
氧化应激对反刍动物胃肠道的影响及其作用机制
2018-12-29 03:13:32孔凡林刁其玉
饲料工业 2018年23期
■孔凡林 刁其玉 屠 焰
(中国农业科学院饲料研究所奶牛营养学北京市重点实验室,北京100081)
目前,随着幼畜早期断奶技术应用的日益广泛和全球变暖带来的不利影响日益严重,由断奶应激引起的幼畜抗氧化功能失调以及由热应激引起的成年反刍动物抗氧化功能失调已成为制约反刍动物正常生长发育的重要因素之一,近年来关于氧化应激发生机制及其对反刍动物胃肠道消化吸收的影响机制的相关研究也相对增多。
现已经证明,在我国现代畜牧业的养殖过程中,氧化应激的发生使畜禽的生长性能下降、免疫性能下降、血清生化指标异常[1],最终导致多种疾病的发生,如乳房炎[2],导致疾病发生的原因在于动物体内氧化系统和抗氧化系统的失衡,这种失衡使得过多的高活性氧自由基、氮自由基在体内堆积,不能被及时的清除,自由基的堆积会通过引起细胞内脂质的过氧化,损伤DNA分子或调节相关基因表达来影响机体的生长发育[3]。动物的消化器官不仅在营养物质的消化吸收过程中担任重要角色,同样也参与机体的免疫和机械防御过程[4]。所以,研究氧化应激的机制及其对胃肠道结构和功能的影响不仅可以为反刍动物的生长调控提供依据,同时也为如何缓解氧化应激提供指导。
1 氧化应激的发生机制
正常动物体内自由基的产生和消耗一直维持着动态的平衡,这种平衡一旦被体内或体外的刺激打破,自由基的数量便会超出机体抗氧化系统的忍耐范围,造成氧化应激进而产生代谢疾病,所以患者机体氧化应激状态对病症的判定具有十分重要的临床指导意义,通过检测血清中的氧化损伤产物、抗氧化剂及抗氧化酶类水平可以对机体氧化应激状态进行判定,相关研究报道蛋白羰基、晚期氧化蛋白产物、丙二醛和8-羟基脱氧尿苷是有效的过氧化指标[5-7]。
1.1 自由基
自由基,在化学上也被称为“游离基”,是指含有不成对电子的原子团,这一性质使得其化学性质极不稳定,容易与其他物质争夺电子。在动物体内含量较多的自由基为氧自由基,主要包括超氧阴离子自由基和羟自由基等,再加上过氧化氢、单线态氧和臭氧等统称为活性氧(ROS)。研究表明低浓度活性氧的存在是正常的生理现象,体内纳摩尔水平的活性氧对激活海脑回发育信号通路至关重要,在其中扮演着信号分子的作用[8],但当自由基的数量超出机体抗氧化系统的承受能力后便会对细胞造成损伤。
在机体中90%的氧在线粒体中被消耗,所以ETC是产生ROS数量最多的途径,ROS产生的机制在于ETC过程不具有绝缘性,会在某些特定地点有漏电现象[9]。线粒体中至少有十个以上已经确定的部位会产生超氧阴离子自由基(superoxide anion free radical,·),如复合体Ⅰ、复合体Ⅲ等,Quinlan等[10]已经证明在使用鱼藤酮高度抑制ETC过程中复合体Ⅰ的活性后,电子无法从还原型辅酶1传递给辅酶Q,但线粒体上的其他酶类却开始产生线粒体氧自由基,如α-酮戊二酸脱氢酶、丙酮酸脱氢酶,且合成效率是复合体Ⅰ的6~8倍。另外,人为使用抗霉素A抑制复合体Ⅲ的酶活性后也会产生ROS。ROS在ETC中的主要产生部位如图1所示。
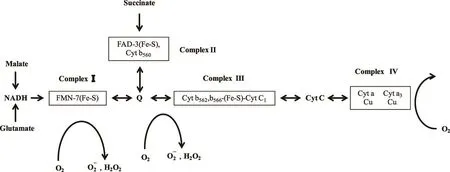
图1 活性氧在ETC中的主要产生部位
除活性氧以外,动物体内还存在着另一类具有高度氧化活性的自由基,该类自由基为体内一氧化氮(NO)代谢途径产生的一系列硝基化合物,总称为活性氮(RNS)。与ROS相比,RNS在体内的含量较少,由细胞内的一氧化氮合酶(nitric oxide synthase,NOS)催化生成,根据NOS在生物体内所在位置的不同可分为三种,分别是神经型、内皮型和诱导型。其中,神经型和内皮型NOS的表达受Ca2+浓度的调控,而诱导型NOS则不受Ca2+浓度的调控[13]。NOS催化L-精氨酸与氧原子结合生成NO和L-瓜氨酸,过多的NO又可继续与·结合形成过氧化亚硝基(ONOO-)[14],ONOO-是RNS功能的实现者,可使蛋白质组成中的酪氨酸硝基化,从而改变蛋白质正常的结构和功能。
1.2 抗氧化系统
生物体内存在着的抗氧化系统十分复杂,由多种成分组成,单一的抗氧化物所具有的抗氧化能力十分有限,往往需要通过联级反应和逐级放大反应来提高机体的抗氧化能力,这就需要从总体的水平上来研究抗氧化系统。
抗氧化系统具体可分为酶类抗氧化系统和非酶类抗氧化系统。
1.2.1 酶类抗氧化系统
酶类抗氧化系统是指一类在氧化还原反应中具有催化作用,能够清除或转化自由基和ROS的酶类,如超氧化物歧化酶(SOD)、过氧化氢酶(CAT)及谷胱甘肽过氧化物酶(GSH-Px)等。SOD广泛的存在于各种生物体内并专一的用于清除·,SOD催化·发生歧化反应并通过转移一个电子生成毒性更低的过氧化氢和氧气(2·+2H+→H2O2+O2)。研究发现Cu/Zn-SOD和Mn-SOD的mRNA表达量在病原体注射后显著升高,且该过程主要的转录调控因子对Mn-SOD启动子的激活情况相较于Cu/Zn-SOD更为有效[15],所以Mn-SOD在抗氧化过程中占主导地位。此外,Khanna-chopra等[16]在藜叶子、花序的叶绿体中成功提取出SOD的同工酶,当温度升高到100℃时,该同工酶的活性仍能保持原有活性的80%,具有良好的热稳定性和应用前景。
GSH-Px为水溶性四聚体蛋白,4个亚基结构相似,每个亚基上有1个硒原子,研究发现在羔羊饲料中添加NanoSe或NaSe可提高血液中GSH-Px的活性(P<0.01)[17],这可能与GSH-Px的结构组成有很大关系。
CAT的主要作用是催化过氧化氢生成氧和水,当机体CAT的含量很低时,非酶类抗氧化系统可代替CAT来清除过氧化氢。
1.2.2 非酶类抗氧化系统
常见的非酶类抗氧化剂包括人工合成的特丁基对苯二酚(tertiary butylhydroquinone,TBHQ)、丁基羟基茴香醚(butylated hydroxyanisole,BHA)等,有研究证明,TBHQ与牛血清蛋白的结合非常紧密,两者之间的相互作用导致了牛血清蛋白构象的改变[18-19],其构象的改变对功能的影响仍需要进一步的研究;维生素及微量元素类抗氧化剂包括维生素A、维生素C、维生素E、α-硫辛酸、β-胡萝卜、硒、锰、铜、锌等,这些物质已经作为饲料中常见的添加成分广泛应用于生产实践中。
酶类抗氧化系统的生成往往需要非酶类抗氧化系统作为前体物质,硒参与构成的硒代半胱氨酸作为合成GSH-Px的原料中之一,研究已经证明在日粮中添加硒可显著提高SOD和GSH-Px的活性[20]。此外,Elshenawy等[21]还发现,在硒的基础上添加维生素E可明显提高叠氮化钠(NaN3)处理小鼠的免疫性能并缓解NaN3造成的器官损伤,这说明二者之间有协同作用。但Liu等[22]的研究表明维生素E和硒虽然都可以减轻氧化应激对育成猪的影响,二者同时添加却不具有组合效应,所以二者在提高反刍动物的抗氧化性能方面是否存在协同效应仍需要进一步的研究。
非酶类抗氧化剂还包括从动植物中提取出来的天然抗氧化剂,如茶多酚、虾青素、花青素等,有研究表明茶多酚、虾青素、花青素能有效缓解氧化应激和炎症反应[23-25]。
2 氧化应激对反刍动物胃肠道的影响
氧化应激对动物机体的神经、体液、免疫系统都有着极其复杂的影响,且三者之间相互关联,根据氧化物对胃肠道器官损伤部位、途径和影响程度的不同,氧化应激对胃肠道的影响可通过三个主要方面进行解释。
2.1 诱导胃肠道上皮细胞凋亡
反刍动物胃肠道结构的完整性是营养物质在消化器官中消化吸收的物质基础,氧化应激可通过诱导胃肠道上皮细胞凋亡来破坏这种完整性,从而降低对营养物质的吸收效率,马燕芬等[26]的研究证明,奶山羊在热应激状态下瘤胃乳头坏死、乳头脱落情况较为严重,瘤胃背囊、腹囊的绒毛长度和宽度均小于正常情况,并且随着作用时间的延长,效果越严重。同时,干物质、粗蛋白、有机物消化率和产奶量均显著降低。朱旺[27]的研究表明肉牛的空肠上皮细胞在应激状态下活性下降,抗氧化能力下降,韩奇鹏等[28]的研究也证明山羊瘤胃上皮细胞在氧化应激状态下时凋亡率显著增加,建议使用H2O2诱导稳定的山羊瘤胃上皮损伤模型。
氧化应激造成细胞凋亡的起始原因是反刍动物在应激状态下会进行血液的重新分配,使消化系统经历“缺血再灌注”过程。缺血再灌注过程是指机体在应激状态下会激活肾素-血管紧张素-醛固酮系统,该系统是调节心血管活动的重要体液因素,这些激素的分泌保证了应激状态下心、脑、骨骼肌的血液供应以应对环境的变化,但消化系统的血液供应则相应减少[29],一方面,对消化系统的血液供给减少会导致胃肠道上皮细胞获得的用于发育、分裂的营养物质减少,这就不可避免的造成消化系统发育受阻;另一方面,“缺血再灌注”过程已经被证明是诱导细胞凋亡的诱因之一,细胞凋亡的主要作用通路是caspase依赖性途径,根据激活caspase家族的途径不同又可分为死亡受体途径和线粒体途径,线粒体作为细胞的“动力工厂”,已经被证明参与细胞凋亡[30],近年来发现内质网也存在有细胞凋亡的通路[31],但相关研究较少。
2.1.1 线粒体凋亡途径
线粒体凋亡途径是caspase依赖性途径的主要分支,他的关键之处在于线粒体外膜通透性(major outer membrane protein,MOMP)的转变,MOMP的升高可释放储存在线粒体中的凋亡因子并启动细胞凋亡程序。而MOMP改变的原因之一在于氧化应激状态下腺嘌呤核糖核苷酸(adenosine monophosphate,AMP)的不断堆积,王兰芳等[32]的体外细胞培养试验证明,外源添加1 000 μmol/l的AMP可显著增加细胞凋亡率,但相对低浓度的AMP对其没有影响,这是因为缺血初期导致的胃肠道上皮细胞缺氧使得线粒体无法有效进行氧化磷酸化产生ATP,机体对ATP的需求导致糖酵解作用增加,进而乳酸堆积,线粒体膜电位下降,为了维持膜电位的稳定,线粒体变成了ATP的主要消耗者,大量ATP转化成AMP导致线粒体中AMP/ATP比例增加,AMP活性蛋白激酶(adenosine monophosphate activated protein kinase,AMPK)活性增加,AMPK磷酸化激活结节性硬化复合物1/2(tuberous sclerosis complex,TSC1/2),TSC1具有激活GTP酶的功能,从而抑制雷帕霉素靶蛋白并促进自噬[33],随着缺血时间的延长,高浓度的AMP可直接激活磷脂酶A2,降解线粒体膜磷脂,破坏线粒体膜完整性,一方面使得调节渗透压的能力下降,另一方面导致细胞色素C逸出,最终诱导细胞凋亡[34]。
氧化应激也可以通过直接对线粒体膜进行氧化,进而降低线粒体膜的通透性。耿雅丽等[35]研究发现,长期饲喂高精料的泌乳奶山羊细胞周期相关基因表达水平显著升高,促细胞凋亡相关基因表达显著升高,胆固醇含量显著下降。胆固醇作为细胞膜和细胞器膜的重要组成成分,对维持生物膜的流动性和完整性有重要作用,研究证明ROS对胆固醇的双键有氧化作用,而细胞膜或线粒体膜上脂质的过氧化会导致自身的通透性增加[36]。
氧化应激还可以通过控制Bcl-2蛋白家族进而改变MOMP,Bcl-2蛋白家族主要定位在线粒体外膜上,按照对凋亡途径的促进或阻碍作用分成效应者、保护者与起始者。吴黎明[37]使用H2O2处理初生奶牛的小肠上皮细胞后发现,Bax和caspase-3基因表达量显著增加,Bax蛋白为效应者,是含有多个同源结构域的促凋亡蛋白,当氧化应激发生后,游离的Bax可直接在线粒体膜上形成Bax-Bax同二聚体,增加MOMP并释放细胞色素C到细胞质,细胞色素C在胞质中能够与凋亡酶激活因子(apoptotic enzyme activator,Apaf-1)的碳端结构域结合形成复合体并通过多聚化作用激活Apaf-1,Apaf-1随后募集caspase-9从而形成凋亡体,随后激活下游的caspase级联反应,最后的激活物caspase-3可诱导细胞的凋亡[38]。然而,除效应者以外还存在着与之有相反作用的保护者,徐霄龙等[39]使用高温诱导肠上皮细胞氧化应激,结果显示caspase-3、caspase-9和Bax表达量显著升高,Bcl-2显著下降。蔡亚非等[40]研究发现,奶牛在氧化应激状态下淋巴细胞凋亡率由2.7%提高到10.6%,同时Bcl-2丰度显著降低。其中的Bcl-2可通过与效应者的结合来降低MOMP,维持线粒体膜的稳定性和内外渗透压,也可通过与线粒体去极化产生通透性转换孔(mitochondrial permeability transition pore,mPTP)相互作用,抑制mPTP功能[41],主要成员除Bcl-2外还有Bcl-xl。
2.1.2 死亡受体途径
在死亡受体途径中,H2O2诱导死亡因子受体(Fas)与其配体(FasL)过渡表达,FasL可与Fas结合并活化Fas的“死亡区域”(dead domain,DD)从而形成三聚体,DD具有分解蛋白质的功能,DD利用该功能可募集Fas相关死亡蛋白(fas-associated protein with death domain,FADD)并形成死亡诱导复合物(deathinducing signaling complex,DISC),DISC在胞质内促使caspase-8前体通过自身的剪切作用断裂、聚集、活化,活化后的caspase-8进一步启动下游的caspase级联反应,最终导致细胞的凋亡[42]。该通路在单胃动物身上已经得到了验证,但在反刍动物上的研究还处于起步阶段,陶诗煜等[43]研究发现饲喂高精料日粮的波尔山羊结肠上皮组织caspase-8基因表达显著增加,证明了死亡通路在胃肠道上皮细胞中确实存在。总之,“缺血再灌注”过程首先通过减少胃肠道的血液供给来影响其发育,更深层次的影响是通过该过程引起的氧化应激及细胞凋亡来完成的。
细胞凋亡与细胞增殖在体内的平衡维持着机体内环境的稳态,位于身体不同部位和器官的细胞因为凋亡过度所引发的疾病也有所不同,研究证明阿兹海默症是由于神经细胞凋亡过多导致的[44],帕金森症是由于多巴胺神经细胞凋亡过多导致的[45],由于中枢神经细胞的不可再生性使得此类病症几乎不能被治愈,但对于可增殖的胃肠道上皮细胞或血管内皮细胞来说,是否致病取决于增殖和凋亡的平衡,如果细胞凋亡在细胞增殖的控制范围内,细胞凋亡便不足以成为致病原因,如果细胞凋亡超出细胞增殖的控制范围则会引发多种疾病。细胞凋亡途径在单胃动物上面已经研究的较为充分,但在反刍动物上还缺乏详尽的报道,所以还需要进一步的研究。
2.2 对胃肠道屏障功能的影响机制
胃肠屏障是指能够阻止外源有害有毒物质穿过肠胃黏膜进入其他器官、组织的结构和功能的总和。肠道的第一道防线为机械屏障,它由肠道黏膜上皮细胞、细胞间紧密连接(TJ)和黏液层三者构成,其功能在于控制水和溶质的跨膜运输,氧化应激则通过改变TJ的组成和结构来影响胃肠道上皮通透性,进而影响营养物质的消化吸收过程和对有害物质的过滤过程[46]。TJ由结构蛋白和功能蛋白两部分组成,结构蛋白包括闭合蛋白(Occludin)、密封蛋白(Claudin)等,其中的Occludin蛋白和Claudin蛋白不仅参与TJ的物理构成,研究证明Occludin蛋白还对细胞间通透性具有调节作用,Claudin蛋白还参与构成TJ中只允许水分子和小分子物质通过的“小孔”结构[47]。闭合小环蛋白(zo-na occludens,Zos)与前两种蛋白一样同样参与到细胞物质转运中,且还具有维持上皮极性的功能,马燕芬等[48]的研究发现应激组奶山羊的瘤胃上皮细胞中claudin、ZO-1基因的mRNA表达量显著低于对照组,并且随着应激时间的增加,紧密连接蛋白的损伤越明显。陈新瑶等[49]使用H2O2对小肠上皮细胞进行处理后得到了同样的结果。以上研究说明氧化应激对反刍动物胃肠道上皮TJ结构中的主要蛋白表达有消极影响,但TJ结构复杂,除Occludin蛋白和Claudin蛋白外还存在着连接粘附分子、闭锁小带蛋白等,他们对控制物质转运有着各自的功能,但目前却少有报道。
2.3 通过体液调节对胃肠道发育的影响机制
应激的发生会引起广泛的神经内分泌变化,为解释这一现象,在1951年,应激学说的奠基人Selye提出了“全身适应综合征”这一概念,全身适应综合征是指当机体内环境稳态受威胁、扰乱后出现的一系列生理和行为的适应性反应,其中最为经典的便是下丘脑-垂体-肾上腺皮质激素轴,通过下丘脑引起血中促肾上腺皮质激素浓度迅速升高,糖皮质激素大量分泌的过程,其主要成分为皮质醇,糖皮质激素受体广泛的存在于机体大部分组织中,胃肠道也不例外,余思佳等[50]研究证明随着妊娠的进行,繁殖母羊体内的自由基代谢逐渐增强,血清皮质醇含量逐渐增加,且在分娩时达到最大值,研究证明人毛发中皮质醇和可的松含量与代谢紊乱和心血管疾病的发生风险呈正相关关系,糖皮质激素的异常分泌容易引起代谢综合征,主要表现为机体蛋白质、脂肪、碳水化合物等物质的代谢紊乱[51],进而间接影响胃肠道发育。近期的研究表明毛发中皮质醇含量不能很好的代表机体皮质醇的分泌量,毛发取样部位不同导致的测定结果也不同[52-53],所以其具体机制还需要试验探究。此外,皮质醇分泌增多对反刍动物胃肠道的影响还需要进一步验证。
氧化应激引起的代谢综合征,另一重要特征为胰岛素抵抗[54],胰岛素抵抗如果进一步恶化可引发Ⅱ型糖尿病和心血管疾病,这两大病症之间相互影响、并发存在,之间有着极其复杂的关联性[55],还有研究表明胰岛素抵抗会降低血管内皮细胞上纤维蛋白溶解酶的活性[56],该酶活性的降低会严重影响瘤胃周围血管的血流畅通,进而降低瘤胃获取营养的效率。
有研究证明在长期应激状态下,血清中促甲状腺释放激素的含量与ROS含量呈负相关关系,即氧化应激导致甲状腺释放激素的释放量减少[57],其导致的结果是瘤胃的正常发育因为甲状腺激素分泌量的减少而受到影响。除糖皮质激素、胰岛素和甲状腺激素外,氧化应激对机体多种激素的合成和释放均造成影响,但对胃肠道的影响微乎其微,所以在此不一一列举。
3 结论
氧化应激的发生机制已经十分完善且后续对人等单胃动物的影响也都有较为系统的陈述,但在反刍动物身上,由于瘤胃的特殊性,氧化应激对于胃肠道的影响机制仍处于理论研究阶段。一方面,关于氧化应激引发的反刍动物胃肠道上皮细胞凋亡,尤其是瘤胃上皮细胞的凋亡机制仍有待研究,目前仅有少数试验证明了表观现象,研究还不够深入;另一方面,氧化应激对反刍动物胃肠道紧密连接的影响机制还研究的不够全面,仅证明了少数紧密连接蛋白的重要性,而构成紧密连接的蛋白种类众多。所以,今后的研究应集中在利用分子生物学技术对氧化应激造成的胃肠道变化进行跟踪,并在此基础上筛选出具有针对性抗氧化作用的饲料添加剂。
猜你喜欢
科学(2022年4期)2022-10-25 02:43:06
中国典型病例大全(2022年9期)2022-04-19 22:12:45
草业科学(2021年7期)2021-08-03 09:22:14
中华养生保健(2020年10期)2021-01-18 06:46:06
科学(2020年2期)2020-08-24 07:56:44
饮食保健(2019年21期)2019-01-12 13:28:58
人人健康(2017年5期)2017-03-20 22:30:54
湖南畜牧兽医(2016年3期)2016-06-05 08:37:56
动物营养学报(2015年10期)2015-12-01 02:26:20
中国合理用药探索(2014年1期)2014-03-11 20:30:17
