
草坪式屋顶绿化中4种景天科植物的抗旱性
2019-01-03 02:05吴小青陈思逸曹丹周郑坤朱明库
江苏农业科学 2019年21期
吴小青 陈思逸 曹丹 周郑坤 朱明库


摘要:景天科植物因具备栽培简便、繁殖容易且抗旱抗寒性强等特点而成为城市屋顶绿化植物的首要选择。为筛选适宜大面积推广的轻型屋顶绿化植物,以形态相近的4种景天科植物为材料,探究其在聚乙二醇6000(PEG-6000)模拟干旱条件下丙二醛(MDA)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)以及脯氨酸(Pro)含量的变化情况。结果表明:在干旱胁迫下,抗旱性较强的景天科品种表现出Pro含量和CAT活性均较高,MDA含量较低,而SOD活性与景天科植物抗旱性的相关性相对较弱。综合分析,4种景天科植物的抗旱能力从强到弱依次表现为凹叶景天>丸叶景天>圆叶景天>胭脂红景天,这些结果为日后屋顶绿化植物的筛选和种植提供科学依据和参考。
关键词:屋顶绿化;景天科植物;干旱胁迫
中图分类号: S688.401文献标志码: A
文章编号:1002-1302(2019)21-0210-04
收稿日期:2018-08-08
基金项目:国家自然科学基金(编号:31700226);住房与城乡建设部科学技术计划(编码:2016-K1-028)。
作者简介:吴小青(1984—),女,陕西南郑人,硕士,讲师,主要从事园林绿化与植物配置等研究。E-mail:wxq8451@163.com。
通信作者:朱明库,博士,硕士生导师,主要从事植物逆境生物学研究。E-mail:mingkuzhu007@126.com。
屋顶绿化作为一种不占用宝贵土地资源的绿化形式,在人口密集的城市中越来越受到重视。屋顶环境具有光照强、干燥、土层薄等特点,因此应尽量选择耐旱抗寒性强的矮小灌木和草本植物。根据植物的选择和种植特点,屋顶绿化又分为多种类型,其中草坪式屋顶绿化对屋顶承重要求极低,且种植成本低,养护难度小,具有广阔的应用前景,但由于受植物种类范围小的影响,在实践中推广受到限制。景天科(Crassulaceae)植物具有根系浅、植株低矮、生长整齐、耐旱、耐寒、耐贫瘠、抗风性强等特点,且采用分株、扦插和播种法繁殖均可,是草坪式屋顶绿化的首选[1]。
干旱会对植物的生理指标和新陈代谢等方面造成不利影响。具体表现为,当植物蒸腾的水分得不到补偿时,原生质的运动、结构、弹性等就会受到损害,膜上脂层分子排列破坏,细胞透性增加,同时植物正常的生理过程被破坏,合成受到抑制,分解加速,导致植物生长减弱,叶片失水、衰老,最终死亡[2]。目前,有关景天科植物在干旱条件下的抗逆性研究依然较少。尽管适应性极强的景天科多年生肉质草本植物佛甲草已得到广泛运用[3],但长期的单一植物种植也带来很多问题,例如虫害、病害、观赏性差等[4]。因此,研究景天植物干旱条件下的生理指标变化,可以反映出景天科植物的耐干旱程度以及干旱对不同品种的影响程度等,这将为草坪式屋顶绿化中的耐旱品种筛选和灌溉提供科学指导。
为了丰富草坪式屋顶绿化景观,扩大品种选择范围,本研究挑选了4种适宜徐州地区气候条件,且形态相似、耐旱性较强的景天属植物作为研究对象。聚乙二醇6000(PEG-6000)可调节渗透势,且因其分子量较大,不会穿越细胞壁进入细胞质,引起质壁分离,给植物造成伤害,已广泛用于模拟干旱胁迫[5-6]。本研究通过浇灌30% PEG-6000来研究这4种景天属植物对干旱胁迫的生理生化反应及调节适应能力,为今后屋顶绿化植物的筛选和种植提供科学依据。
1材料与方法
1.1试验材料
供试材料为丸叶景天、凹叶景天、圆叶景天和胭脂红景天,均通过扦插繁殖于江苏师范大学植物智能温室。
1.2试验设计
试验于2017年3月在江苏师范大学温室中进行,将生长状况良好的4种品种扦插于穴盘中,穴盘平底有网孔(长× 宽=42 cm×42 cm),种植基质采用泥炭 ∶田园土 ∶通气型珍珠岩(体积比)=2 ∶1 ∶1,厚度为4 cm。每盘扦插35株,每种2盘。植株生长稳定后移栽至花盆中培养,培养条件为:温度(25±2) ℃,湿度(50±5)%,平时进行正常养护和管理。1个月后选取生长状况良好、长势相近的植株用于干旱胁迫试验。干旱胁迫通过浇灌30% PEG-6000进行,每隔3 d浇灌1次,每次每株50 mL,于浇灌后0、8、18 d拍照记录植物萎蔫表型的变化,并于浇灌后0、8、12、18、24 d取样测定胁迫相关生理指标变化。
1.3指标测定与数据处理
丙二醛(MDA)含量、过氧化氢酶(CAT)活性、超氧化物歧化酶(SOD)活性、脯氨酸(PRO)含量的测定为4次生物重复的结果,均通过南京建成生物工程研究所(http://www.njjcbio.com/)销售的检测试剂盒进行。
采用IBM SPSS Statistics 20软件进行数据分析,并通过Origin 8.0进行作图。
2结果与分析
2.1干旱胁迫期间4种景天科植物的表型变化
干旱胁迫开始前,筛选生长一致、大小相似的4种景天科植物用于胁迫试验(图1-A)。干旱胁迫5 d后,胭脂红景天即出现萎蔫迹象,且6 d后萎蔫现象十分明显(结果未展示)。干旱胁迫8 d后,圆叶景天叶子开始变软,颜色变暗;丸叶景天和凹叶景天暂时变化不大。而此时胭脂红景天叶片几乎完全掉落(图1-B),这说明胭脂红景天的抗旱性最差,因此此后的表型记录及生理指标测定将不再包含胭脂红景天。干旱胁迫9 d后,圆叶景天出现明显萎蔫(结果未展示)。干旱胁迫12 d后,圆叶景天叶子颜色暗化明显,丸叶景天出现萎蔫迹象(结果未展示)。干旱胁迫18 d后,凹叶景天才开始出现轻微的萎蔫现象,而此时圆叶景天和丸叶景天枯萎性狀更为显著,这说明凹叶景天的抗旱性最强(图1-C)。
2.2干旱胁迫对叶片丙二醛(MDA)含量的影响
MDA是膜脂过氧化作用的主要产物之一,其含量反映着细胞膜脂过氧化作用和质膜被破坏程度[7]。通常来说,当植物受干旱胁迫时,其质膜受到不同程度的破坏,MDA含量增高。此外,在受到相同条件的干旱胁迫时,不同植物的质膜受到不同程度的破坏,MDA含量也有所不同。含量越高,组织伤害越严重,说明抗旱性越弱;反之,积累含量越低,组织伤害相对越轻,说明抗旱性越强[2,8]。
如图2所示,干旱胁迫能够显著诱发景天科植物中MDA含量的增加,总体表现出“升—降—升”的变化趋势。在干旱胁迫处理下,胭脂红景天MDA含量增幅变化最大,其次为圆叶景天,说明干旱能够严重破坏胭脂红景天质膜。丸叶景天和凹叶景天中MDA含量变化增加程度较小且含量相似。胁迫初期,MDA含量上升的原因可能是植物尚未感受到干旱胁迫,因此未启动膜质过氧化防御系统,之后防御系统启动导致MDA含量下降,胁迫末期由于保护酶的消耗MDA含量再次上升[9]。以上結果表明,凹叶景天和丸叶景天在干旱胁迫下MDA含量变化较小,抗旱性较强,其余依次为圆叶景天和胭脂红景天。
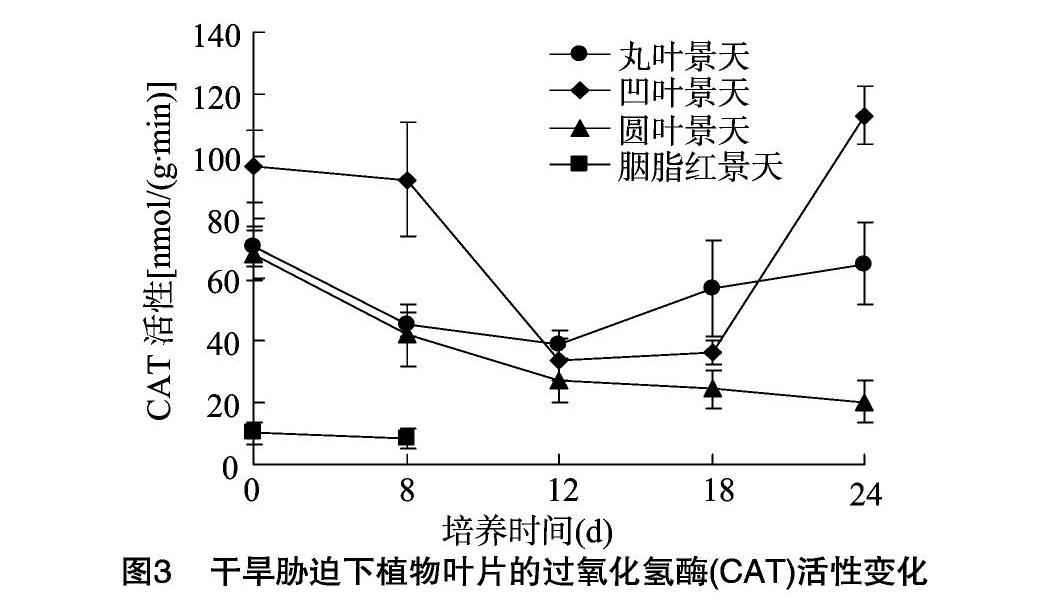
2.3干旱胁迫对叶片过氧化氢酶(CAT)活性的影响
CAT广泛存在于动物、植物、微生物和细胞中,是最主要的H2O2清除酶,在活性氧清除系统中具有重要作用。当植物受到环境胁迫时,生物体内自由基增多,CAT与SOD、POD共同组成了生物体内的活性氧防御系统[10]。
如图3所示,在受到干旱胁迫时,凹叶景天和丸叶景天叶片内CAT活性变化基本趋势为先降后升,并在12 d时均达到最低值。这可能是因为植物尚未感受到明显的伤害,因此没有启动自身防御系统。之后持续上升,说明此时植物已感知到自身受到干旱胁迫,须要增加CAT活性来保护细胞膜系统[11]。二者相比,凹叶景天叶片中CAT活性更高,说明其耐旱性更强。圆叶景天叶片内CAT活性变化呈逐渐下降趋势,说明其H2O2清除能力减弱,因而抵御干旱胁迫的能力相应减弱。而相同条件下,胭脂红景天叶片中CAT活性最低,说明其干旱抗性最差,这与干旱胁迫下其表型变化特征相符。以上结果表明,干旱胁迫下凹叶景天CAT活性增加最为明显,防御抵抗系统最强,抗旱性最强,其次依次为丸叶景天、圆叶景天、胭脂红景天。
2.4干旱胁迫对叶片超氧化物歧化酶(SOD)活性的影响
细胞受到干旱胁迫时,胞内活性氧产生和清除的动态平衡被打破,细胞膜受到损伤,此时,超氧化物歧化酶能有效清除超氧阴离子自由基,对细胞进行防御[12]。因此,SOD活性变化能在一定程度上反映植物的抗旱性。
如图4所示,景天科植物叶片中的SOD活性在干旱胁迫下总体上呈现先降后升的趋势。干旱胁迫初期时,植物尚未感受到明显的伤害,因此没有启动自身防御系统,之后植株受干旱胁迫加重时SOD活性随即上升。在未受干旱胁迫时,凹叶景天和圆叶景天中SOD活性较高,而胁迫处理24 d后,丸叶景天中SOD活性最高,但是三者的差异并不显著。尽管胭脂红景天在未受干旱胁迫和胁迫处理8 d后也表现出较高的SOD活性,但是其抗旱性仍是最弱的。以上结果表明干旱胁迫下,景天科植物中SOD活性的变化比较接近。
2.5干旱胁迫对叶片脯氨酸(Pro)含量的影响
作为一种理想的渗透调节物质,植物体内积累Pro含量的高低及增幅的大小可在一定程度上反映植物的抗旱性[13]。
如图5所示,景天科植物叶片中的Pro含量在干旱胁迫下总体上呈现先升后降的趋势。当叶片持续受到干旱胁迫时,Pro含量下降,可能与植株受到严重伤害时Pro的合成能力受到影响有关[14]。其中,凹叶景天在胁迫处理下Pro含量的增加程度保持的最为稳定,说明其通过增加Pro含量来增强细胞的持水力,继而维持细胞正常代谢的能力更强。在胁迫处理12 d时,丸叶景天中Pro含量大幅上升,但与凹叶景天上升的程度没有差异。圆叶景天受到干旱胁迫后Pro含量一直处于下降趋势,可能是圆叶景天已经受到比较严重的伤害,Pro合成能力受到影响,无法继续高效合成。以上结果表明干旱胁迫下,凹叶景天和丸叶景天中Pro含量较高,其次依次为圆叶景天和胭脂红景天。
3讨论与结论
目前对景天科植物的抗旱性研究大多采用了盆栽控水方法,该方法的优点是便于控制浇水量和浇水时间[15]。然而由于不同植物的植株大小、叶片数量、根系发育不同,导致植物吸收和蒸发水分的能力也有所差距,因此,控水法难以保证一批试验材料中每盆土壤的含水量一致,这导致前人的研究中出现矛盾的研究结论。例如,张寅媛等采用盆栽控水法研究干旱胁迫对景天科植物生理生化指标的影响时,发现堪察加景天的抗旱性大于德国景天[16];而段锦兰等从相似的研究中发现,德国景天的抗旱性强于堪察加景天,而试验采用的方法也是盆栽控水法[17]。类似的矛盾还出现在吴永华等的研究[18-19]中。PEG-6000可调节渗透势,并保证试验材料受到同样的渗透胁迫,目前已广泛用于模拟干旱胁迫[6]。因此,本试验采用PEG-6000高渗溶液法进行干旱胁迫处理,相比盆栽控水法,更具便捷性、精确性和科学性[20]。
植物在面临干旱胁迫时可以通过多种途径来抵抗和防御干旱[21]。目前,鉴定植物抗性的指标主要有形态结构、生长产量和生理生化等[22]。干旱胁迫下,抗旱性越弱的植物受干旱胁迫伤害程度越高,叶片越早失水、枯黄,因而,最终整体呈现最先萎蔫的状态,反之,抗旱性越强[23]。本研究发现干旱胁迫时胭脂红景天最先受到伤害,在其余3种植物依然青绿时,其叶片最先脱落,以致后续无法采样,因此胭脂红景天抗旱性最弱。圆叶景天和丸叶景天抗旱性一般,而凹叶景天无论是生长状况还是各项生理指标的测定都优于其他3种,因此其抗旱性最强。
本研究中鉴定抗旱性采用的生理生化指标包括丙二醛(MDA)含量、过氧化氢酶(CAT)活性、超氧化物歧化酶(SOD)活性和脯氨酸(Pro)含量。膜系统通常被认为是干旱伤害的最初和最关键部位。植物受到干旱胁迫时,细胞膜会发生不同程度的破坏,膜脂系统受到影响发生膜质过氧化作用,进而膜透性增大。丙二醛(MDA)作为其产物之一,常被用来作为膜质过氧化指标。本研究中4种植物随干旱胁迫程度加剧,MDA含量呈现“升—降—升”的变化趋势,除胭脂红景天外,其他3种植物的MDA含量总体较低。胭脂红景天增幅最大,说明其膜脂抗氧化能力最弱,细胞膜破坏程度最大,抗旱性最弱。凹叶景天和丸叶景天MDA含量较低,说明干旱胁迫对其细胞膜伤害程度较轻。另外,Pro作为理想的渗透调节物质,当植物受到干旱胁迫时,可作为抗逆性筛选的指标。通常来说,抗逆性强的植株在受到干旱胁迫时往往积累较多的Pro,增幅较大[24]。本研究中4种植物的Pro含量都受干旱胁迫的显著诱导,但总体而言凹叶景天和丸叶景天在干旱胁迫下Pro含量的增加程度相对稳定。
当植物遇到逆境时,会产生大量H2O2,H2O2启动膜脂过氧化或膜脂脱脂作用,进而破坏膜结构,膜差别透性消失,导致一系列有害的生理生化变化,给植物体造成严重的损伤[25]。SOD是重要的抗氧化酶,能有效清除超氧阴离子自由基,减轻膜质过氧化导致的膜伤害。本研究发现,随着干旱胁迫程度加剧,SOD活性呈现先降后升的趋势。这说明初期下降可能是植物尚未感受到干旱胁迫,因此未启动自身防御系统,当干旱胁迫加重后,SOD活性上升,从而提高自身抗干旱能力。本研究中,除胭脂红景天外,其他3种景天科植物的SOD活性差异并不显著。此外,在活性氧清除系统中,CAT作为清除活性氧的膜保护酶类,也可将活性氧转变为低活性物质,从而保护细胞膜系统[26]。本研究中凹叶景天CAT活性的增加程度最大,说明其抵抗胁迫能力最强,其余依次为丸叶景天、圆叶景天、胭脂红景天。
結合4种景天科植物在干旱胁迫过程中的形态变化及其相应的MDA含量、CAT活性、SOD活性和Pro含量的变化,由此可以得出4种景天科植物的抗旱性强弱顺序,为凹叶景天>丸叶景天>圆叶景天>胭脂红景天,该结论可为日后屋顶绿化植物的筛选和种植提供科学依据和参考。
参考文献:
[1]高小燕. 几种景天科植物抗旱性及其生理机制的研究[D]. 呼和浩特:内蒙古农业大学,2009.
[2]宋海鹏,刘君,李秀玲,等. 干旱胁迫对5种景天属植物生理指标的影响[J]. 草业科学,2010,27(1):11-15.
[3]刘爱荣,张远兵,谭志静,等. 模拟干旱对佛甲草生长和渗透调节物质积累的影响[J]. 草业学报,2012,21(3):156-162.
[4]白淑媛,梁芳. 客观看待佛甲草在屋顶绿化中的作用[J]. 北京园林,2005(1):28-31.
[5]Attree S M,Fowke L C. Embryogeny of gymnosperms:advances in synthetic seed technology of conifers[J]. Plant Cell Tissue & Organ Culture,1993,35(1):1-35.
[6]郝曦煜,王红丹,尹智超,等. PEG胁迫对小豆苗期抗旱生理指标的影响及抗旱鉴定体系建立[J]. 作物杂志,2017(4):134-142.
[7]王娟,李德全,谷令坤. 不同抗旱性玉米幼苗根系抗氧化系统对水分胁迫的反应[J]. 西北植物学报,2002,22(2):285-290.
[8]许桂芳. PEG胁迫对2种过路黄抗性生理生化指标的影响[J]. 草业学报,2008,17(1):66-70.
[9]周媛,徐冬云,董艳芳,等. 9种轻型屋顶绿化景天属植物的抗旱性研究[J]. 中国农学通报,2012,28(25):294-301.
[10]张坤生,田荟琳. 过氧化氢酶的功能及研究[J]. 食品科技,2007,32(1):8-11.
[11]林植芳,李双顺,林桂珠,等. 水稻叶片的衰老与超氧物歧化酶活性及脂质过氧化作用的关系[J]. 植物学报,1984(6):605-615.
[12]Dhindsa R S,Matowe W. Drought tolerance in two mosses:correlated with enzymatic defence against lipid peroxidation[J]. Journal of Experimental Botany,1981,32(1):79-91.
[13]Hong Y,Zhang H,Huang L,et al. Overexpression of a stress-Responsive NAC transcription factor gene ONAC022 improves drought and salt tolerance in rice[J]. Frontiers in Plant Science,2016,7(e0116646):4.
[14]杨虎臣. 干旱胁迫对幼苗期甜菜脯氨酸(Pro)代谢通路的影响[D]. 哈尔滨:哈尔滨工业大学,2016.
[15]彭民贵. PEG-6000模拟干旱胁迫下紫斑牡丹的生理响应及其抗旱性研究[D]. 兰州:西北师范大学,2014.
[16]张寅媛,刘英,白龙. 干旱胁迫对4种景天科植物生理生化指标的影响[J]. 草业科学,2014,31(4):724-731.
[17]段锦兰,付宝春,康红梅,等. 10种景天属植物的抗旱性评价[J]. 山西农业科学,2013,41(6):557-560.
[18]吴永华,许宏刚,廖伟彪,等. 5种景天植物抗旱性比较研究[J]. 甘肃林业科技,2009,34(3):15-17,26.
[19]布凤琴,张闽,燕坤蛟. 济南轻型屋顶绿化七种景天类植物的适应性研究[J]. 山东建筑大学学报,2011,26(6):551-555.
[20]杨强胜,唐晓蓉,邬晓红,等. 林木耐旱技术研究进展[J]. 北方农业学报,2006(5):26-28.
[21]倪郁,李唯. 作物抗旱机制及其指标的研究进展与现状[J]. 甘肃农业大学学报,2001,36(1):14-22.
[22]李瑞雪,孙任洁,汪泰初,等. 植物抗旱性鉴定评价方法及抗旱机制研究进展[J]. 生物技术通报,2017,33(7):40-48.
[23]刘惠芬,史铭均,高玉葆. 不同种群羊草幼苗对渗透胁迫的反应和生理生态适应[J]. 南开大学学报(自然科学版),2003,36(2):31-36.
[24]张博文,李富平,许永利,等. PEG-6000模拟干旱胁迫下五种草本植物的抗旱性[J]. 分子植物育种,2017(8):1-12.
[25]陆小静,张振文,罗秀芹,等. 木薯耐旱基因CAT的生物信息学预测与分析[J]. 西南农业学报,2013,26(6):2214-2220.
[26]张武. 马铃薯叶绿素含量、CAT活性与品种抗旱性关系的研究[J]. 农业现代化研究,2007,28(5):622-624.
猜你喜欢
山东农业科学(2016年11期)2016-12-17
科技资讯(2016年18期)2016-11-15
现代园艺(2016年17期)2016-10-17
现代园艺(2016年2期)2016-03-15
