物种决定,大量的科学研究为这一比例提供了参考,也需要依据当地湿地特征及利用湿地的鸟类情况进行调整。有时,也可通过构造不同标高、具有更大周长/面积比的鸟岛来实现栖息地的空间异质性。一般来说,地形的塑造(例如,在最深和最浅的区域之间的差异是30~40 cm),湿地的平均深度为15~20 cm,应该能容纳最大的水鸟丰度(Taft et al,2002)。枯木或倒木也是重要的小型生境单元,能够为鸟类提供栖木。微地形和鸟岛可以构造不同水位深浅的缓坡水域,但水位的调控依然是鸟类栖息地恢复与管理的核心内容。植被控制包括食源性植被恢复、生态隔离带植被恢复和干扰性植被控制。按照主要保护鸟类和优势水鸟的觅食习性,恢复相应的食源性水生植被和外围保护隔离带植被;同时,控制地面植被的高度及覆盖面积,以免侵占水鸟的栖息觅食空间。水鸟对养殖塘的利用也表现出随潮汐起落的周期性。养殖塘内的鱼虾可以作为水鸟的补充食物来源,一般而言,人工投喂这种方式是不推荐的。养殖塘内食物资源有限,且不能像滩涂一样天然更新,所以水鸟对养殖塘的利用主要作为高潮位的停歇地,在潮水退去后水鸟则又返回滩涂。

图2 水鸟栖息地修复流程Fig.2 Process of waterbird habitat restoration
3 展望
2016年我国发布了《湿地保护修复制度方案》,严格湿地用途监管,确保湿地面积不减少,增强湿地生态功能,维护湿地生物多样性,全面提升湿地保护与修复水平。我国各级湿地保护管理单位也在积极推进湿地保护法规政策和制度建设,强化湿地保护管理,加强湿地保护科技支撑,加大湿地保护力度,建立湿地保护多元化投入机制,开展湿地保护宣传教育,加强湿地国际合作与履约。
人工湿地的修复能为水鸟提供可利用生境。养殖塘水鸟栖息地常常面临着与当地经济的冲突,寻找湿地恢复与社区发展的结合点,建立社区参与式的湿地保护恢复长效机制是栖息地尺度湿地与鸟类保护与维持的保障,实现湿地的合理利用。
湿地修复前后的监测与评价,对于管理者而言至关重要,可以帮助判断恢复过程是否有效,在效果不理想时能及时诊断并采取措施。主动管理是栖息地维持的重要途径,有时一个恢复项目在短期内已被判断为成功,但在之后的几年里缺乏积极的主动管理,尤其是水文管理下将会失去原有的效果。鸟类的栖息地,尤其是人工湿地需要长期的监测评估与适应性管理来维持。对于迁徙候鸟而言,应该从迁飞线路的尺度考虑更大尺度的毗邻集水区、流域、沿海岸带湿地生态系统的结构与功能。
猜你喜欢
疯狂英语·初中版(2023年7期)2023-08-18 05:01:35
学与玩(2022年9期)2022-10-31 02:54:08
幼儿教育·父母孩子版(2022年3期)2022-03-14 17:08:51
文苑(2020年12期)2020-04-13 00:54:14
小太阳画报(2019年1期)2019-06-11 10:29:48
汉语世界(The World of Chinese)(2018年3期)2018-10-22 01:50:04
厦门航空(2018年4期)2018-04-25 10:49:27
小学生必读(低年级版)(2017年5期)2017-08-12 03:47:07
骏马(2016年6期)2017-01-09 08:48:46
启蒙(3-7岁)(2016年10期)2016-02-28 12:27:12
