
草鱼皮酶解工艺优化及产物促嗜热链球菌增殖和抗氧化性研究
2019-01-04 07:47葛俊苗李启乐代启虎陈建康蒋采贝张璐盛洁宋益善
山东农业大学学报(自然科学版) 2018年6期
葛俊苗,李启乐,代启虎,陈建康,蒋采贝,张璐,盛洁,2,宋益善,2,3*
草鱼皮酶解工艺优化及产物促嗜热链球菌增殖和抗氧化性研究
葛俊苗1,李启乐1,代启虎1,陈建康1,蒋采贝1,张璐1,盛洁1,2,宋益善1,2,3*
1. 上海海洋大学食品学院, 上海 201306 2. 上海水产品加工及贮藏工程技术研究中心, 上海 201306 3. 国家淡水水产品加工技术研发分中心(上海), 上海 201306
以草鱼皮为原料,利用风味中性蛋白酶酶解,并以酶解产物促嗜热链球菌增殖效果为指标,利用响应面分析对酶解工艺进行了优化。结果表明,酶解产物促进嗜热链球菌增殖的最佳酶解工艺为:酶解时间4.13 h,料水比1:10 (w/v),风味中性蛋白酶加酶量3.1%。另外,根据分子量大小将最佳条件下得到的胶原蛋白肽分为4段: >10000 Da;5000~10000 Da;3000~5000 Da;< 3000 Da,研究不同分子量的肽段对嗜热链球菌增殖作用。结果表明,分子量< 3000 Da的肽段相比其他分子量的肽段更适合促进嗜热链球菌生长。体外抗氧化性实验为进一步了解胶原蛋白肽特性,结果表明酶解产物对DPPH、超氧阴离子具有较好的清除能力。GFSPH Ⅳ既可促进益生菌增殖,还具有抗氧化的生物活性,可以为多组分功能性食品的开发提供理论依据。
嗜热链球菌; 风味中性蛋白酶; 多肽;响应面分析; 抗氧化性
我国是水产品养殖大国,2016年全国水产品总产量6699.65万t,其中淡水养殖产量3290.04万t,占全国水产品总量的49.11%。草鱼是我国四大淡水鱼之一,营养价值丰富,其产量占淡水鱼总产量的20.9%,是我国重要的水产品资源[1]。在水产品加工过程中产生了大量的副产物,包括鱼头、鱼皮、鱼鳍、鱼尾、鱼骨及残留鱼肉,其中鱼皮占有8%以上[2],然而我国的水产品加工率仅为30%,远远低于发达国家的80%以上[3]。研究表明,鱼皮中I型胶原蛋白含量最大可超过其含量的80%,比鱼体其他部位高许多[4],然而其脂肪含量却很低,采用酶解法从草鱼皮提取胶原蛋白肽,不仅提高了草鱼的综合利用价值,对进一步促进水产养殖业的可持续发展也具有重大的现实意义[5]。
近几年来,益生菌产品越来越受到消费者的青睐。嗜热链球菌是一种多功能益生菌,具有改善肠道微生态、调节血压、抗癌、延缓衰老、对胃酸及胆盐具有耐受性、缩短酸奶凝乳时间和增加酸奶黏度等特点,在乳制品工业中扮演着非常重要的角色[6-8]。嗜热链球菌的益生特性主要表现在被人体吸收后,定植在人体的肠道中,可调节肠道微生态(菌群)平衡,提高机体免疫力,促进人体健康[9,10]。但嗜热链球菌蛋白代谢能力弱[11],且大多数嗜热链球菌的生长都需要蛋氨酸、组氨酸、脯氨酸以及谷氨酸[12]。有研究表明寡肽是嗜热链球菌生长的最佳氮源[13,14]。
本实验以水产品废弃物草鱼皮为原料,研究其酶解产物对嗜热链球菌增殖效果。以促进嗜热链球菌增殖为指标,即在600 nm处的吸光值(Optical density, OD),利用风味中性蛋白酶酶解,并通过Box-Behnken中心组合设计对酶解工艺进行了优化。该研究成果可以为水产品新型益生菌功能食品的研发提供理论依据。
1 材料与方法
1.1 材料与试剂
草鱼(3~5 kg):购于上海市浦东新区农工商附近菜市场(鲜活)。风味中性蛋白酶(1946 U/mg):山东隆大生物工程有限公司;嗜热链球菌:购于上海鲁微科技有限公司。
试验中所有化学试剂均购于上海国药集团化学试剂有限公司。MSC300国产杯式超滤器:上海摩速科学器材有限公司。
1.2 实验方法
1.2.1 样品前处理将鲜草鱼置于装有冰块的密封箱,运输至实验室,宰杀后清洗干净,然后剥皮(全程在0 ℃冰水混合物中操作),获得的鱼皮用0 ℃蒸馏水洗至中性,再用吸水纸擦干表面水分,用剪刀剪至约0.2 cm×0.2 cm碎片,放-20 ℃冰箱,备用。
1.2.2 草鱼皮胶原蛋白肽制备工艺流程工艺流程:鱼皮切碎物→加蒸馏水混匀→热处理(100 ℃,30 min)→冷却至酶解温度→调pH值→加酶→酶解→灭酶(100 ℃,10 min)→离心(5000 r,20 min)→过滤→冷冻干燥。
本实验选用的草鱼皮,其蛋白质含量高达27.0%,而脂肪和灰分含量较低,仅占1.8%,0.3%,具有高蛋白、低灰分、低脂肪的特点[9,15]。考虑在实际产业化过程中,尽量减少操作步骤,本研究中原材料没有进行脱脂处理,而是通过4 ℃离心过滤操作除去大多数脂肪;酶解结束后采用100 ℃沸水处理10 min使酶失活。
1.2.3 单因素试验风味蛋白酶天然安全,反应条件温和、酶解产物品质高,可应用于各种动植物蛋白的酶解,后期风味优化,去除苦味,改善口感,采用风味中性蛋白酶可制取良好酶解产物,提高产品质量,降低成本。风味中性蛋白酶最适温度为50 ℃、最适pH为7.0,因此本研究主要考察了酶解时间、料水比、加酶量三个单因素对嗜热链球菌增殖效果的影响,选择酶解时间4 h,料水比1:10(w/v),加酶量4%,固定其中两个单因素,另一因素水平分别如下:酶解时间:1 h,2 h,3 h,4 h,5 h,6 h,7 h;料水比:1:5,1:10,1:15,1:20;加酶量:1%,2%,3%,4%,5%。
1.2.4 响应面试验在单因素的基础上,选出三种因素最佳因素水平进行响应面实验设计,采用Box-Behnken模型设计,优化促进嗜热链球菌增殖的酶解工艺。
1.2.5 不同分子量草鱼皮酶解物的制备酶解液离心过滤后,上清液用超滤膜进行逐级分离。膜的截留分子量分别为10 k Da,5 k Da和3 k Da,控制蠕动泵转速为30 rpm,超滤压力为0.1 MPa。分别收集四个组分GFSPH Ⅰ (>10 k Da)、GFSPH Ⅱ (5-10 k Da)、GFSPH Ⅲ (3-5 k Da)和GFSPH Ⅳ(< 3 k Da)。然后将各组分进行冷冻干燥,放-20 ℃冰箱,备用。
1.3 鱼皮酶解产物促进嗜热链球菌生长活性的测定
1.3.1 化学限定培养基基本组成为了直观测定酶解产物作为氮源对嗜热链球菌增殖效果,本实验选择化学限定培养基培养嗜热链球菌,化学限定培养基基本组成参考宋益善等[15]并稍加修改,如表1。
配置过程中,维生素和无机盐用0.22 μm膜过滤以除杂菌,其余配置成溶液后121 ℃高压灭菌30 min。维生素需避光保存,现配现用。
1.3.2 嗜热链球菌增殖培养 5 mL嗜热链球菌活化试管斜面加入1 mL无菌蒸馏水,震荡制备菌悬液;接种量为2%(v/v),37 ℃培养18 h,培养过程中摇床转速为120 r/min。
1.3.3 嗜热链球菌增殖效果OD值的测定采用紫外-可见分光光度计在600 nm处的吸光度值来表示嗜热链球菌的增值效果,调零基准为不加任何氮源、相同条件下的液体培养基。
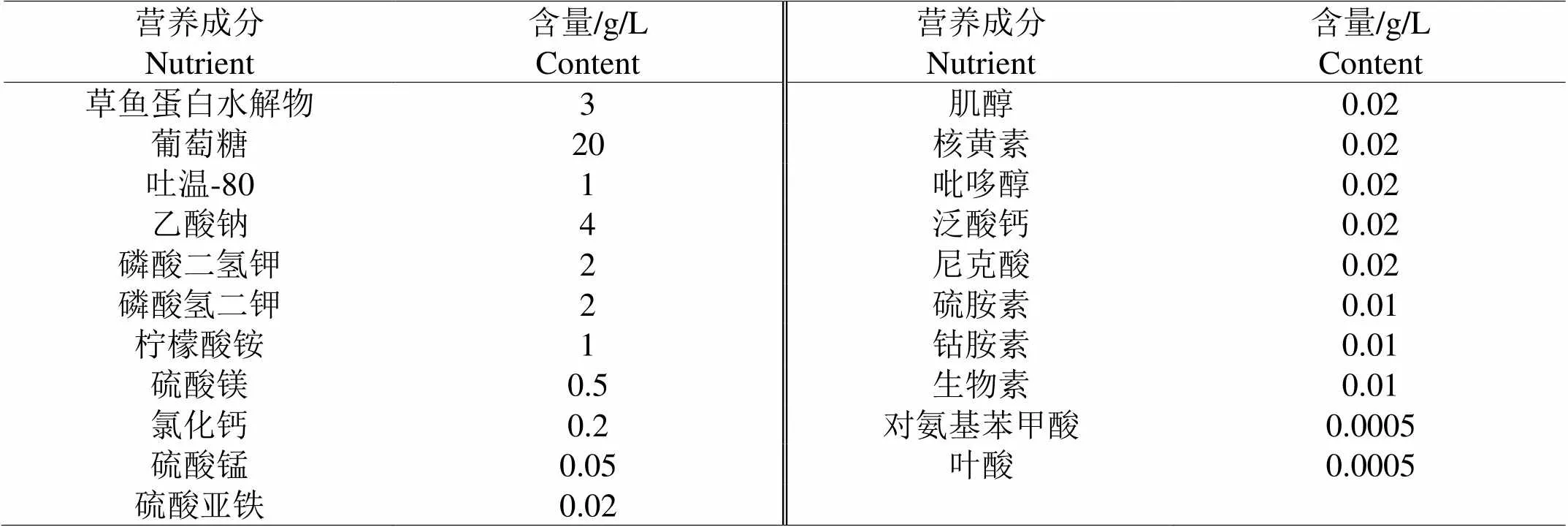
表 1 化学限定培养基基本成分
1.4 酶解产物体外抗氧化活性研究
为进一步了解胶原蛋白肽的特性,进行了体外抗氧化活性测定。
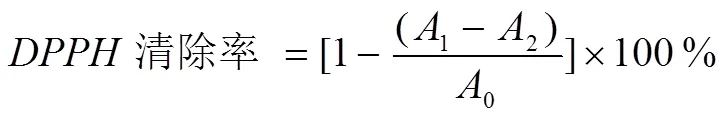
式中:0—2 mL蒸馏水+2 mL DPPH溶液的吸光值;1—2 mL样品溶液+2 mL DPPH溶液的吸光值;2—2 mL样品溶液+2 mL DPPH无水乙醇的吸光值。
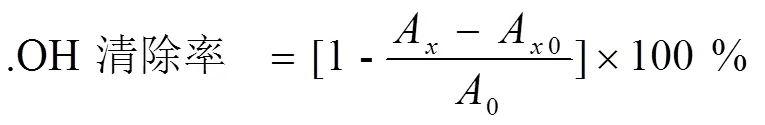
式中:0—为空白对照的吸光度;A—加样品溶液的吸光度;A0—不加显色剂H2O2溶液的本底吸光度。
1.5 实验数据处理
实验数据采用Excel 2016软件处理分析;响应面数据Design Expert 7.1.3软件处理分析。
2 结果与分析
2.1 酶解反应时间对嗜热链球菌增殖作用研究
酶解反应时间对嗜热链球菌增殖作用的影响如图1所示。由图可知,随着反应时间的增加,嗜热链球菌的OD值逐渐增加,然后降低,并在4 h达到最高值。其可能原因是在酶解反应前期,随着时间的增加,更多的鱼皮被酶解为多肽,但当达到一定时间后,大部分草鱼皮已被酶解,同时酶解产物中多肽也会被风味中性蛋白酶酶解,导致多肽含量降低。显著性差异分析表明,在0.05显著水平上,4 h与5 h、3 h、6 h、7 h、2 h和1 h有显著性差异;5 h与3 h无显著性差异;5 h与6 h、7 h、2 h和1 h有显著性差异;3 h与6 h、7 h、2 h和1 h有显著性差异;6 h与7 h、2 h和1 h有显著性差异;7 h与2 h和1 h有显著性差异;2 h和1 h有显著性差异。
2.2 料水比对嗜热链球菌增殖作用研究
料水比对嗜热链球菌增殖作用的影响如图2所示。由图可知,随着加水量的增加,嗜热链球菌的OD值先增加,然后下降,当料水比为1:10时,嗜热链球菌的OD值最高。这可能是因为酶解反应是在水中进行的,料水比数值较低时,高浓度的底物蛋白过于聚集而不利于与风味中性蛋白酶活性部位结合,从而产生较少的多肽;随着加水量的增加,分子运动活跃,风味中性蛋白酶与底物接触机会增多,水解反应充分,生成的多肽的量也增多;当加水量达到一定数值后,酶与草鱼皮的浓度降低,两者过于分散,接触机会减少,导致多肽含量生成量减少。显著性差异分析表明,在0.05显著水平上,料水比(w/v) 1:10与1:5、1:15和1:20有显著性差异;料水比(w/v) 1:5与1:15(w/v)无显著性差异;料水比(w/v) 1:5与1:20有显著性差异;料水比(w/v) 1:15与1:20有显著性差异。
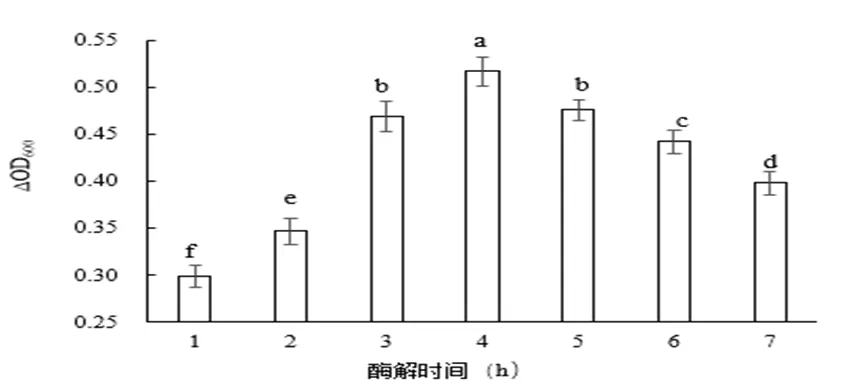
图 1 酶解时间对嗜热链球菌增殖作用的影响

图 2 料水比对嗜热链球菌增殖作用的影响
Fig.2 Effect of ratio of material to water on proliferation of
2.3 加酶量对嗜热链球菌增殖作用研究
加酶量对嗜热链球菌增殖作用的影响如图3所示。由图可知,随着加酶量的增加,嗜热链球菌的OD值逐渐增加,在加酶量为3%时OD值最大,之后随着加酶量的增加而降低。可能原因是当草鱼皮数量一定时,随着加酶量的增加,风味中性蛋白酶与底物蛋白的接触机会增加,水解反应充分,生成的多肽增多,但加酶量继续增大时,过多的酶进入反应体系,导致过度酶解从而使多肽进一步分解为氨基酸,使多肽数量降低。其原因是氨基酸虽也可以被嗜热链球菌生长所利用,但其增殖效果远不如寡肽[12],从而导致嗜热链球菌的OD值降低。显著性分析表明,在0.05显著水平上,加酶量3%与4%无显著性差异;3%与2%、5%和1%有显著性差异;4%与2%、5%和1%有显著性差异;2%与5%和1%有显著性差异;5%与1%有显著性差异。从节约生产成本考虑,将加酶量定为3%。

图 3 加酶量对嗜热链球菌增殖作用的影响
2.4 促进嗜热链球菌增殖酶解工艺的响应面优化
2.4.1 模型建立及其显著性采用Box-Behnken Design(BBD)实验设计方法,对促进嗜热链球菌增殖酶解工艺进行响应面优化,分别对酶解时间,料水比,加酶量三因素在单因素基础上进行三因素三水平试验,以OD600为响应值,结果如表2。
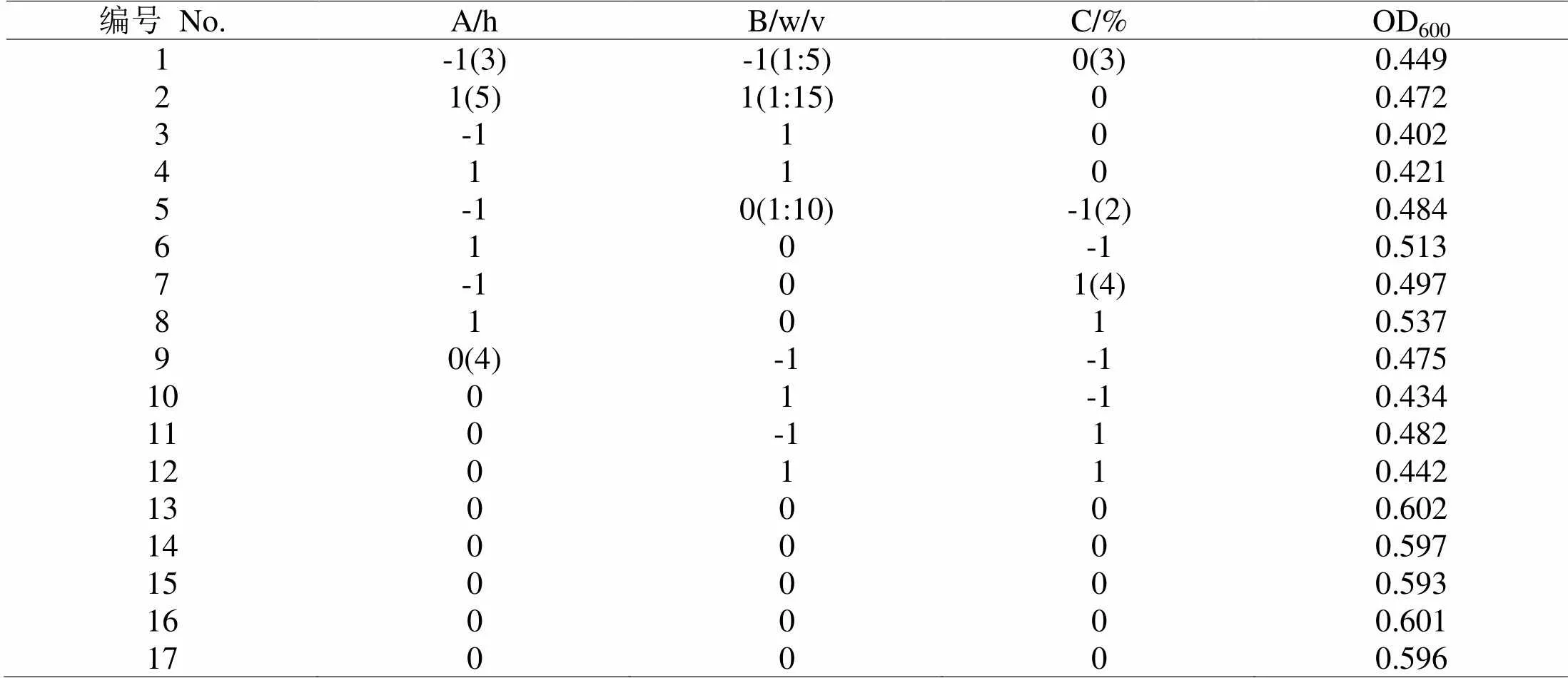
表 2 风味中性蛋白酶响应面分析
利用Design Expert 7.1.3软件,以OD600为响应值,对表2试验数据二次多元回归拟合,得到回归模型方程:
OD600()=0.60+0.014A-0.022B+0.0065 C-0.001AB+0.00275AC+0.00025BC-0.056A2-0.11B2-0.034C2
式中A、B、C依次为酶解时间、料水比(w/v)、加酶量(%)。
通过方差分析对回归方程进行了评价,并对其显著性进行了检验,结果如表5所示。该模型的值为246.22,值小于0.0001,表明所得到的模型极显著,模型误差失拟项中值为4.57,值为0.0882,该模型失拟项不显著即说明该方程对试验拟合情况好,可靠性较高。此外该模型的决定系数(2)为0.9969和调整决定系数(R2)为0.9928,都接近1.0,表示该模型可用于嗜热链球菌增殖酶解工艺实验预测。该模型中,一次项B对嗜热链球菌增殖影响极显著(< 0.01),比较A、B、C3因素F值大小可知,3因素对嗜热链球菌增殖影响大小的顺序为:料水比>酶解时间>加酶量。二项式A2、B2、C2的值均小于0.0001,说明这些因素对嗜热链球菌增殖影响极显著;交互项AB、AC、BC对嗜热链球菌增殖作用影响不显著(< 0.05)。
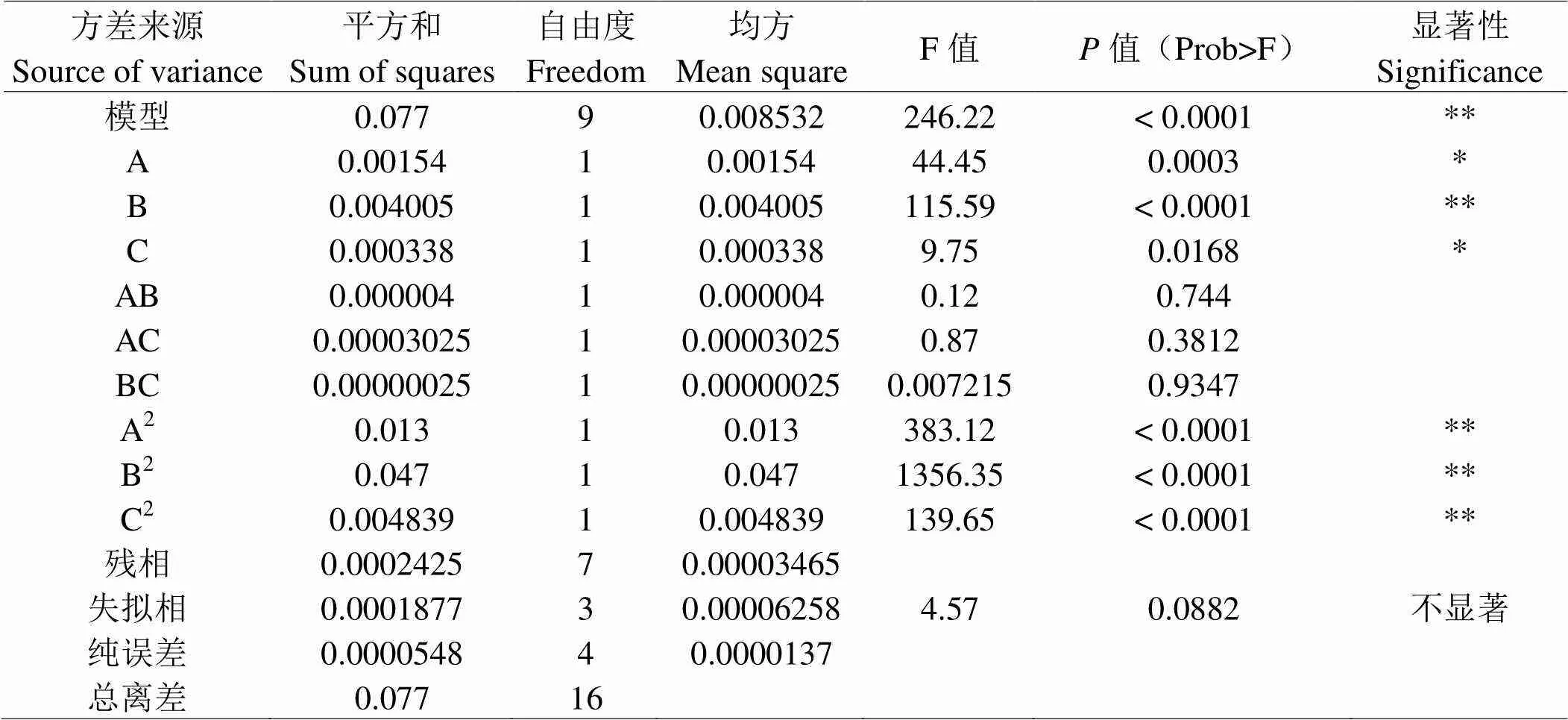
表 3 风味中性蛋白酶响应面实验方差分析
注:**表示极显著(<0.01); *表示显著(<0.05)。
Note: **very significance(<0.01);* Significance(<0.05).
2.4.1 响应面因素交互作用分析优化工艺试验中,酶解时间、料水比、加酶量三因素间的交互作用对嗜热链球菌增殖的影响,如图4。图4(a)、(b)、(c)直观地反映了各因素间的交互影响,可以看出AB、AC、BC交互作用的图形均不陡峭,它们对嗜热链球菌增殖作用影响均不显著。

图 4 各因素交互作用影响嗜热链球菌增殖的响应面图
2.4.3 验证性试验 根据软件分析,得到促进嗜热链球菌增殖最佳条件为:酶解时间4.13 h,料水比1:9.89(w/v),风味中性蛋白酶加酶量3.1%,在此条件下,嗜热链球菌OD600理论值0.600。结合实际,将酶解参数修改为酶解时间4.1 h,料水比1:10(w/v),风味中性蛋白酶加酶量3.1%,在此条件下进行三次重复验证试验,OD值为0.597±0.006,与模型预测值0.601基本一致。
2.5 不同分子量草鱼皮胶原蛋白肽对嗜热链球菌增殖效果
由图5可知,相比于未分段的酶解物,GFSPH Ⅳ更能促进嗜热链球菌生长。推测可能是促进嗜热链球菌生长的多肽大多在分子量小于3000 Da。GFSPH Ⅰ、GFSPH Ⅱ、GFSPH Ⅲ也能促进嗜热链球菌增长,但效果不如GFSPH Ⅳ,这是由于分子量小的肽更容易被嗜热链球菌利用,且分子量越小,对嗜热链球菌增殖效果越明显,这表明分子量小于3000 Da的多肽是嗜热链球菌最佳氮源,结果与Zhao Hongfei等[13]和Juillard V等[18]结果一致。显著性差异分析表明,在0.05显著水平上GFSPH Ⅳ与未分段、GFSPH Ⅲ、GFSPH Ⅱ和GFSPH Ⅰ有显著性差异;未分段与GFSPH Ⅲ、GFSPH Ⅱ和GFSPH Ⅰ有显著性差异;GFSPH Ⅲ与GFSPH Ⅱ无显著性差异;GFSPH Ⅲ与GFSPH Ⅰ有显著性差异;GFSPH Ⅱ与GFSPH Ⅰ无显著性差异。
2.6 GFSPH Ⅳ添加量对嗜热链球菌增殖效果
为分析GFSPH Ⅳ对嗜热链球菌增殖作用,研究了不同GFSPH Ⅳ添加量(0.2 g/L, 0.4 g/L,0.6 g/L,0.8 g/L,1.0 g/L,2.0 g/L,3.0 g/L,4.0 g/L,5.0 g/L)对嗜热链球菌增殖效果。由图6可以看出,GFSPH Ⅳ添加量由0.2~3.0 g/L时,OD值先增加,随后逐渐稳定。因此,结合经济方面考虑,GFSPH Ⅳ最佳添加量为3.0 g/L。
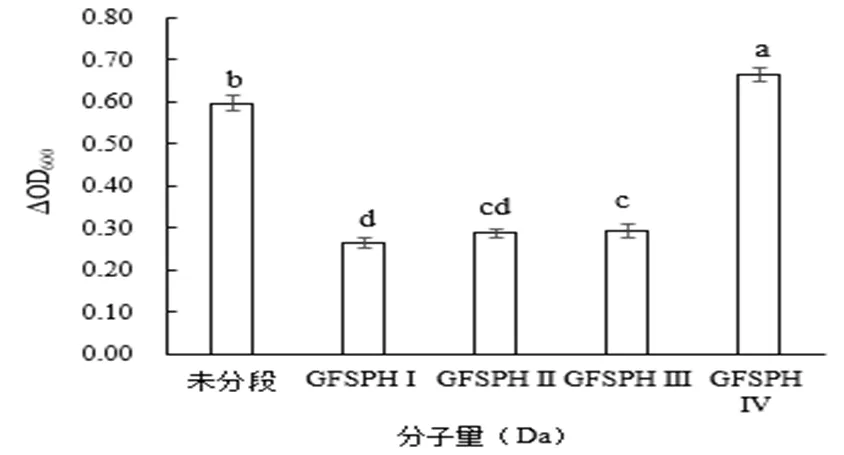
图 5 不同分子量酶解物对嗜热链球菌增殖作用

图 6 GFSPH Ⅳ添加量对嗜热链球菌增殖效果
Fig.6 OD600values ofproliferated by different concentrates of GFSPH Ⅳ
2.7 体外抗氧化性研究
2.7.1 DPPH自由基清除能力不同浓度GFSPH Ⅳ对DPPH自由基清除能力见图7。由图可以看出,GFSPH Ⅳ对DPPH自由基均有较强的清除活性,在拟合范围内,随着浓度的增加,对自由基的清除率呈上升的趋势。对样品浓度与自由基清除率的数据选用二次函数模型进行拟和,可得到拟和方程,以自由基清除率为50%时样品的浓度IC50来衡量样品对DPPH自由基的清除能力。结果可得:=-0.17912+7.0131+11.971,²=0.9956,拟合范围1~10 mg/mL,IC50=6.50 mg/mL。这表明GFSPH Ⅳ具有一定的DPPH清除能力,在浓度为5 mg/mL时,GFSPH Ⅳ DPPH清除能力达到42.71%,远高于张华[19]制备的比目鱼皮多肽的DPPH清除率(15.38%),然而略低于贾韶千[20]制备的黄鳝鱼骨多肽的DPPH清除率(90.85%)。
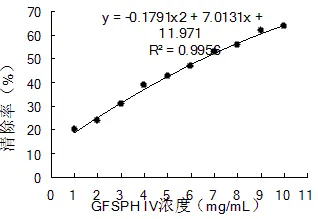
图 7 不同浓度GFSPH Ⅳ的DPPH清除能力

图 8 不同浓度GFSPH Ⅳ的羟自由基清除能力
2.7.1 羟自由基清除能力不同浓度GFSPH Ⅳ对羟自由基清除能力见图8。由图可以看出,GFSPH Ⅳ对自由基均有较强的清除活性,随着浓度的增加,对自由基的清除率呈上升的趋势,当浓度达到一定值时,清除率增加趋向平衡。对样品浓度与自由基清除率的数据选用二次函数模型进行拟和,可得到拟和方程。结果可得:=-0.1142+5.607+17.393,²=0.9854,拟合范围2~20 mg/mL,IC50=6.74 mg/mL。实验结果与张华[19]制备的比目鱼皮多肽的羟自由基清除能力相当,而远高于苏永昌[21]制备的海参多肽IC50(12.5 mg/mL)值。
3 结论
本文对风味中性蛋白酶草鱼皮酶解物对嗜热链球菌增殖效果进行了研究,并对酶解条件进行了优化。利用单因素试验选出对嗜热链球菌增殖较好的酶解时间、料水比和加酶量的最佳单因素水平,进行响应面优化,得到促进嗜热链球菌增殖的最佳酶解工艺为:酶解时间4.13 h,料水比1:9.89(w/v),风味中性蛋白酶加酶量3.1%,验证实验表明该模型与实际结果基本拟合,可较好地预测实验结果。分段实验结果表明,草鱼皮胶原蛋白肽的分子量越小(<3000Da),对嗜热链球菌增殖效果越明显。体外抗氧化性表明,GFSPH Ⅳ对DPPH自由基清除的IC50值为6.50 mg/mL,对羟自由基自由基清除的IC50值为6.74 mg/mL,说明GFSPH Ⅳ对DPPH自由基的清除能力强于羟自由基。GFSPH Ⅳ既可促进益生菌增殖,还具有抗氧化生物活性,可以为多组分功能性食品的开发提供理论依据。
[1] 农业部渔业局.中国渔业年鉴[M].北京:中国农业出版社,2016:30-31
[2] Guo HH, Hong ZA, Yi RZ. Core-Shell Collagen Peptide Chelated Calcium/Calcium Alginate Nanoparticles from Fish Scales for Calcium Supplementation[J]. Journal of Food Science, 2015,80(7):N1595-1601
[3] 刘朝霞,陈海光,黄东雨.鱼皮胶原蛋白的提取及其应用[J].广东农业科学,2011(20):100-102
[4] Ogawa M, Moody MW. Biochemical Properties of black drum and sheep shead seabream Skin collagen[J]. Journal and Agriculture and Food Chemistry, 2003,51(27):8088-8092
[5] 付鸿,李靖靖,岳春.酸奶中杆菌和球菌的快速分离及发酵剂的制备[J].化学与生程,2013,30(8):70-73
[6] 曾丽,李丽,王加斌,等.水产胶原蛋白肽功能活性及其制备工艺研究进展[J].浙江海洋学院学报:自然科学版,2013,32(2):163-168
[7] Liu SQ, Holland R, Crow VL. The potential of dairy lactic acid bacteria to metabolize amino acids via non-transamination reactions and endogenous transamination[J]. International Journal of Food Microbiology, 2003,86:257-269
[8] Aguirre L, Garro MS, Savoy G. Enzymatic hydrolysis of soybean protein using lactic acid bacteria[J]. Food Chemistry, 2008,111(4):976-982
[9] 陈建康,包建强.酶解法制备草鱼皮蛋白水解物增殖嗜热链球菌[J].食品科学,2016,37(5):144-148
[10] Thomas TD. Role of Lactic Acid Bacteria and Their Improvement for Production of Better Fermented Animal Products[J]. New Zealand journal of dairy science and technology, 1985,2:1-10
[11] 支潇,田晓宇,田辉,等.水解大豆蛋白对嗜热链球菌增殖的促进作用[J].食品科技,2012,37(8):13-17
[12] 白凤翎,张柏林,赵宏飞.大豆蛋白水解物促酸奶乳酸菌增殖及生长动力学[J].食品与发酵工业,2012,38(1):51-56
[13] Zhao HF, Bai FL, Zhou F,. Characterization of soybean protein hydrolysates able to promote the proliferation of Streptococcus thermophiles ST[J]. Journal of Food Science,2013,78(4):M575-M581
[14] Li CM, Zhong ZH, Wan QH,. Preparation and thermal stability of collagen from scales of grass carp ( Ctenopharyngodon idellus)[J]. European Food Research and Technology, 2008,227(5):1467-1473
[15] 宋益善,王肖南,秦梅,等.草鱼皮酶解工艺及水解物对嗜热链球菌增殖作用研究[J].科学技术与工程,2016,16(33):249-254
[16] Shu GW, He YX, Lei N,. Cellulase-Assisted Extraction of Polysaccharides from White Hyacinth Bean: Characterization of Antioxidant Activity and Promotion for Probiotics Proliferation[J]. Molecules, 2017,22(10):1-13
[17] 段超,许刚豪,王冬冬,等.茯苓发酵液多糖含量分析及其抗氧化性能研究[J].日用化学品科学,2016,39(2):31-34
[18] Juillard V, Bars DL, Kunji ER,. Oligopeptides are the main source of nitrogen for Lactococcus lactis during growth in milk[J]. Applied and Environmental Microbiology, 1995,61(8):3024-3230
[19] 张华.比目鱼皮抗氧化肽的制备及分离纯化[D].天津:天津大学化工学院,2013:49-50
[20] 贾韶千,李艳霞.黄鳝鱼骨多肽制备及其抗氧化活性[J].食品科学,2016,37(1):133-138
[21] 苏永昌,刘淑集,吴成业.海参多肽的制备工艺优化及其抗氧化测定[J].福建水产,2009,6(2):6-10
Optimization of Hydrolysis Conditions for Peptides from Grass Carp Fish Skin: Promotion forProliferation and Characterization of Antioxidant Activity
GE Jun-miao1, LI Qi-le1, DAI Qi-hu1, CHEN Jian-kang1, JIANG Cai-bei1, ZHANG Lu1, SHENG Jie1,2, SONG Yi-shan1,2,3*
1.201306,2.201306,3.()201306,
The hydrolysates from grass carp fish skin were prepared by using flavor neutral protease, and the response surface methodology (RSM) was employed to optimize the enzymatic hydrolysis conditions, by using proliferation effect ongrowth of hydrolysate as the evaluation index. The results showed that the optimum conditions were hydrolysis time of 4.13 h, the ratio of water to raw material of 1:10 (w/v), enzyme concentration of 3.1%. The Grass Carp fish skin hydrolysate prepared under the optimum conditions was fractionated to four fragments according to molecular weight sizes: >10000 Da;5000~10000 Da;3000~5000 Da 0;< 3000 Da, in which the fragments with molecular weights of less than 3000 Da significantly radicals (2,2-diphenyl-1-picrylhydrazyl (DPPH) > superoxide > hydroxyl).
; flavor neutral protease; polypeptides; Response Surface Methodology; antioxidant activity
TS254.9
A
1000-2324(2018)06-0968-07
10.3969/j.issn.1000-2324.2018.06.013
2018-06-05
2018-07-13
上海海洋大学科技发展专项基金(2018)资助(A2-0203-00-100229)
葛俊苗(1995-),女,在读研究生,主要研究方向为功能性食品. E-mail:947852513@qq.com
Author for correspondence. E-mail:yssong@shou.edu.cn
猜你喜欢
化工管理(2022年14期)2022-12-02
当代水产(2022年7期)2022-09-20
今日农业(2022年4期)2022-06-01
石油沥青(2022年2期)2022-05-23
当代水产(2022年3期)2022-04-26
纺织科学研究(2021年7期)2021-08-14
当代水产(2019年9期)2019-10-08
广东第二课堂·小学(2018年9期)2018-10-24
天然产物研究与开发(2018年7期)2018-08-21
分析化学(2017年12期)2017-12-25
