
农业景观组成对玉米螟天敌数量的影响*
2019-01-03 03:14:44边振兴龚玲春果晓玉
中国生态农业学报(中英文) 2019年1期
边振兴, 龚玲春, 果晓玉, 于 淼
农业景观组成对玉米螟天敌数量的影响*
边振兴1, 龚玲春1, 果晓玉1, 于 淼2**
(1. 沈阳农业大学土地与环境学院 沈阳 110161; 2. 沈阳农业大学理学院 沈阳 110161)
高比例和多样的非耕作生境带来的农田景观高异质性是维持农业生物多样性的关键, 推测提高非耕作生境组成能够促进农田中玉米螟天敌数量和种类增加。为揭示不同非耕作生境组成对玉米螟天敌数量的影响, 本文选取东北平原玉米种植典型县——辽宁省昌图县, 按照5个非耕作生境比例(0~10%、10%~20%、20%~30%、30%~40%和40%~50%)选取20个直径为600 m的样区。陷阱法采集玉米螟天敌, 并用体视显微镜鉴别到科。主成分分析法筛选主要非耕作生境类型, 逐步回归模型对非耕作生境组成(比例和类型)与玉米螟天敌数量进行拟合。并采用“赤池信息准则(AIC)”模型筛选, 评估不同非耕作生境组成对玉米螟天敌数量的影响。研究结果表明, 随着非耕作生境比例增加, 玉米螟天敌数量呈先增后减的趋势, 非耕作生境比例为20%~30%时天敌数量达最大值。非耕作生境比例20%~30%时, 非耕作生境组成与玉米螟天敌数量关系最优模型(AIC=4.24, 为最小值)是=0-1+2PC2, 最优模型表明PC2(=3.787,=0.005)代表以林地为主的非耕作生境组成与玉米螟天敌数量呈显著正相关, 景观多样性指数(=-3.173,=0.005)与玉米螟天敌数量呈显著负相关。该比例的非耕作生境组成与玉米螟天敌数量关系的其他模型表明, 以草地和村落为主(PC1)的非耕作生境与玉米螟天敌呈显著正相关(=1.957,=0.000)。而其他比例下, 所有关系模型说明农田周边林地生境和其他作物耕地有利于提高农田中天敌数量。最适宜玉米螟天敌聚集的非耕作生境比例为20%~30%, 以林地为主或村落、草地为主的非耕作生境, 以及分布有其他作物的耕作生境能够提高玉米螟天敌数量。研究结果可为今后从景观生态角度构建适宜非耕作生境组成来防治玉米螟提供理论依据和方法。
农业景观; 非耕作生境; 玉米田; 玉米螟; 天敌
集约化越来越成为现代农业生产的主要方式, 这种集约化生产导致农田景观结构单一, 农田内生物多样性减少[1-2], 而且改变了昆虫间的关系, 影响了天敌对害虫的防治作用[3-4]。集约化的农田管理生产措施导致农业生态系统发生了较大变化。农业景观是由作物生境和非耕作生境组成的[5], 由农田边缘草地和林地等组成的非耕作生境[3]为当地害虫天敌提供栖息地和食物源等[6-9], 在农业生态系统资源缺乏时, 非耕作生境作为天敌的补充食物源及避难的场所[10-11], 当耕作生境中作物能为天敌提供充足的食物源时, 天敌则从非耕作生境向耕作生境中相互转移扩散, 这种扩散方式增加了害虫天敌在农田生境中的聚集, 显著提高了农田生境中天敌的丰富度[12]。非耕作生境还可以避免因杀虫剂的使用[13]、作物管理措施等因素带来的人为干扰产生的影响, 有利于农田生境天敌多样性的维持和保护[14-16]。非耕作生境是多种植物、动物等生物的生活储备库[3], 为天敌提供必要的食物来源和栖息地, 提高了天敌丰富度, 例如天敌步甲(Carabidae)[17-18]、蟹蛛(Thomisidae)[12,19-20]、蜘蛛(Aranelida)[21]、瓢虫(Coccinellidae)[22]等。大量研究证明农田周边非耕作生境比例显著提高了农田中天敌多样性[23]。有关研究认为, 农田景观中非作物生境比例达20%时最能保护生物多样性和相联系的生态系统, 有利于生物害虫防治[2]。国际生物防治组织IOBC建议农田生态系统中存在不低于5%的半自然生境用地[2], 当非耕作生境比例超过15%时能较好地发挥非耕作生境对天敌的保护作用, 从而实现农业景观中天敌的生物防治效果[5]。但这种效用具有局限性, 当非耕作生境比例高于或低于某一值时, 天敌数量随之减少。近年来, 非耕作生境构成受到越来越多的重视[3,24]。Bianchi等[3]从农业生态系统结构对自然天敌保护和害虫控制的影响研究, 着重论述了非耕作生境组成以及作物与非作物生境组合对天敌丰富度保护具有至关重要的作用[3]。刘雨芳等[25]认为增加农田景观内的生境类型能提高农田内的生物多样性, 从而降低害虫密度。Hawro等[26]认为景观结构越复杂, 害虫密度越低, 寄生性天敌数量越高。复杂的农业景观结构不仅能提高自然天敌的数量, 还有效降低了害虫密度[27]。国内外很多研究者从农业景观格局对步甲、蜘蛛等天敌的影响展开了研究, 如, 张旭珠等[28]在华北地区农田中通过陷阱法分析农田边缘不同植被及相邻农田对步甲和蜘蛛多样性的影响, 结果表明边界较高的草本层盖度增加了步甲和皿蛛科(Linyphiidae)的多样性; 赵爽等[29]采用陷阱法从不同尺度上分析景观要素和环境要素对林地生境中蜘蛛多样性的影响, 结果表明, 100 m尺度内夏季林地中植被高度和盖度对蜘蛛多样性影响最大, 草本植被对蜘蛛多样性影响极为显著; Zhou等[30]通过研究农田景观格局以及农药的使用对棉田瓢虫发生的影响, 发现农田景观系统中作物玉米()和草地生境有利于瓢虫在麦田聚集的发生; Dong等[31]研究农田景观系统中林带对瓢虫多样性的影响, 发现农田生态系统大面积的林地有利于瓢虫迁入麦田。但关于农业景观组成对玉米螟()害虫天敌控害效果研究较少。玉米螟作为玉米作物的重要害虫之一, 对其防治具有重要意义。
东北平原是我国重要的粮食主产区, 昌图县地处东北平原南部, 是全国著名的农业大县, 也是重要的粮食生产基地。该县玉米种植面积约占耕地面积的56%以上, 以农业景观为主。对农业景观中非耕作生境进行研究, 有助于维持农田中天敌物种多样性, 通过景观途径实现病虫害防治, 保护农业生态环境, 从而减少农药的使用。本研究以昌图县为研究区, 探讨粮食主产县不同非耕作生境比例、景观组成与天敌数量之间的关系, 为未来耕地生态型整治和生态农业建设提供有效技术手段, 同时为农田景观生态规划设计提供科学依据。
1 研究区概况与研究方法
1.1 研究区概况
昌图县位于辽宁省最北部, 东北平原南端, 辽、吉、蒙三省(区)交界处, 北部与吉林省的四平市、梨树县、双辽县相邻, 西部隔辽河与沈阳市的法库县、康平县, 内蒙古的科左后旗相望。地处123°32′~ 124°26′E, 42°33′~43°29′N, 总面积4 317 km2。主要作物为玉米, 是典型的玉米种植区, 耕地面积2 666.67 km2, 其中玉米种植面积为1 500 km2, 占县域面积的34.75%, 占耕地面积的56.25%。昌图县地貌自东向西由低山丘陵向辽河平原过渡, 主要分布的土壤类型为暗棕壤、黑土或草甸土、风沙土。属于中温带亚湿润大陆性季风气候, 日照充足, 四季分明, 雨热同季。
1.2 研究方法
1.2.1 采样点设置
结合研究区遥感影像图与土地利用现状图, 划分1 km×1 km样方, 并依据不同非耕作生境比例和农药使用情况选取20个样方(图1), 将样方的非耕作生境比例分为0~10%、10%~20%、20%~30%、30%~40%和40%~50% 5类。上述样方地势平坦, 主要作物为玉米, 农田景观配置相似(避免景观构成对生物多样性的影响), 农田种植模式一致, 且没有施用农药, 不放赤眼蜂。
1.2.2 采样方法
采样时间为2017年6月22—30日(夏季)和9月22日—10月1日(秋季)。在上述20个样方内每样方选取农田周围景观异质性相近的2个半径为300 m的采样区, 夏、秋季用同样的方法采样, 共40个采样区。采用陷阱法采集节肢动物。按平行线法布设陷阱, 从玉米田块边缘20 m起沿垄向田内布设陷阱, 每行设置4个, 纵向间距10 m, 横向间距30 m, 共布设8个(图2)。依此法布设, 共布设320个陷阱。
陷阱具体布设方法为: 将直径7 cm、高度17 cm、容积500 mL的PP塑料杯放入土壤中, 倒入150~200 mL的乙二醇水溶液和洗洁精混合液, 塑料杯杯盖在3个方向用铁丝固定插入土壤里, 杯盖上方距地面约1 cm, 放置7 d后收回。将各陷阱中捕获的节肢动物按对应样本编号放入PE瓶中, 倒入10 mL酒精(75%)。利用体式显微镜(ZEIS:Stemi 2000-C), 并参照《中国土壤动物检索图鉴》、《昆虫分类》等分类系统及相关资料对节肢动物样品进行鉴别, 鉴定其所属门、纲、目和科。并分析样点周边的木本和草本植物的种类及长势。
1.2.3 农业景观格局分析
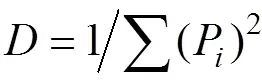
1.2.4 数据分析
采用统计分析中单因素Duncan方法对不同非耕作生境比例下的天敌数量进行比较分析, 并对上述6大类景观变量进行主成分分析, 一般选择累积贡献率>80%的主成分, 根据主成分分析结果, 保留前2或3个主成分, 即主成分1(PC1)、主成分2(PC2)和主成分3(PC3)。采用多模型推理方法, 基于赤池信息量准则(AIC)[33]的多模型推理, 比较PC1、PC2、PC3、及其组合作用对天敌群落数量的影响。并通过多重线性回归模型分析解释变量和响应变量之间的关系。基于已定义的模型集合, 在每个非耕作生境面积比例(0~10%、10%~20%、20%~30%、30%~40%和40%~50%), 用线性指示变量来完成模型拟合程度。采用赤池信息准则(AIC)评估所有模型, 根据AIC值最小, 模型最好的原则优先选出最好的模型进行分析。以上分析通过软件SPSS 18.0完成。赤池信息准则(AIC)表示为:
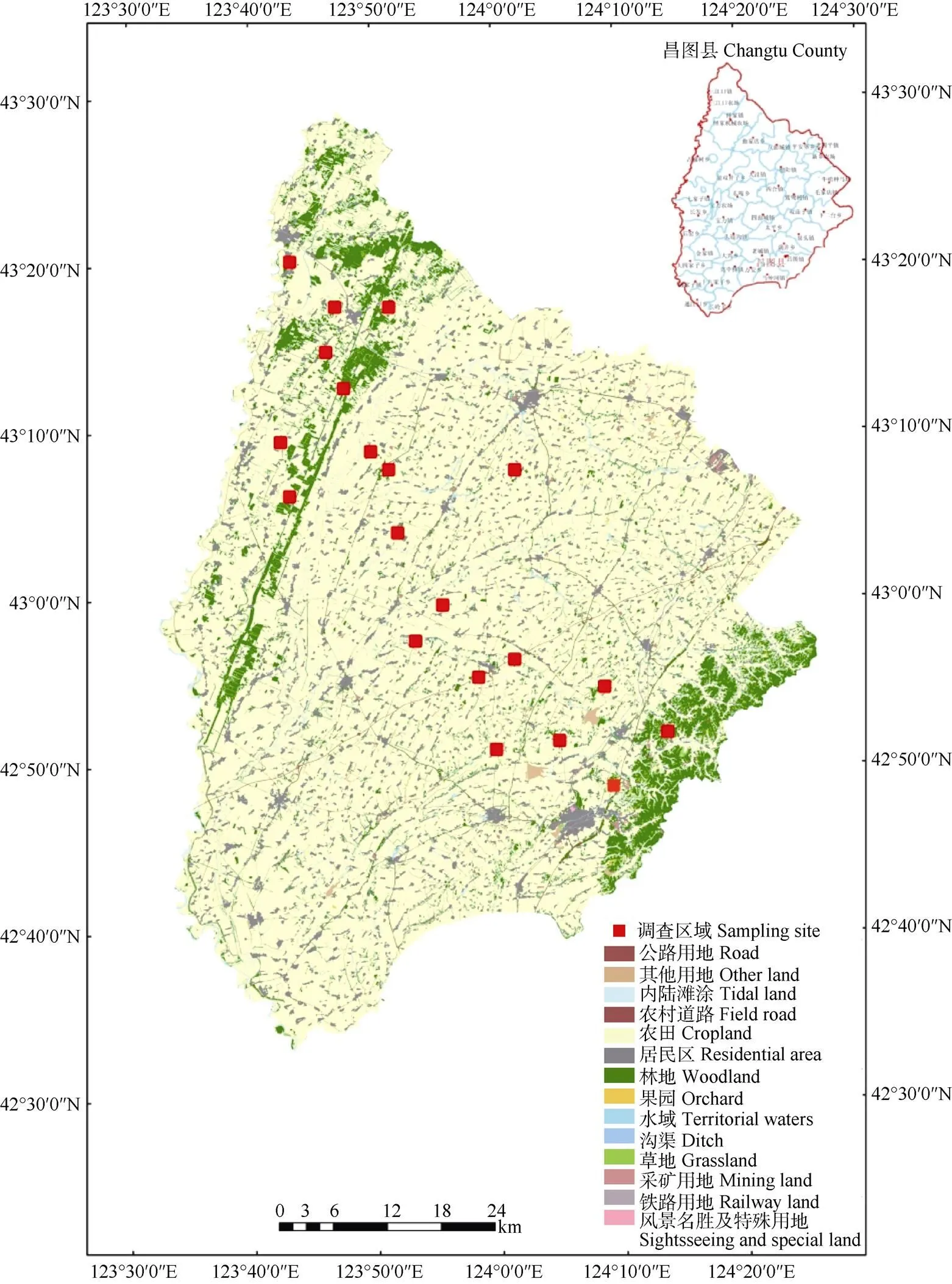
图1 研究区样点分布图
AIC=-2ln(模型的极大似然函数)+2(模型的独立参数个数) (1)
对于线性回归模型, AIC公式为:

2 结果与分析
2.1 6大类景观类型的主成分分析
通过对6大类16小类景观类型进行主成分分析, 选取累积贡献率大于80%的为主成分。在不同非耕作生境比例区间, 前2个或3个主成分的累积贡献率分别为82.5%、80.8%、81.0%、86.4%、87.6%。主成分载荷图如图3所示。不同非耕作生境比例下景观多样性指数(Simpson)值为0.67~3.65。
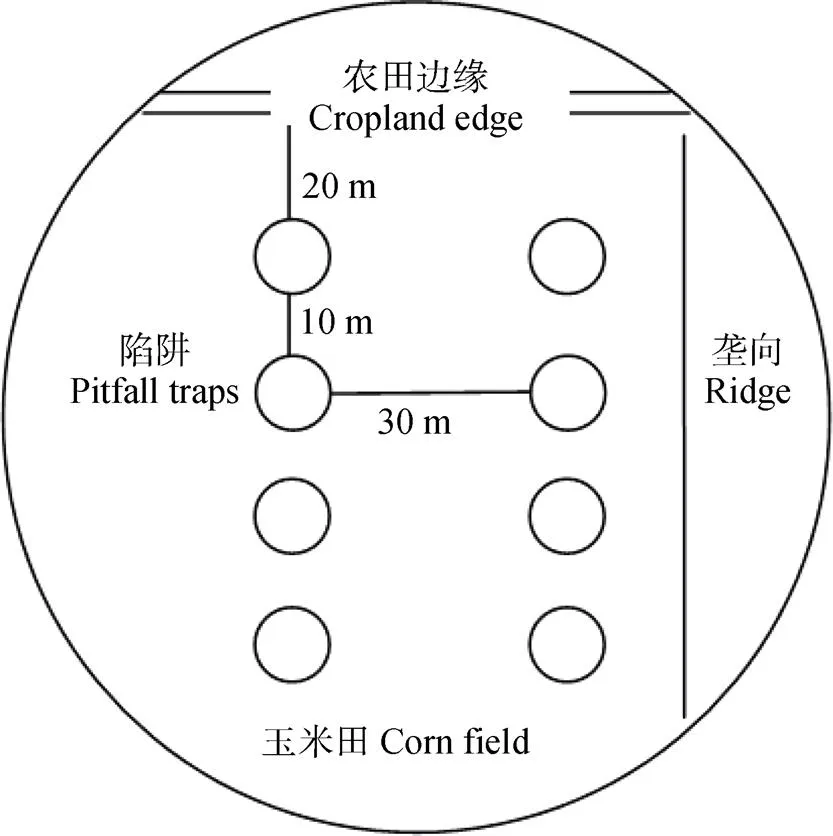
图2 玉米田中300 m半径的样方陷阱布设示意图
在非耕作生境比例0~10%时, 主成分1(PC1)代表以水域和草地为主的景观, 主成分2(PC2)代表以其他作物为主的景观, 主成分3(PC3)代表以林地为主的景观。在非耕作生境比例10%~20%时, 主成分1(PC1)代表以林地和村落为主的景观, 主成分2(PC2)代表以其他作物为主的景观。在非耕作生境比例20%~30%时, 主成分1(PC1)代表以草地和村落为主的景观, 主成分2(PC2)代表以林地为主的景观。在非耕作生境比例30%~40%、40%~50%时, 主成分1(PC1)均代表以林地为主的景观, 主成分2(PC2)均代表以其他作物为主的景观, 40%~50%时主成分3(PC3)代表以玉米为主的景观。
2.2 40个采样区中不同科的天敌种类及组成
采样收集天敌数量共7 455只, 均隶属于节肢动物门, 分为2纲6目10科。2纲分别为昆虫纲(Insecta)和蛛形纲(Arachnida), 6目为鞘翅目(Coleoptera)、膜翅目(Hymenoptera)、革翅目(Dermaptera)、半翅目(Hemiptera)、脉翅目(Neuroptera)和蛛形目(Aranelida), 10科为步甲科(Carabidae)、瓢虫科(Coccinellidae)、隐翅甲科(Staphylinidae)、蚁科(Formicinae)、姬蜂科(Lchneumqnidae)、蠼螋科(Labiduridae)、花蝽科(Anthocoridae)、草蛉科(Chrysopidae)、蟹蛛科(Thomisidae)和球蛛科(Theridiidae)。昆虫纲中尤以鞘翅目和膜翅目数量突出: 鞘翅目包括步甲科、瓢虫科、隐翅甲科3科, 分别占捕获量的36.00%、2.32%和2.10%; 膜翅目包括蚁科和姬蜂科, 分别占捕获量的41.76%和3.11%; 革翅目为蠼螋科, 占捕获量的2.35%; 半翅目为花蝽科, 占捕获量的2.64%; 脉翅目为草蛉科, 占捕获量的2.31%。蛛形目分为蟹蛛科和球蛛科, 分别占捕获量的3.74%和2.37%(表1)。
图4为玉米螟天敌在不同非耕作生境比例下的分布情况。由图显示, 在6月22—30日夏季采集样本中, 蚁科和步甲科数量最高。不同非耕作生境比例中蚁科数量分布不同, 在非耕作生境比例20%~30%时数量最高, 且在不同非耕作生境比例下差异性较显著; 步甲科在非耕作生境比例20%~30%时蚁科数量最高, 该比例下的标准差最小, 其次为10%~20%。9月22日—10月1日秋季采集样本中, 步甲科和蚁科数量最高, 非耕作比例为20%~30%时数量最高, 标准差最小, 其次为10%~20%。
研究区内在夏季和秋季不同非耕作生境比例下天敌数量研究结果表明(图5): 在夏季, 非耕作生境比例20%~30%时天敌数量最高, 0~10%时次之, 40%~50%时最低, 且0~10%、10%~20%和20%~30%比例之间差异性不显著, 30%~40%和40%~50%比例之间差异性不显著; 在秋季, 非耕作生境比例20%~30%时天敌数量显著高于其他4个比例, 10%~20%时次之, 40%~50%时最低。研究表明, 随着非耕作生境比例的增加, 天敌数量总体呈先上升后下降趋势, 在非耕作生境比例为20%~30%时达到峰值。其中, 夏季天敌数量在非耕作生境比例为0~10%时天敌数量较高, 推测原因可能是由于采样区紧挨着农田边缘的河流, 玉米螟天敌喜湿性环境, 形成大量聚集, 使得该比例区间内天敌数量较高。综合分析得出, 在非耕作生境比例20%~30%时天敌数量相对较高。
2.3 不同景观组成对天敌数量的影响
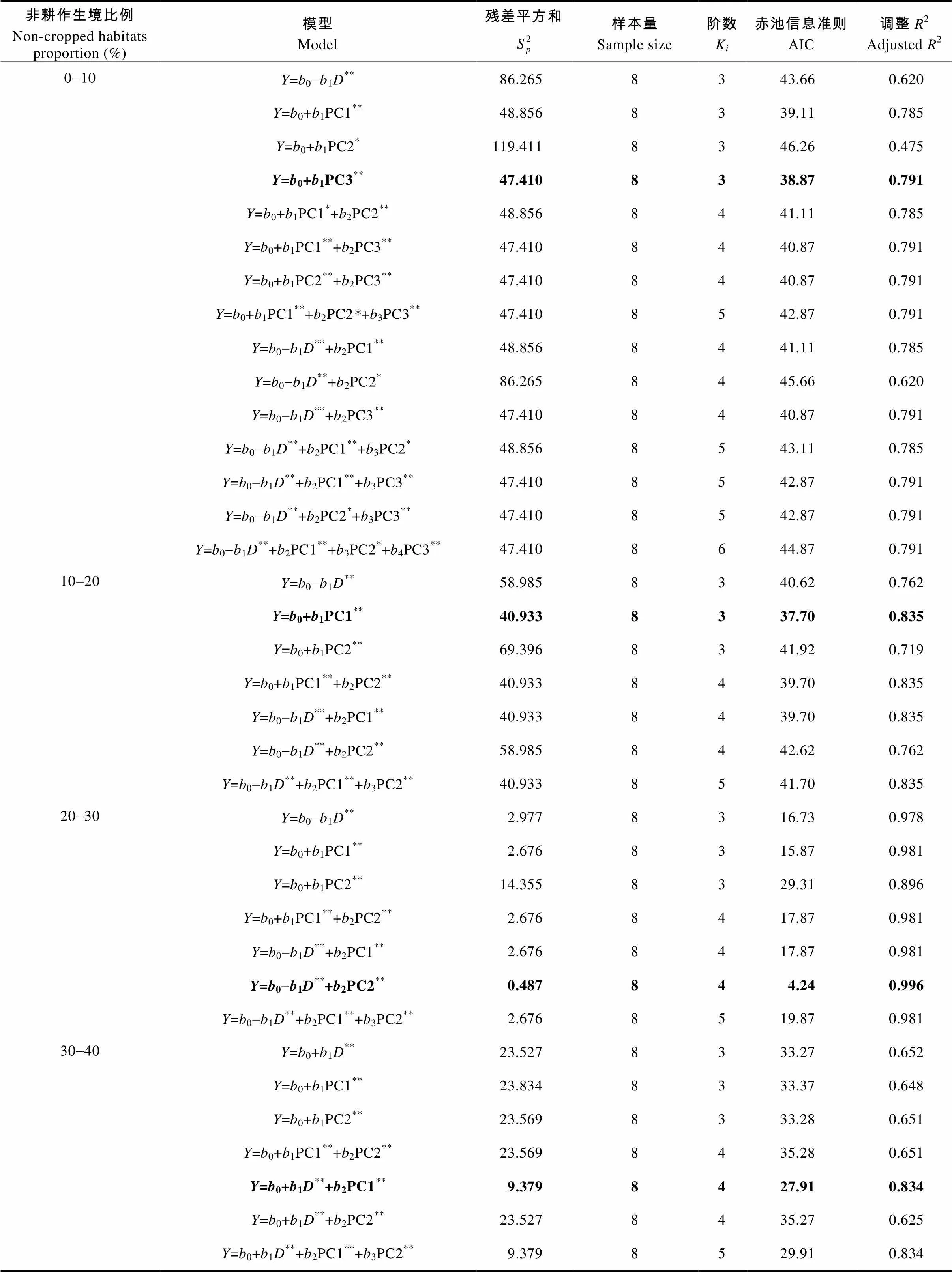
天敌数量与不同非耕作生境比例相关性分析表明(表2), 非耕作生境比例20%~30%时模型的AIC值最小(AIC=4.24), 为所有比例下的最优模型(=12.615-3.1731+3.7872,为玉米螟天敌数量,1为景观多样性指数,2为主成分2, 代表以林地为主的景观)。最优模型分析显示, 玉米螟天敌数量和主成分2(=3.787,=0.005)呈显著正相关, 与景观多样性指数(=-3.173,=0.005)呈显著负相关, 说明PC2代表的以林地生境为主的景观有利于提高农田中天敌数量, 但较高多样性的景观背景不利于天敌在农田的聚集。该比例下其他模型均显示玉米螟天敌数量与PC1(=1.957,=0.000)呈显著正相关, 说明PC1代表的以草地和村落为主的景观显著提高了农田中天敌数量。

图3 不同非耕地生境比例下农田景观类型的主成分分析

表1 20个调查区域采集的玉米螟天敌种类及数量

图4 夏季和秋季不同非耕作生境比例的玉米螟天敌数量分析特征

图5 夏季和秋季不同非耕作生境比例的玉米螟天敌数量分析特征
同一季节不同字母表示处理间差异显著。Different letters in the same season show significant differences at 0.05 level.
在另外4个非耕作生境比例中模型显示: PC3为非耕作生境比例0~10%的最优模型, PC1为10%~20%的最优模型,+PC1为30%~40%、40%~50%的最优模型, 模型中PC3(=6.456,=0.001)和PC1(=2.061,=0.000)均代表以林地生境为主的景观, 所有最优模型说明农田周边林地生境面积有利于提高农田中天敌数量。在非耕作生境10%~20%时, PC1(=2.061,=0.000)也代表以村落为主的景观, 说明村落面积增加也增加了农田内天敌数量, 在非耕作生境比例30%~40%和40%~50%时, 景观多样性指数(=2.939,=0.005;=2.291,=0.020)说明多样性的景观背景不利于农田内天敌的聚集。另外, PC2(<0.05)代表以其他作物为主的耕作生境与天敌数量呈显著正相关, 说明种植其他作物提高了农田内天敌的数量。
表2 不同非耕作生境比例下不同景观变量与玉米螟天敌数量分布的模型分析
Table 2 Models for quantities of corn borer’ natural enemies in corn field and different landscape variables under different proportions of non-cropped habitats
非耕作生境比例Non-cropped habitats proportion (%)模型Model残差平方和样本量Sample size 阶数Ki赤池信息准则AIC 调整R2Adjusted R20-10Y=b0-b1D**86.2658343.660.620Y=b0+b1PC1**48.8568339.110.785Y=b0+b1PC2*119.4118346.260.475Y=b0+b1PC3**47.4108338.870.791Y=b0+b1PC1*+b2PC2**48.8568441.110.785Y=b0+b1PC1**+b2PC3**47.4108440.870.791Y=b0+b1PC2**+b2PC3**47.4108440.870.791Y=b0+b1PC1**+b2PC2*+b3PC3**47.4108542.870.791Y=b0-b1D**+b2PC1**48.8568441.110.785Y=b0-b1D**+b2PC2*86.2658445.660.620Y=b0-b1D**+b2PC3**47.4108440.870.791Y=b0-b1D**+b2PC1**+b3PC2*48.8568543.110.785Y=b0-b1D**+b2PC1**+b3PC3**47.4108542.870.791Y=b0-b1D**+b2PC2*+b3PC3**47.4108542.870.791Y=b0-b1D**+b2PC1**+b3PC2*+b4PC3**47.4108644.870.79110-20Y=b0-b1D**58.9858340.620.762Y=b0+b1PC1**40.9338337.700.835Y=b0+b1PC2**69.3968341.920.719Y=b0+b1PC1**+b2PC2**40.9338439.700.835Y=b0-b1D**+b2PC1**40.9338439.700.835Y=b0-b1D**+b2PC2**58.9858442.620.762Y=b0-b1D**+b2PC1**+b3PC2**40.9338541.700.83520-30Y=b0-b1D**2.9778316.730.978Y=b0+b1PC1**2.6768315.870.981Y=b0+b1PC2**14.3558329.310.896Y=b0+b1PC1**+b2PC2**2.6768417.870.981Y=b0-b1D**+b2PC1**2.6768417.870.981Y=b0-b1D**+b2PC2**0.487844.240.996Y=b0-b1D**+b2PC1**+b3PC2**2.6768519.870.98130-40Y=b0+b1D**23.5278333.270.652Y=b0+b1PC1**23.8348333.370.648Y=b0+b1PC2**23.5698333.280.651Y=b0+b1PC1**+b2PC2**23.5698435.280.651Y=b0+b1D**+b2PC1**9.3798427.910.834Y=b0+b1D**+b2PC2**23.5278435.270.625Y=b0+b1D**+b2PC1**+b3PC2**9.3798529.910.834
表2 (续)

非耕作生境比例Non-cropped habitats proportion (%)模型Model残差平方和样本量Sample size 阶数Ki赤池信息准则AIC 调整R2Adjusted R240-50Y=b0+b1D*33.0868335.990.451Y=b0+b1PC1*29.5238335.080.510Y=b0+b1PC2**19.8098331.890.671Y=b0+b1(PC3*)28.3918334.770.529Y=b0+b1PC1*+b2PC2**19.8098433.890.671Y=b0+b1PC1*+b2(PC3*)19.8098433.890.671Y=b0+b1PC2**+b2(PC3*)28.3918436.770.529Y=b0+b1PC1*+b2PC2**+b3(PC3*)19.8098535.890.671Y=b0+b1D*+b2PC1*8.9118427.500.822Y=b0+b1D*+b2PC2**19.8098433.890.671Y=b0+b1D*+b2(PC3*)28.3918436.770.529Y=b0+b1D*+b2PC1*+b3PC2**19.8098535.890.671Y=b0+b1D*+b2PC1*+b3(PC3*)28.3918538.770.529Y=b0+b1D*+b2PC2**+b3(PC3*)19.8098535.890.671Y=b0+b1D*+b2PC1*+b3PC2**+b4(PC3*)19.8098637.890.671
列出了所有模型。其中字体加粗表示为最优模型, AIC值最小; 括号表示自变量和天敌数量呈负相关关系。: 天敌数量; D: 景观多样性指数; PC1: 主成分1; PC2: 主成分2; PC3: 主成分3。**: 在0.01水平(单侧)上显著相关; *: 在0.05水平(单侧)上显著相关。All the models are shown. Bold characters and numbers indicate the best model with the minimum AIC. Independent variables in parentheses are negatively related with quantities of corn borer’ natural enemies.represents Simpson; PC1 represents principal component 1; PC2 represents principal component 2; PC3 represents principal component 3. ** represents significant correlation at 0.01 level (unilateral); * represents significant correlation at 0.05 level (unilateral).
图6显示最优模型中天敌数量与景观组成的相关性。结果表明: 天敌数量和PC2(=3.787,=0.005)呈显著正相关, 但与景观多样性指数(=-3.173,=0.005)呈显著负相关。由此说明, 在非耕作生境20%~30%时, 林地生境面积增加有利于提高农田中天敌数量, 但较高多样性的景观背景不利于玉米螟天敌在农田内的集聚。
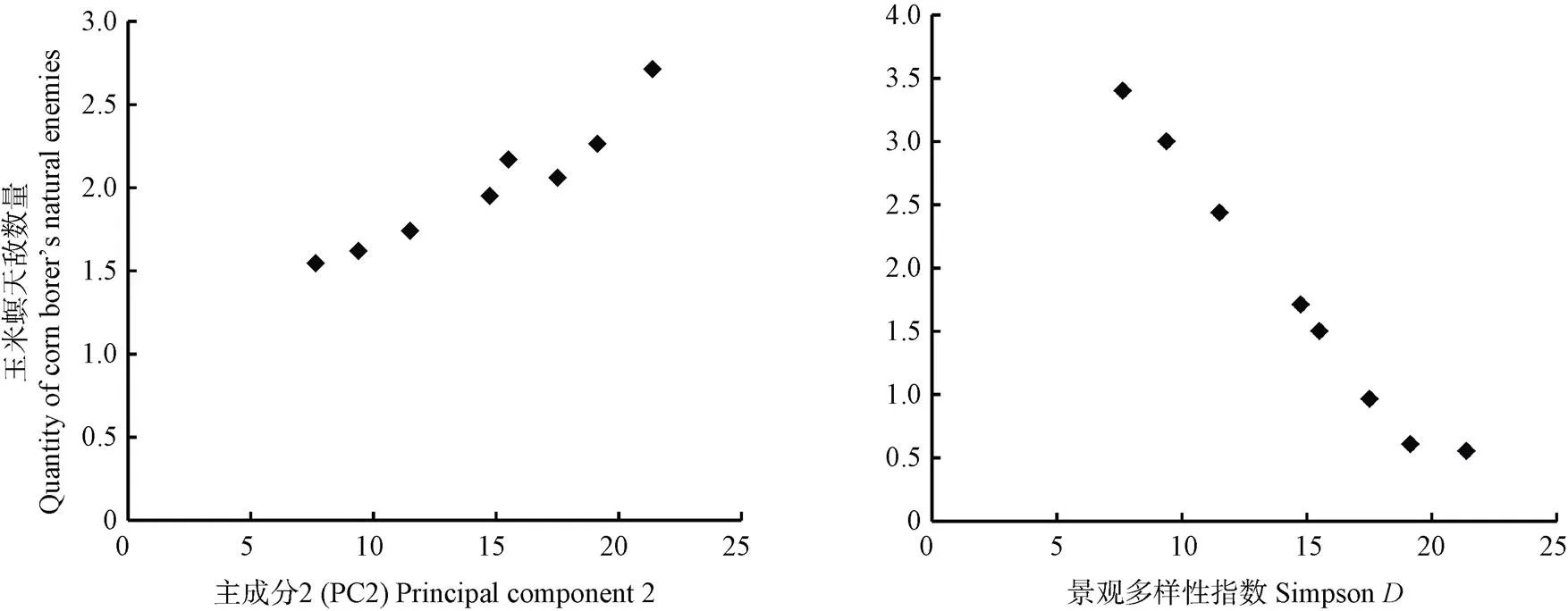
图6 非生境比例为20%~30%时最优模型中景观变量与玉米螟天敌数量的相关性分析
非耕作生境比例20%~30%上+PC2为所有区间最优模型。The best fit model is+PC2 under 20%-30% non-cropped habitats
2.4 农田景观中非耕作生境最优比例选取
不同非耕作生境比例对玉米螟天敌数量的影响具有一定的差异。在研究区内, 使用调整2平均值的大小判断不同非耕作生境比例对天敌数量的影响, 且调整2值越大, 相关性越大。从图7可以看出, 在非耕作生境比例为20%~30%时, 调整2值为0.97, 大于其他4个比例(0.69、0.80、0.70、0.64)的值。因此, 非耕作生境比例20%~30%是保持农田中玉米螟天敌最大数量的最佳比例。
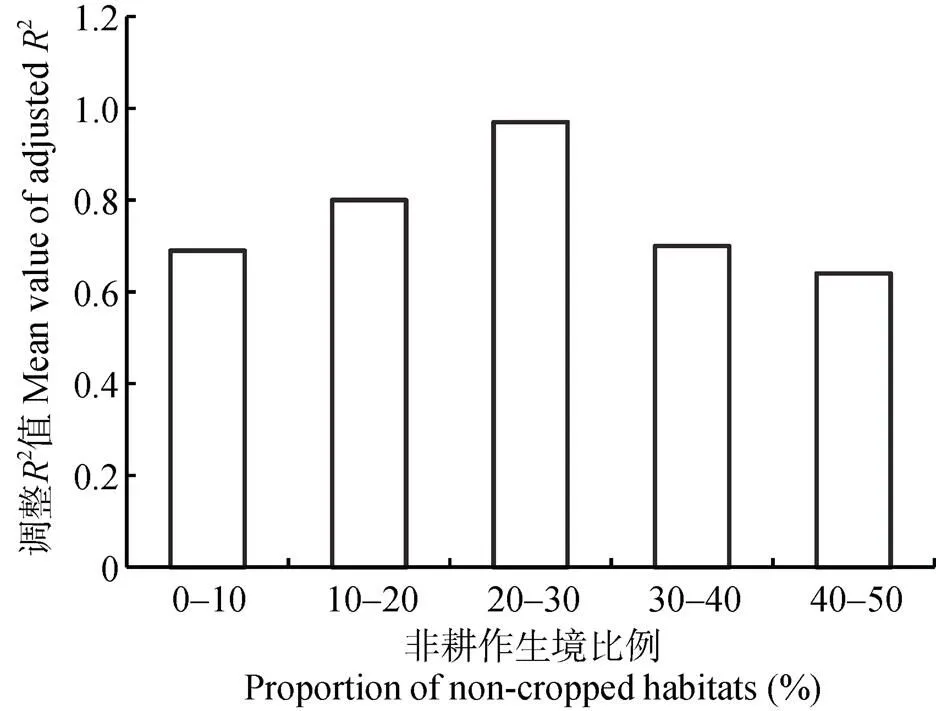
图7 非耕作生境比例与玉米螟天敌数量的关系
3 讨论与结论
本文研究了在0~50%不同非耕作生境比例区间内景观组成对玉米螟天敌数量的影响。研究表明随着非耕作生境比例的增加, 玉米螟天敌数量呈先上升后下降的趋势, 在非耕作生境比例为20%~30%时达最大值。由此证明非耕作生境比例为20%~30%时最有利于玉米螟天敌生存, 即对玉米螟防治最为有利。国外研究表明, 农田景观中非作物生境达20%时最能保护生物多样性和相联系的生态系统, 有利于生物害虫防治[35]。本研究结果与边振兴等[36]通过划分不同非耕作生境比例研究农业景观植物多样性得出的非耕作生境最佳比例为20%~30%的结果相同。在非耕作生境比例区间20%~50%时, 最优模型说明具有较高多样性的景观背景不利于天敌在农田的聚集。可能由于农田周边多样性的景观类型为天敌提供了更丰富的物质来源、避难所以及繁育场所等, 天敌更倾向于选择多样性的景观背景作为栖息环境, 从而减少了农田景观中天敌数量。不同季节对天敌数量表现出不同的影响, 其中夏季耕作生境为天敌提供了更多的食物来源和生存场所[16], 作物正处于生长季节, 害虫数量多, 人为干扰较少, 天敌更倾向于食物更丰富的农田生境。
研究结果同样表明以林地生境为主的景观组成有利于提高农田玉米螟天敌的数量。林地对农田中玉米螟天敌数量影响较为显著, 林地生境面积比例增加能够明显提高农田中玉米螟天敌数量。其原因在于农田景观中林地为永久型生境, 受人为干扰较少, 比耕地更具稳定性; 林地的保护与营建, 为农田景观中的天敌提供食物来源、避难场所和迁移廊道等。林地内有较多动植物残骸, 为天敌提供充足的食物源[37-39], 同时大部分天敌更倾向于温暖、潮湿及隐蔽性较好的生活环境, 例如蜘蛛、步甲、蠼螋等, 林地生境地表覆盖了枯枝落叶及草本植物, 形成了林地地表的湿润环境, 为天敌提供了更合适的生存场所。当进行除草、施肥及收割等耕作行为时导致耕作生境受到强烈扰动, 玉米螟天敌则迁移至农田周边的林地中躲避来降低危害。当耕作生境趋于稳定时, 天敌从林地迁移到耕作地境, 维持了农田生态系统的玉米螟天敌数量和多样性。村落、草地以及其他作物种植也提高了农田中玉米螟天敌的数量, 村落周边可能是因为住宅周边的林带、菜地、草垛等生境能为天敌提供越冬的场所和物质来源[40-41], 通过种植其他作物可以增加农田种植结构多样性, 同样可以为部分玉米螟天敌提供一定生存空间, 为天敌在作物间相互迁移提供扩散廊道和更多的食物源, 维持了更高的天敌丰富度, 对害虫的控害作用增强[42]。
本研究从景观生态角度研究玉米螟的防治效果, 对减少农药使用量, 保护农业生态系统中生物多样性具有重要的意义。同时研究证明在非耕作生境比例为20%~30%时玉米螟天敌数量达到最大值, 但对玉米螟天敌种类只划分到了不同科, 未细分到所属种。建议在今后研究中, 可细化天敌所属种, 具体研究所属种的生活习性, 以及具体哪种景观组合能提高农田里天敌数量, 依据景观特征制定合理有效的农业管理措施, 形成有效的农田景观类型, 减少农药施用, 实现可持续发展。
[1] THIES C, ROSCHEWITZ I, TSCHARNTKE T. The landscape context of cereal aphid-parasitoid interactions[J]. Proceedings of the Royal Society B: Biological Sciences, 2005, 272(1559): 203–210
[2] TSCHARNTKE T, STEFFAN-DEWENTER I, KRUESS A, et al. Contribution of small habitat fragments to conservation of insect communities of grassland-cropland landscapes[J]. Ecological Application, 2002, 12(2): 354–363
[3] BIANCHI F J J A, BOOIJ C J H, TSCHARNTKE T. Sustainable pest regulation in agricultural landscapes: A review on landscape composition, biodiversity and natural pest control[J]. Proceedings of the Royal Society B: Biological Sciences, 2006, 273(1595): 1715–1727
[4] 赵紫华, 关晓庆, 贺达汉. 农业景观结构对麦蚜寄生蜂群落组成的影响[J]. 应用昆虫学报, 2012, 49(1): 220–228 ZHAO Z H, GUAN X Q, HE D H. Community composition of parasitoids and hyperparasitoids of wheat aphids in different agricultural landscapes[J]. Chinese Journal of Applied Entomology, 2012, 49(1): 220–228
[5] FAHRIG L, BAUDRY J, BROTONS L, et al. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes[J]. Ecology Letters, 2011, 14(2): 101–112
[6] LAZZERINI G, CAMERA A, BENEDETTELLI S, et al. The role of field margins in agro-biodiversity management at the farm level[J]. Italian Journal of Agronomy, 2007, 2(2): 127–134
[7] SCHNEIDER S, WIDMER F, JACOT K, et al. Spatial distribution ofclade 1 in agricultural landscapes with arable land and different semi-natural habitats[J]. Applied Soil Ecology, 2012, 52: 20–28
[8] LANDIS D A, WRATTEN S D, GURR G M. Habitat management to conserve natural enemies of arthropod pests in agriculture[J]. Annual Review of Entomology, 2000, 45: 175–201
[9] RUSCH A, VALANTIN-MORISON M, SARTHOU J P, et al. Multi-scale effects of landscape complexity and crop management on pollen beetle parasitism rate[J]. Landscape Ecology, 2011, 26(4): 473–486
[10] CARVALHEIRO L G, SEYMOUR C L, NICOLSON S W, et al. Creating patches of native flowers facilitates crop pollination in large agricultural fields: Mango as a case study[J]. Journal of Applied Ecology, 2012, 49(6): 1373–1383
[11] GEIGER F, WÄCKERS F L, BIANCHI F J J A. Hibernation of predatory arthropods in semi-natural habitats[J]. BioControl, 2009, 54(4): 529–535
[12] SCHMIDT M H, THIES C, NENTWIG W, et al. Contrasting responses of arable spiders to the landscape matrix at different spatial scales[J]. Journal of Biogeography, 2008, 35(1): 157–166
[13] LEE J C, MENALLED F B, LANDIS D A. Refuge habitats modify impact of insecticide disturbance on carabid beetle communities[J]. Journal of Applied Ecology, 2001, 38(2): 472–483
[14] 尤民生, 侯有明, 刘雨芳, 等. 农田非作物生境调控与害虫综合治理[J]. 昆虫学报, 2004, 47(2): 260–268 YOU M S, HOU Y M, LIU Y F, et al. Non-crop habitat manipulation and integrated pest management in agroecosystems[J]. Acta Entomologica Sinica, 2004, 47(2): 260–268
[15] CARVELL C, MEEK W R, PYWELL R F, et al. Comparing the efficacy of agri-environment schemes to enhance bumble bee abundance and diversity on arable field margins[J]. Journal of Applied Ecology, 2007, 44(1): 29–40
[16] 刘文惠, 洪波, 胡懿君, 等. 不同景观结构下麦田地面甲虫和蜘蛛物种多样性及优势种分布的时空动态[J]. 应用昆虫学报, 2014, 51(5): 1299–1309 LIU W H, HONG B, HU Y J, et al. Space-time dynamic distribution of species diversity and dominant species of ground-dwelling beetle and spider populations on wheat crops in different agricultural landscape structures[J]. Chinese Journal of Applied Entomology, 2014, 51(5): 1299–1309
[17] PURTAUF T, DAUBER J, WOLTERS V. The response of carabids to landscape simplification differs between trophic groups[J]. Oecologia, 2005, 142(3): 458–464
[18] PURTAUF T, ROSCHEWITZ I, DAUBER J, et al. Landscape context of organic and conventional farms: Influences on carabid beetle diversity[J]. Agriculture, Ecosystems & Environment, 2005, 108(2): 165–174
[19] CLOUGH Y, KRUESS A, KLEIJN D, et al. Spider diversity in cereal fields: Comparing factors at local, landscape and regional scales[J]. Journal of Biogeography, 2005, 32(11): 2007–2014
[20] SCHMIDT M H, TSCHARNTKE T. Landscape context of sheetweb spider (Araneae: Linyphiidae) abundance in cereal fields[J]. Journal of Biogeography, 2005, 32(3): 467–473
[21] GARDINER M M, LANDIS D A, GRATTON C, et al. Landscape composition influences the activity density of Carabidae and Arachnida in soybean fields[J]. Biological Control, 2010, 55(1): 11–19
[22] GARDINER M M, LANDIS D A, GRATTON C, et al. Landscape composition influences patterns of native and exotic lady beetle abundance[J]. Diversity and Distributions, 2009b, 15(4): 554–564
[23] FENG Y, KRAVCHUK O, SANDHU H, et al. The activities of generalist parasitoids can be segregated between crop and adjacent non-crop habitats[J]. Journal of Pest Science, 2017, 90(1): 275–286
[24] DUELLI P, OBRIST M K. Regional biodiversity in an agricultural landscape: The contribution of seminatural habitat islands[J]. Basic and Applied Ecology, 2003, 4(2): 129–138
[25] 刘雨芳, 张古忍, 古德祥. 农田生态系统中生境与植被多样性对节肢动物群落的影响及其作用机制探讨[J]. 湘潭师范学院学报, 2000, 21(6): 74–78 LIU Y F, ZHANG G R, GU D X. Effect and the acting mechanisms of the habitats and vegetational diversity on arthropod community in agroecosystem[J]. Journal of Xiangtan Normal University, 2000, 21(6): 74–78
[26] HAWRO V, CERYNGIER P, TSCHARNTKE T, et al. Landscape complexity is not a major trigger of species richness and food web structure of European cereal aphid parasitoids[J]. BioControl, 2015, 60(4): 451–461
[27] ÖSTMAN Ö, EKBOM B, BENGTSSON J. Landscape heterogeneity and farming practice influence biological control[J]. Basic and Applied Ecology, 2001, 2(4): 365–371
[28] 张旭珠, 韩印, 宇振荣, 等. 半自然农田边界与相邻农田步甲和蜘蛛的时空分布[J]. 应用生态学报, 2017, 28(6): 1879–1888 ZHANG X Z, HAN Y, YU Z R, et al. Spatio-temporal distribution of carabids and spiders between semi-natural field margin and the adjacent crop fields in agricultural landscape[J]. Chinese Journal of Applied Ecology, 2017, 28(6): 1879–1888
[29] 赵爽, 宋博, 丁圣彦, 等. 黄河下游农业景观中景观结构和生境特征对林表生蜘蛛多样性的影响[J]. 生态学报, 2017, 37(6): 1816–1825 ZHAO S, SONG B, DING S Y, et al. Effects of landscape structure and habitat characteristics on spider diversity in the agro-landscape along the lower reaches of the Yellow River[J]. Acta Ecologica Sinica, 2017, 37(6): 1816–1825
[30] ZHOU K, HUANG J K, DENG X Z, et al. Effects of land use and insecticides on natural enemies of aphids in cotton: First evidence from smallholder agriculture in the North China Plain[J]. Agriculture, Ecosystems & Environment, 2014, 183: 176–184
[31] DONG Z K, OUYANG F, LU F, et al. Shelterbelts in agricultural landscapes enhance ladybeetle abundance in spillover from cropland to adjacent habitats[J]. BioControl, 2015, 60(3): 351–361
[32] GARDINER M M, LANDIS D A, GRATTON C, et al. Landscape diversity enhances biological control of an introduced crop pest in the north-central USA[J]. Ecological Applications, 2009, 19(1): 143–154
[33] AKAIKE H. Information theory and an extension of the maximum likelihood principle[M]//PARZEN E, TANABE K, KITAGAWA G. Selected Papers of Hirotugu Akaike. New York: Springer, 1998: 199–213
[34] 杨福芹, 戴华阳, 冯海宽, 等. 基于赤池信息准则的冬小麦植株氮含量高光谱估算[J]. 农业工程学报, 2016, 32(23): 161–167 YANG F Q, DAI H Y, FENG H K, et al. Hyperspectral estimation of plant nitrogen content based on Akaike’s information criterion[J]. Transactions of the Chinese Society of Agricultural Engineering, 2016, 32(23): 161–167
[35] TSCHARNTKE T, TYLIANAKIS J M, RAND T A, et al. Landscape moderation of biodiversity patterns and processes-eight hypotheses[J]. Biological Reviews, 2012, 87(3): 661–685
[36] 边振兴, 李晓璐, 于淼. 东北平原典型玉米种植区农业景观植物多样性研究——以昌图县为例[J]. 中国生态农业学报, 2018, 26(4): 480–492 BIAN Z X, LI X L, YU M. The plant diversity of agro-landscapes in typical maize planting areas in the Northeast Plain, China —A case study of Changtu County[J]. Chinese Journal of Eco-Agriculture, 2018, 26(4): 480–492
[37] TURNOCK W J, WISE I L. Density and survival of lady beetles (Coccinellidae) in overwintering sites in Manitoba[J]. The Canadian Field-Naturalist, 2004, 118(3): 309–317
[38] 杨龙, 徐磊, 刘冰, 等. 农田景观格局对华北地区麦田早期瓢虫种群发生的影响[J]. 应用昆虫学报, 2016, 53(3): 612–620 YANG L, XU L, LIU B, et al. Effects of landscape pattern on the occurrence of ladybeetles in wheat fields in northern China[J]. Chinese Journal of Applied Entomology, 2016, 53(3): 612–620
[39] LABRIE G, CODERRE D, LUCAS É. Overwintering strategy of multicolored Asian lady beetle (Coleoptera: Coccinellidae): Cold-free space as a factor of invasive success[J]. Annals of the Entomological Society of America, 2008, 101(5): 860–866
[40] Wang S, Michaud J P, Tan X L, et al. The aggregation behavior ofin its native range in Northeast China[J]. BioControl, 2011, 56(2): 193–206
[41] WERLING B P, GRATTON C. Local and broad scale landscape structure differentially impact predation of two potato pests[J]. Ecological Applications, 2010, 20(4): 1114–1125
[42] 邬建国. 景观生态学——格局、过程、尺度与等级[M]. 第2版. 北京: 高等教育出版社, 2007 WU J G. Landscape Ecology: Pattern, Process, Scale and Hierarchy[M]. 2nd ed. Beijing: Higher Education Press, 2007
Effect of agricultural landscape composition on natural enemy population of corn borer*
BIAN Zhenxing1, GONG Lingchun1, GUO Xiaoyu1, YU Miao2**
(1. College of Land and Environment, Shenyang Agricultural University, Shenyang 110161, China; 2. College of Science, Shenyang Agricultural University, Shenyang 110161, China)
The agricultural landscape with high proportion and diversity of non-cropped habitats is the key to maintaining agricultural biodiversity. It is a hypothesis that the natural enemy population and types of corn borer could increase by complex and high composition of non-cropped habitats in cornfields. To clarify the relationship between non-cropped habitats proportion and natural enemy, this study investigated the effects of composition of non-cropped habitats on natural enemy population of corn borer in Changtu County in the Northeast China Plain. Twenty plots (= 600 m) were selected and classed into five groups based on the proportions of non-cropped habitats (0-10%, 10%-20%, 20%-30%, 30%-40% and 40%-50%). The samples of natural enemies of corn borer were collected by the pitfall trap method. The families of enemies were identified using stereomicroscope. Then principal component analysis was used to classify the main types of non-cropped habitats in the region. Stepwise regression model was used to fit relation curve of non-cropped habitat composition (ratio and type) and population of natural enemies of corn borer. Then the optimal model was selected based on the Akaike’s Information Criterion (AIC) for effects of different proportions of non-cropped habitats on natural enemy population of corn borer. Results showed that natural enemy population of corn borer increased at first and then decreased with increasing proportion of non-cropped habitats. Natural enemy population of corn borer was highest [with the optimal model of AICminimum value= 4.24,=0-1+2PC2, PC2 (= 3.787,= 0.005)] when the proportion of non-cropped habitats was 20%-30%. This optimal model implied that there was significant positive correlation between natural enemy population of corn borer and the composition of non-cropped habitats dominated by woodland. There was also significantly negative correlation between natural enemy population of corn borer and the landscape diversity index. Other models [PC1 (= 1.957,= 0.000)] for 20%-30% proportion of non-cropped habitats implied that significant positive correlation existed between natural enemy population of corn borer and the composition of non-cropped habitats dominated by grassland and village. Under other proportions of non-cropped habitats, the optimal models suggested that corn filed neighboring woodland or other crops fields was beneficial for increase of natural enemy population of corn borer. In conclusion, the study showed that natural enemy population of corn borer was highest when the proportion of non-cropped habitats was 20%-30%. Non-cropped habitats dominated by woodland, grassland, village or in intercropped patterns attracted natural enemies of corn borer in farmlands with different proportions of non-cropped habitats. The study further provided theoretical basis for the prevention and control of corn borer by ways of landscape ecology.
Agricultural landscape; Non-cropped habitat; Corn field; Corn borer; Natural enemy
, E-mail: yumiao77@163.com
Mar. 7, 2018;
Jun. 20, 2018
P901
A
2096-6237(2019)01-0030-12
10.13930/j.cnki.cjea.180221
边振兴, 龚玲春, 果晓玉, 于淼. 农业景观组成对玉米螟天敌数量的影响[J]. 中国生态农业学报(中英文), 2019, 27(1): 30-41
BIAN Z X, GONG L C, GUO X Y, YU M. Effect of agricultural landscape composition on natural enemy population of corn borer[J]. Chinese Journal of Eco-Agriculture, 2019, 27(1): 30-41
* 沈阳农业大学土地与环境学院青年创新项目(20140102)资助
于淼, 研究方向为数理统计与景观生态学。E-mail: yumiao77@163.com
边振兴, 研究方向为农地利用与保护、农业景观生态学。E-mail: zhx-bian@263.net
2018-03-07
2018-06-20
* This study was supported by the Youth Innovation Project of Land and Environment College of Shenyang Agricultural University (20140102).
猜你喜欢
今日农业(2021年14期)2021-11-25 23:57:29
落叶果树(2021年6期)2021-02-12 01:29:10
今日农业(2020年14期)2020-08-14 01:10:20
水土保持研究(2016年4期)2016-10-21 02:28:59
新农业(2016年15期)2016-08-16 03:40:48
现代农业(2016年5期)2016-02-28 18:42:38
体育科技(2016年2期)2016-02-28 17:06:07
土壤与作物(2015年3期)2015-12-08 00:46:58
现代农业(2015年5期)2015-02-28 18:40:49
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:17
