
退化高寒草甸植物功能群与土壤因子关系的冗余分析
2019-01-07 08:55:16刘育红杨元武
生态与农村环境学报 2018年12期
刘育红,杨元武,张 英
青海大学农牧学院,青海 西宁 810016)
高寒草甸是在青藏高原特有的气候、地理、植被等条件下经长期演化和发展形成的独特生态系统。近几十年来,在全球气候变化、草地载畜量增加等因素共同作用下,高寒草甸呈退化态势,表现为群落逆向演替,草地生产力降低,退化草地面积增加等,不仅削弱了其生产和服务功能,而且使得区域生态环境恶化[1-3]。在此背景下,高寒草甸生态环境问题成为草地生态学领域的研究热点。一段时间以来,研究集中于退化草甸植物群落特征[4]、土壤化学性质[5]、土壤有机碳及组分[6-7]等方面。
植物功能群(plant functional groups,PFGs)是基于植物分类和植物形态、生理、生活史等特性划分的对环境因子有相似反应、具有某些确定功能特征的物种群。将PFGs作为研究植物随环境变化的基本单元,可揭示PFGs在生态系统中的作用、功能及其与生态系统功能的关系[8]。针对PFGs方面的研究包括森林生态系统PFGs特征的分析[9]以及土壤氮磷含量对PFGs叶片氮磷比的影响[10]等。在草地生态系统退化研究中,PFGs可以用来表征草地群落结构的复杂性和稳定性[11],如添加氮素和水分[12]、添加肥料[13]对草地PFGs地上生物量的影响,不同载畜率对荒漠草原群落及植物功能群生物量的影响[14],捡拾牦牛粪对高寒草甸植物功能群生产力的影响[15],放牧干扰对草地PFGs组成时空变化的影响[16],土壤有机碳含量与PFGs数量特征间的关系等[17]。已有研究大多探讨草地PFGs生物学产量、组成、作用、数量特征等对人为干预措施的响应,而将退化高寒草甸土壤中有代表性的化学、物理因子结合在一起,探讨多种土壤因子的共同作用与退化高寒草甸PFGs间关系的研究鲜见报道。所以,研究退化高寒草甸PFGs与土壤理化因子间呈现怎样的关系、退化高寒草甸PFGs与土壤理化因子间关系的密切程度如何、哪些土壤因子对退化高寒草甸PFGs的变化更重要等问题具有科学探索意义。因此,该研究在具有典型高寒草甸特征的研究样地进行植物群落数量特征观测及土壤样品采集与分析,以数量生态学排序理论为依据[18],运用冗余分析(redundancy analysis,RDA)方法,研究退化高寒草甸PFGs与草地土壤因子的内在联系,以期为退化高寒草甸的相关研究和生态治理提供理论依据,也为高寒地区草地群落、物种与环境间关系的研究提供一些实例。
1 材料与方法
1.1 研究区概况
研究区位于青海省果洛藏族自治州玛沁县、甘德县和达日县。果洛州(32°21′~35°45′ N,96°56′~101°45′ E)草地面积625.1万 hm2,其中高寒草甸占56.3%;全州平均海拔4200m;属高原大陆性气候区,年均温-4℃,年均降水量513.2mm,年均蒸发量1462.4mm,年均日照时数2260.3h;高寒草甸原生草地植物群落中莎草科高山嵩草(Kobresiapygmaea)、矮嵩草(Kobresiahumilis)等为优势种,其他植物属于禾本科(Gramineae)、菊科(Compositae)、豆科(Leguminosae)、蔷薇科(Rosaceae)等;高寒草甸草地广泛分布高山草甸土。
1.2 样地设置
在具有典型高寒草甸特征的研究样地,按照空间分布代替时间演替[19]及任继周[20]、赵新全[21]对高寒草地退化程度划分的方法,依据群落植被盖度(未退化、轻度退化、中度退化、重度退化、极度退化盖度分别为>90%、>70%~90%、>50%~70%、>30%~50%、≤30%)、可食牧草比例(未退化、轻度退化、中度退化、重度退化、极度退化可食牧草比例分别为>70%、>50%~70%、>30%~50%、>15%~30%、≤15%)、土壤侵蚀状况(未退化、轻度退化、中度退化、重度退化、极度退化土壤侵蚀状况分别为极轻侵蚀、轻度侵蚀、中度侵蚀、重度侵蚀、严重侵蚀)等指标综合评价,将研究样地归为5种退化程度,即未退化(un-degradation,UD)、轻度退化(light degradation,LD)、中度退化(moderate degradation,MD)、重度退化(heavy degradation,HD)和极度退化(extreme degradation,ED)。研究样地为高山嵩草草地型,优势植物种为高山嵩草、矮嵩草、线叶嵩草(Kobresiacapillifolia)等;样地土壤为高山草甸土;样地均为阳坡,坡度9°~17°,研究样地大小为50 m × 30 m。研究样地基本情况见表1。
1.3 测定方法
2017年7月,在每个研究样地内垂直等高线布设间距10 m、长度50 m的2条平行观测样线,每条样线间隔10 m布设一个1 m×1 m观测样方,样方重复5次。其中一条样线的样方用于观测植物群落数量特征,另一条样线的样方测定土壤容重、土壤含水量等指标并采集土样。
植物群落特征观测包括样方植被总盖度(多人目测平均法),植物物种数、分物种盖度和高度(自地表至植株顶端自然高度,每样方测定20株,不足20株的物种按实际株数测定)等;植物地上生物量(分物种齐地面刈割称鲜重)。采集土样时,每样方内按对角线土钻采集0~30 cm深度土样5钻,同一样地土样混合为一个土壤样品;采集土样的同时,每样方内采用环刀法测定土壤容重,测温计测定土壤温度(15至16时温度),水分测定仪(TDR300)测定土壤含水量(当日无连续性降水时)。土壤有机碳含量测定采用重铬酸钾容量法,全氮含量测定采用半微量凯氏定氮法,全磷含量测定采用钼锑抗比色法,有效氮含量测定采用比色法,有效磷含量测定采用碳酸氢钠浸提钼锑抗比色法,全钾和有效钾含量测定采用火焰光度法,土壤颗粒组成测定采用比重计法,pH值测定采用水土体积比2.5∶1的电位法[22-23]。
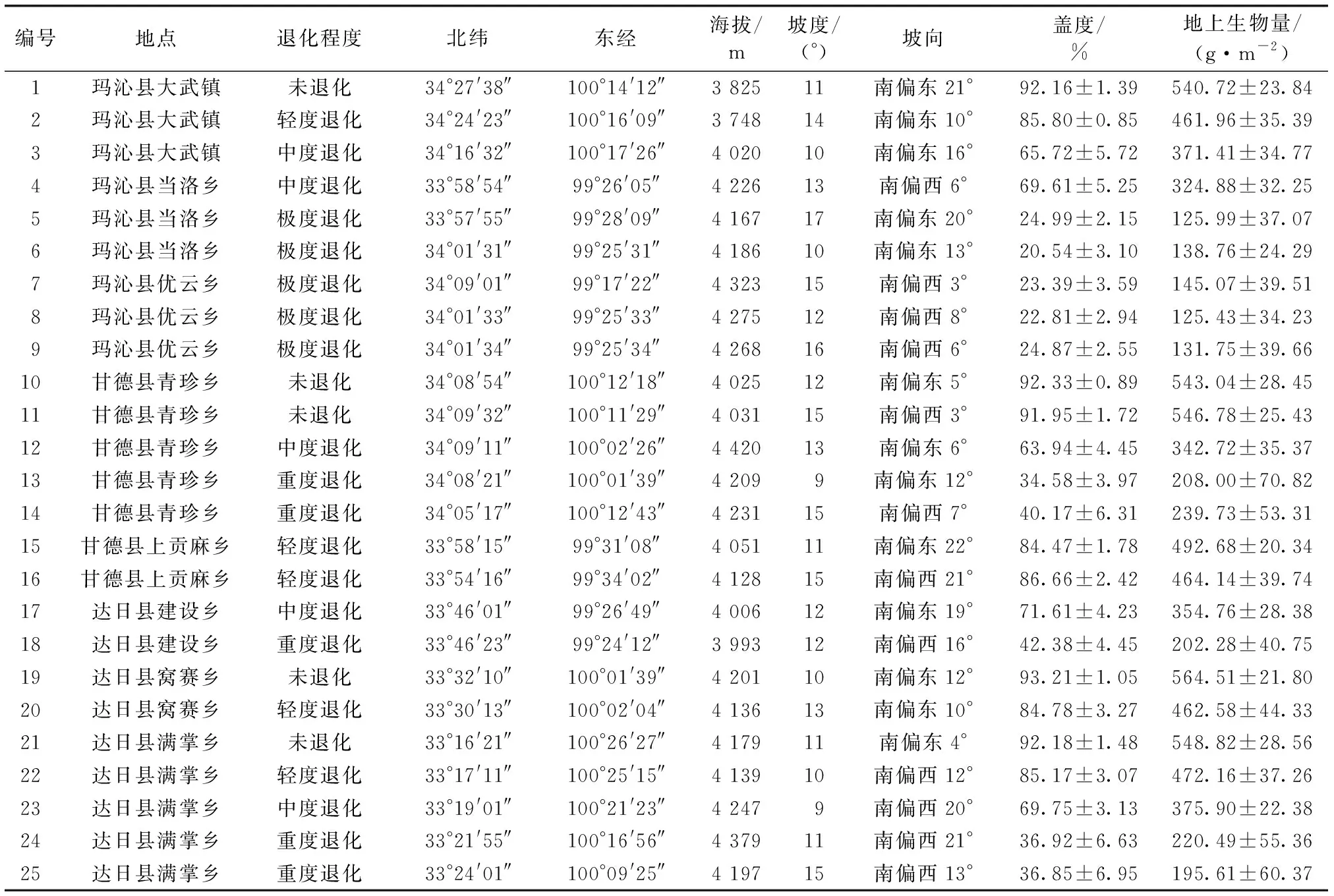
表1 研究样地基本情况Table 1 Basic information of the study area
1.4 PFGs的划分
PFGs的划分一般采用非系统发育分类法,且无统一的划分标准。研究参考王长庭等[24]和赵建中等[25]的方法,将高寒草甸研究样地植物划分为莎草科(Cyperaceae)、禾本科(Gramineae)、豆科(Leguminosae)和杂类草(Forbs)4个功能群。
1.5 数据处理与分析
计算研究样地群落物种重要值(VI),PFGs重要值为该功能群内所有物种重要值之和。
VI=(Rc+Rh+Rb)/3。
(1)
式(1)中,Rc为相对盖度,其值为群落中某物种盖度/群落中所有物种盖度之和;Rh为相对高度,其值为群落中某物种平均高度/群落中所有物种平均高度之和;Rb为相对生物量,其值为群落中某物种生物量/群落中所有物种生物量之和。
应用Canoco 4.5软件对退化高寒草甸PFGs与土壤因子进行冗余分析[26]。其中物种数据为4种PFGs构建的4×25维重要值矩阵,用土壤主要理化因子构建13×25维土壤环境因子矩阵,包括土壤有机碳(SOC)含量、全氮(TN)含量、有效氮(AN)含量、全磷(TP)含量、有效磷(AP)含量、全钾(TK)含量、有效钾(AK)含量、容重(BD)含量、土壤含水量(SWC)含量、土壤温度(ST)含量、黏粒(粒径<0.002 mm)比例、砂粒(粒径0.05~2 mm)比例和pH值共13个指标。冗余分析时,排序轴特征值采用蒙特卡罗检验显著性,前向选择法检验13个土壤因子的边际作用[27],依土壤因子特征值进行排序;采用Cano Draw绘制PFGs-土壤因子RDA二维排序图;采用SPSS 20.0软件进行ANOVA分析,Duncan法进行多重比较。
2 结果与分析
2.1 退化高寒草甸植物种类
研究样地分布有13科26种植物,分别属于莎草科、禾本科、豆科、杂类草4种PFGs。植物物种数在LD样地最多,ED样地最少;物种重要值因退化程度和物种种类的不同而变化,UD、LD样地高山嵩草、矮嵩草等莎草科植物重要值较大,是群落优势种,HD、ED样地黄帚橐吾(Ligulariavirgaurea)、细叶亚菊(Ajaniatenuifolia)等植物重要值较大,演替为群落优势种,使得高寒草甸生产功能降低乃至丧失(表2)。
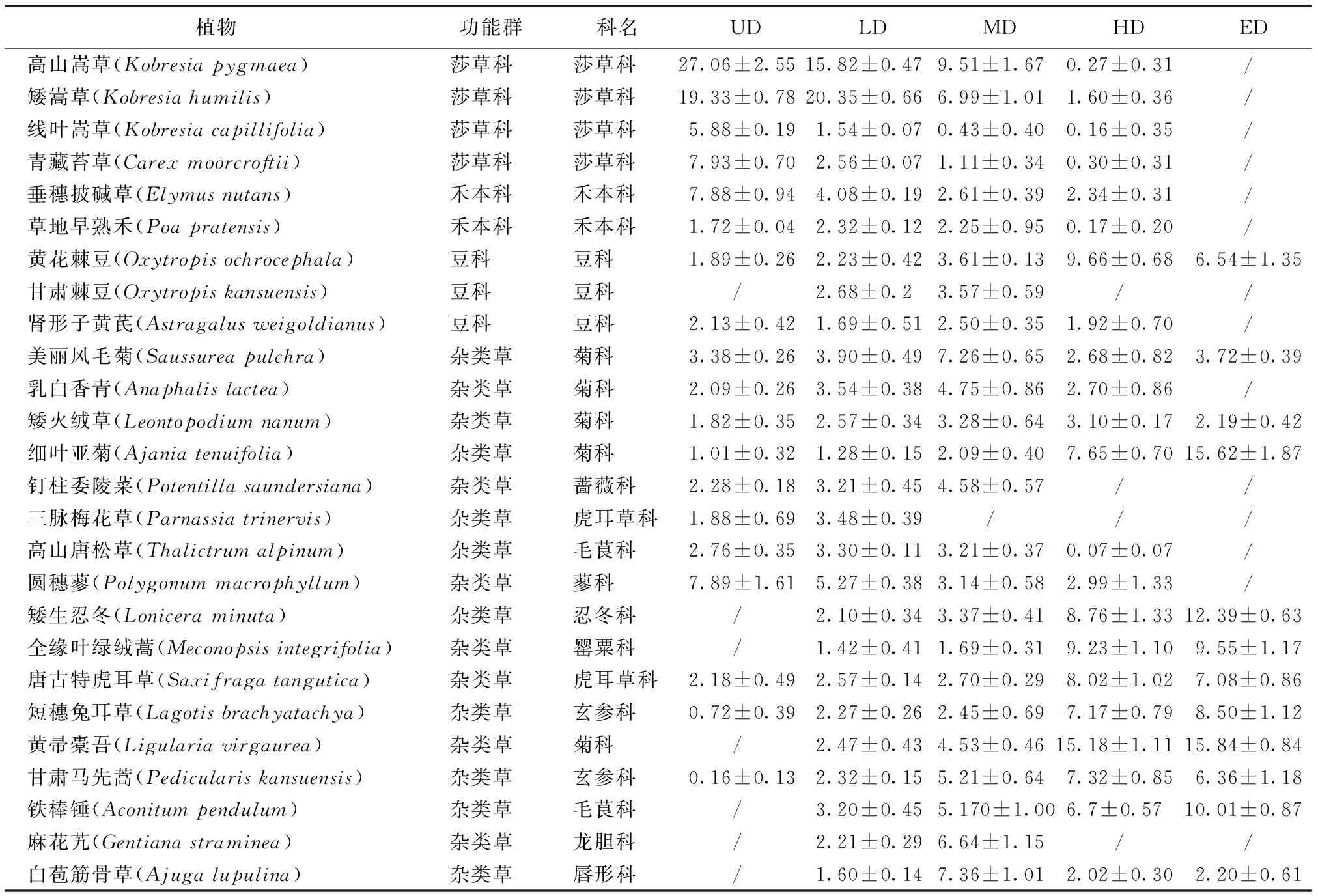
表2 研究样地植物种类及重要值Table 2 Plant species and important values of studyarea
2.2 退化高寒草甸PFGs重要值特征
随退化程度的加剧,研究样地莎草科、禾本科PFGs重要值减小,杂类草PFGs重要值增加,豆科PFGs重要值先增加后减少,不同退化程度间差异显著(P<0.05)。随退化程度加剧,高寒草甸群落中莎草科、禾本科PFGs逐渐被豆科、杂类草PFGs所取代,表现为高寒草甸植被退化;在极度退化发生时,莎草科、禾本科PFGs从群落中退出,高寒草甸PFGs种类减少,退化严重(表3)。
2.3 退化高寒草甸土壤因子特征
高寒草甸退化时,土壤化学、物理性质也发生相应变化。13个土壤因子在不同退化程度下变化趋势不同。SOC、TN、AN、TP含量与SWC均呈UD>LD>MD>HD>ED的变化趋势;AP、TK含量随退化程度的加剧先增大后减小;AK含量总体上table_title随退化程度加剧而减小;ST、BD、pH值总体上随退化程度加剧而增大;黏粒比例则先增大后减小,砂粒比例总体先减小后增大(图1)。
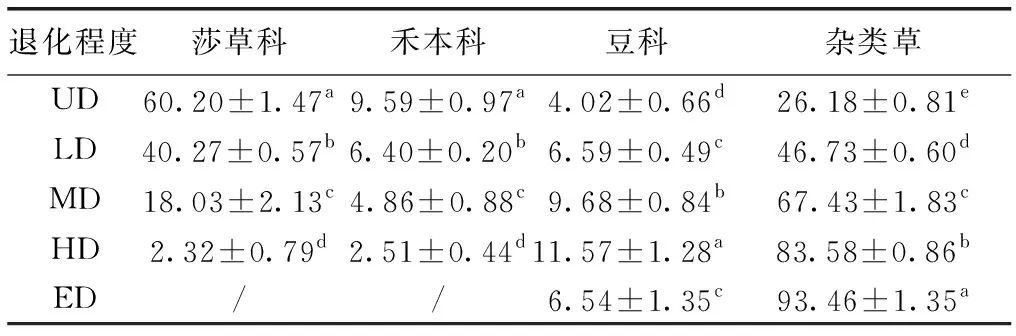
表3 退化高寒草甸植物功能群重要值Table 3 PFGs important values of the degraded alpine meadow
2.4 高寒草甸PFGs与草地退化关系
利用退化高寒草甸PFGs重要值和土壤因子数据矩阵进行冗余分析,RDA排序结果显示,25个高寒草甸研究样地沿第1排序轴因PFGs重要值、退化程度的不同分布在5个区域(图2)。说明RDA第1排序轴主要反映高寒草甸退化程度,自右向左退化程度加剧。
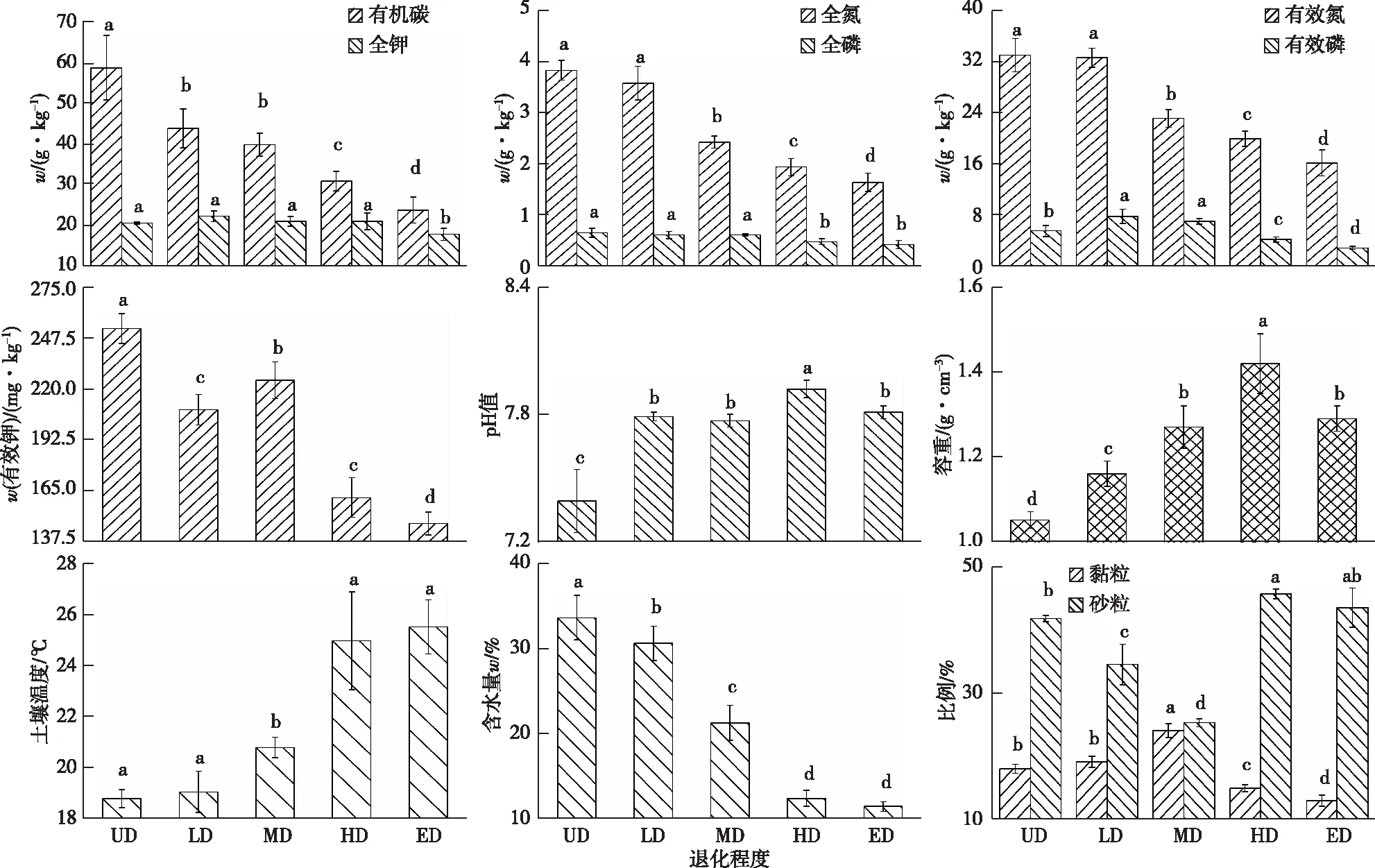
UD—未退化;LD—轻度退化;MD—中度退化;HD—重度退化;ED—极度退化。同一幅图中同一指标直方柱上方英文小写字母不同表示不同退化程度间某指标差异显著(P<0.05)。
RDA二维排序图显示,高寒草甸4种PFGs与不同退化程度样地间关系不同。莎草科、禾本科PFGs与UD、LD样地接近,与MD、HD、ED样地较远,反映出PFGs中莎草科、禾本科是UD、LD样地的重要植被类型;豆科PFGs与MD、HD样地更加接近,反映出PFGs中豆科在退化演替进行到中度、重度程度时具有较大的优势度;杂类草PFGs则与HD、ED样地接近,反映出杂类草PFGs在重度、极度退化程度下演替为高寒草甸植物群落中的主要类群,由此说明高寒草甸PFGs的演替能够反映草地退化状况。莎草科、禾本科PFGs重要值沿第1排序轴自右向左减小,反映出这2个PFGs与高寒草甸退化程度间呈负相关关系,豆科、杂类草PFGs重要值则沿第1排序轴自右向左增大,反映出这2个PFGs与高寒草甸退化程度间呈正相关关系。4个PFGs与高寒草甸退化的相关程度为莎草科>禾本科>杂类草>豆科。RDA第1排序轴主要反映莎草科、禾本科、杂类草PFGs的变化,第2排序轴则主要反映豆科PFGs的变化。
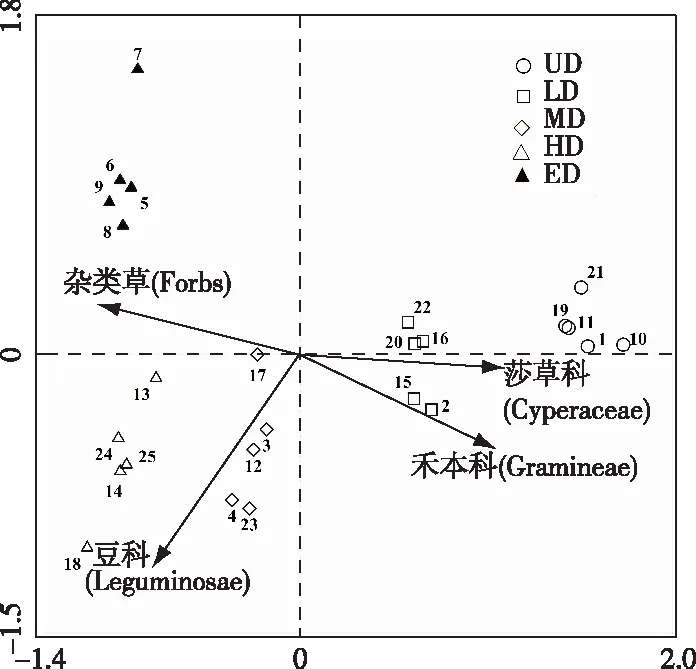
UD—未退化;LD—轻度退化;MD—中度退化;HD—重度退化;ED—极度退化。图中数字表示样地编号。
2.5 高寒草甸PFGs与退化草地土壤因子关系
高寒草甸PFGs与退化草地土壤因子冗余分析结果(表4)表明,RDA第1、第2排序轴共解释了97.1%的PFGs变化和99.6%的PFGs与土壤因子关系。第1、第2排序轴PFGs与土壤因子的相关系数分别为0.996、0.948,进一步反映出PFGs与土壤因子关系密切;蒙特卡罗检验也表明第1排序轴及所有排序轴反映的PFGs与土壤因子呈极显著相关关系(P<0.01)。
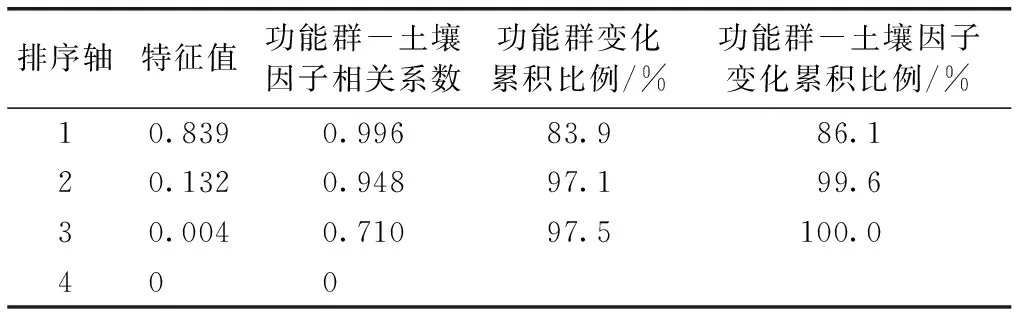
表4 高寒草甸PFGs与土壤因子的RDA二维排序结果Table 4 RDA ranking results of PFGs and soil factor under alpine meadow
RDA排序结果(图3、表5)显示,SWC、TN含量等与第1排序轴呈正相关,相关系数分别为0.952、0.945;BD、ST等与第1排序轴呈负相关,相关系数分别为-0.899、-0.854,说明第1排序轴主要反映SWC、BD等因子的综合变化。沿第1排序轴自右到左,ST升高,BD增大,TN含量、SWC等降低。黏粒比例、TK含量等与第2排序轴呈负相关,相关系数分别为-0.543、-0.510;砂粒比例与第1排序轴呈正相关,相关系数为0.342,说明第2排序轴主要反映土壤机械组成的综合变化,沿第2排序轴自下向上,黏粒比例减小,砂粒比例增大。
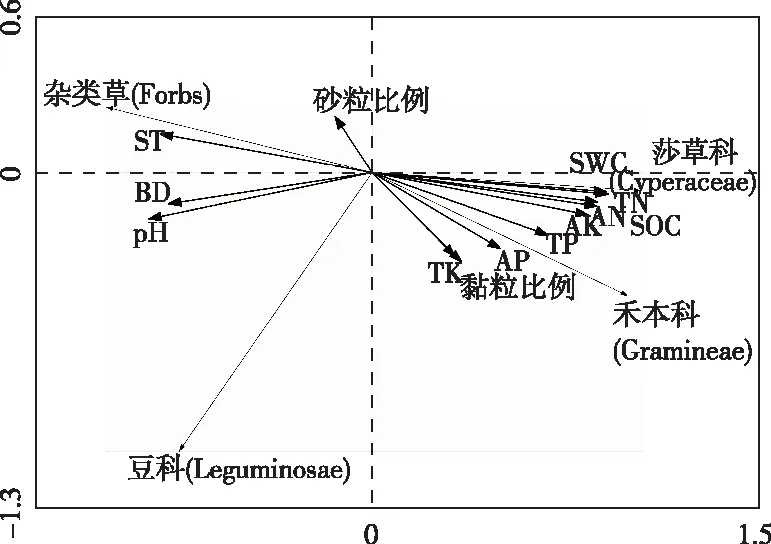
SWC—土壤含水量;SOC—有机碳;AN—有效氮;AP—有效磷;AK—有效钾;BD—容重;ST—土壤温度。
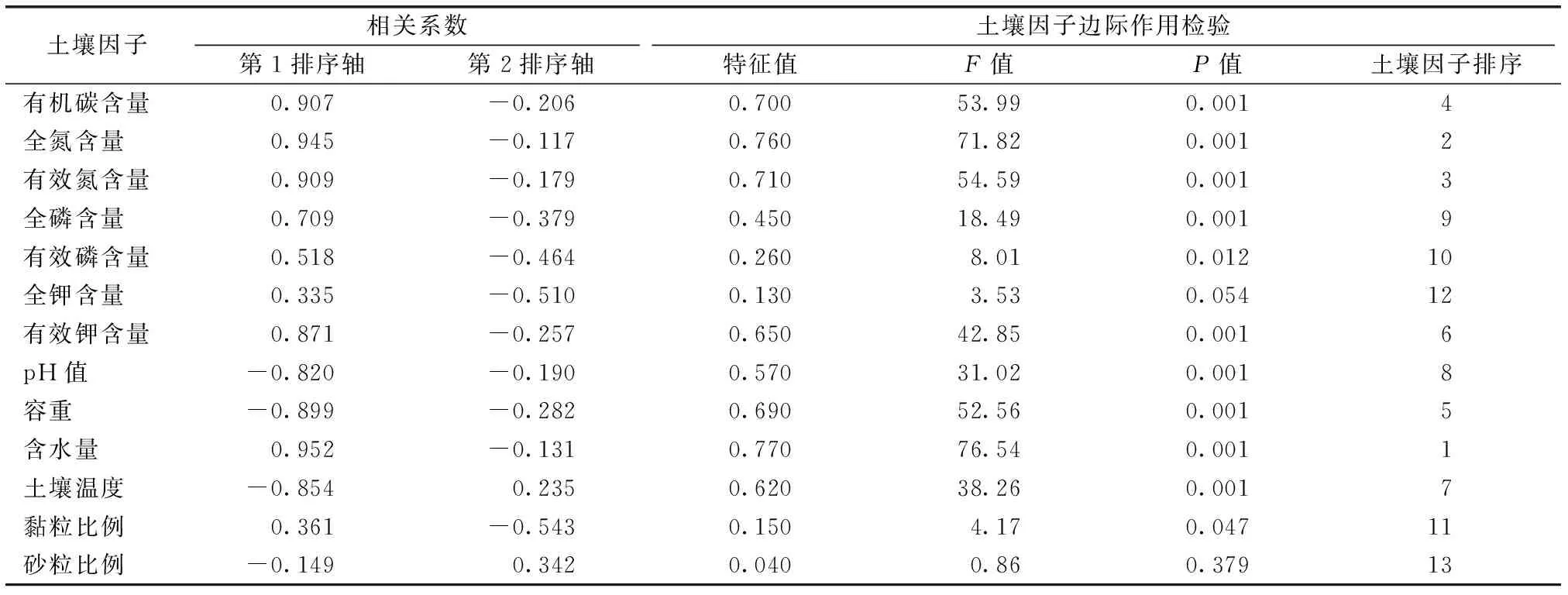
表5 高寒草甸PFGs与土壤因子RDA相关系数及土壤因子边际作用检验结果Table 5 Correlation coefficient between PFGs and RDA of soil factors as well as soil factors marginal effects tests under alpine meadows
对13个土壤因子边际作用检验并按特征值排序(表5)发现,不同土壤因子对高寒草甸PFGs的影响不同,反映出高寒草甸SWC等物理指标和土壤TN、AN含量等化学指标对退化高寒草甸PFGs的分布更重要,因子边际作用差异极显著(P<0.01);而土壤机械组成中黏粒比例、砂粒比例指标对PFGs分布影响较弱,因子边际作用未达极显著水平(P>0.01)。
由PFGs与土壤因子的RDA二维排序结果可知,莎草科PFGs受土壤TN含量、SWC等影响更大,与土壤TN含量、SWC的相关系数分别为0.960、0.959(P<0.01);禾本科PFGs受SWC、SOC含量等的影响更大,相关系数分别为0.937、0.929(P<0.01);说明SWC、TN、SOC含量较高有利于高寒草甸莎草科、禾本科PFGs的分布。豆科PFGs受BD、pH值等的影响更大,与BD等因子间相关性均达极显著水平(P<0.01);杂类草受ST、BD等的影响较大,与ST、BD等因子间相关性也达极显著水平(P<0.01);说明在ST、BD水平较高且SWC、TN、AN等含量相对较低水平下,高寒草甸群落中豆科、杂类草仍有一定的分布。
3 讨论
高寒草甸生态系统易遭受环境和人为因素的扰动而发生草地植被退化和土壤退化,退化高寒草甸植被与土壤因子的关系以复杂的动态形式存在。PFGs在群落中以一个相对统一的整体对环境因子的变化产生响应,当PFGs中个别物种由于环境干扰消失时,在一定限度内并不显著影响功能群的地位和作用。因此,PFGs能够客观反映包括退化高寒草甸植被与土壤因子间关系在内的高寒草甸生态规律。
3.1 退化高寒草甸植物功能群逆向演替
青藏高原高寒草甸是在特有的气候、地理、地质等环境条件下经长期演化和发展形成的独特生态系统[28],草地生产力低,系统稳定性差,自我修复能力弱,一旦发生逆向演替,恢复困难、缓慢,并由此引发一系列的生态环境问题。
高寒草甸退化是植物群落和草地土壤协同退化的结果,植物群落的退化既可以表现为物种种群的逆向演替,也可表现为PFGs的逆向演替。该研究发现,随着高寒草甸退化程度的加剧,群落中占优势的莎草科、禾本科逐渐被豆科、杂类草所取代,PFGs由莎草科、禾本科向豆科、杂类草演替,在极度退化样地植物群落中仅剩豆科、杂类草功能群。
导致这一现象的原因是多方面的。气候的演变通过影响PFGs结构、生长发育时期、生物量等成为PFGs演替的驱动力[29]。高寒草甸地区的气候暖干化,使得不同功能群植物的响应不同,在一定限度内的暖干化不利于莎草科生长,而对豆科和杂类草中的部分植物影响相对较小,使得PFGs由莎草科向豆科、杂类草演替。气候变化还会导致高寒冻土区冻土层上界下降,为啮齿动物等鼠虫的生存提供温床,加剧鼠虫危害,特别是高原鼠兔(Ochotonacurzoniae)危害严重[30],引起草地土壤结构、养分、水分等的变化,杂类草逐步侵入莎草科的生活空间[31],加速高寒草甸PFGs的逆向演替。
诸多研究人员还认为高寒草甸退化与放牧行为有关。不同放牧强度对高寒草甸群落和土壤的影响不同,放牧强度越大退化越重。YANG等[32]认为,不放牧和轻度放牧时,高寒草甸群落生物量、物种丰富度最高,随着放牧强度的增加,土壤总氮含量降低。由于高寒草甸群落中莎草科、禾本科功能群是优良牧草,草场超载过牧,势必影响莎草科、禾本科功能群的生长发育和生产能力,出现地上部生物量下降[33]、PFGs养分储存欠缺、致密草皮层破坏等问题,成为引起PFGs逆向演替的人为因素。放牧行为之所以导致PFGs逆向演替,还与莎草科、杂类草等功能群对放牧反应的差异有关。牛钰杰等[16]在研究了高寒草甸功能群对放牧的响应后认为,不同放牧强度对莎草科与杂类草的影响存在差异是导致草甸群落特征变化的原因;张小红等[34]对克氏针茅草原的研究也发现,随着放牧强度的增加,多年生禾草在群落中的优势地位下降。笔者的研究结论与这些研究相符。
笔者认为,随高寒草甸退化程度的加剧,PFGs中尚存的豆科虽对放牧生产不利,但从维护草地生态系统稳定性角度看,其具备的土壤氮素影响能力对高寒草甸土壤、植被的恢复和退化草甸的生态治理具有积极意义,这与LEGENDRE等[27]的观点一致。
高寒草甸PFGs的逆向演替既对当地畜牧业生产不利,也恶化了当地的生态环境条件,高寒草甸退化加剧的发展方向之一就是草地的荒漠化。要恢复与重建高寒草甸植被,使生态位宽度较小的莎草科功能群重新获得优势度并不是一件容易的事情,这也反映出退化高寒草甸治理的严峻性。因此,通过合理、科学、可行的途径对退化草地加以治理意义重大。
3.2 高寒草甸PFGs与退化草地土壤因子间存在响应与反馈的作用
高寒草甸退化不仅是植物群落的退化,草地土壤同样发生退化。该研究发现,随退化程度加剧,土壤有机碳、氮磷含量与含水量等总体呈下降趋势,而土壤温度、容重等总体呈相反变化趋势。 LI等[35]认为,高寒草地退化梯度对土壤含水量的影响最高;REN等[36]认为,土壤水分、养分含量因草地退化而减少。笔者的研究结论与上述研究一致。
在高寒草地植物群落和草地土壤退化时,PFGs与退化土壤因子存在相关性。研究发现,莎草科、禾本科功能群与土壤含水量、有机碳含量等因子呈极显著正相关,与土壤温度、容重等呈极显著负相关,豆科、杂类草与容重、土壤温度等呈极显著正相关,与全氮含量等呈极显著负相关。这一研究结果仅表明土壤含水量、全氮含量等因子水平相对较高,土壤温度等因子水平相对较低,对莎草科、禾本科PFGs的分布与生长更有利;而当容重、土壤温度等因子水平相对较高时,容重、土壤温度等对豆科、杂类草PFGs分布和生长的影响要小于莎草科等植物,并不代表豆科、杂类草PFGs就适合在养分含量较低条件下分布与生长。这一结论与ZHANG等[37]的研究结果一致。从高寒草甸退化的实际情况看,当退化严重时,土壤养分匮乏、土壤理化性质恶化,草地植物群落中所有PFGs都会受到极为不利的影响。由于PFGs的演替要早于草地土壤退化,而土壤物理性质的恶化及主要营养成分含量的降低会反馈于PFGs,使得竞争资源能力相对较弱的莎草科逐渐失去优势度,加速PFGs的演替进程。当然,引起高寒草甸PFGs与土壤退化的深层次原因及机理值得继续研究,方能服务于高寒草甸植被恢复重建及建立稳定的高寒草地生态系统。
3.3 PFGs可应用于高寒草地生态环境领域的研究
利用PFGs对生态环境问题开展研究时,PFGs的划分是一个重要的问题。可以依据植被群落中具有近似反应、作用和功能特征的物种归为一类这一属性,结合所研究问题的需要,对研究对象进行PFGs的合理划分,如SHEN等[38]从牧草的营养属性对高寒草甸植被进行了PFGs划分。在PFGs划分时,若要减少人为划分对研究结果的影响,可利用如聚类分析等客观方法加以辅助,以使PFGs的划分更科学。
该研究在选取影响植被的因子时,侧重选择土壤理化性质,而样地海拔、降水量等地形、气象因素未纳入分析中,应在今后的研究中给予补充,以期在更广的维度揭示退化高寒草甸PFGs与土壤因子间的关系,为研究和治理高寒草甸生态系统问题提供科学依据。
4 结论
(1)高寒草甸退化使得群落植物盖度、地上生物量等均发生变化,随草甸退化程度的加剧,PFGs中莎草科、禾本科重要值减小,杂类草重要值增大,PFGs由莎草科、禾本科向豆科、杂类草演替。
(2)随着高寒草甸退化程度的急剧,土壤有机碳、氮磷、含水量等总体呈下降趋势,土壤温度、容重等总体呈上升趋势。
(3)PFGs与高寒草甸退化的相关程度为莎草科>禾本科>杂类草>豆科,冗余分析解释了PFGs的变化以及PFGs与土壤因子间的极显著相关关系。
(4)退化高寒草甸土壤含水量、全氮含量等对PFGs分布的影响更显著,莎草科、禾本科受土壤全氮、有机碳含量及含水量等的影响更大,豆科、杂类草在土壤含水量、全氮含量等水平相对较低,容重等水平相对较高时仍有分布。
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
青海草业(2022年2期)2022-07-23 09:34:58
中学生数理化·七年级数学人教版(2022年11期)2022-02-14 07:14:12
现代园艺(2021年23期)2021-12-01 07:47:44
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
科普童话·学霸日记(2020年1期)2020-05-08 16:45:11
活力(2019年21期)2019-04-01 12:17:10
小天使·一年级语数英综合(2019年2期)2019-01-10 11:57:30
