
荧光原位杂交技术在蔷薇属植物研究中的应用综述
2018-12-19 09:04唐开学邱显钦晏慧君王其刚蹇洪英
江苏农业科学 2018年22期
田 敏, 张 婷, 唐开学, 张 颢, 邱显钦, 晏慧君, 王其刚, 蹇洪英
(云南省农业科学院花卉研究所/云南省花卉育种重点实验室/国家观赏园艺工程技术研究中心,云南昆明 650200)
荧光原位杂交(fluorescent in situ hybridization,FISH)技术是一种非放射性原位杂交技术。它的基本原理是将待测的核酸(DNA或RNA)用连接有报告分子的核苷酸(如biotin-dUTP或digoxigenin-dUTP)标记成探针,然后将标记的探针按碱基互补配对原理杂交到染色体上;检测时用与荧光素耦连的抗报告分子的单克隆抗体(如anti-digoxigenin-Rhodamine)或特异结合蛋白(如avidin-FITC)与报告分子标记的核苷酸探针特异性结合,通过荧光素的显色来确定探针与探针标记的靶核酸的共有序列及这些序列在受体细胞核和染色体上的物理分布,对其进行定位、定性及定量研究。FISH技术具有试验周期短、灵敏度高、分辨率高、直观可见等优点[1]。随着荧光原位杂交技术的深入和多元发展,该方法已用于核酸研究的各个方面[2]。在植物中,荧光原位杂交技术主要应用于多倍体的起源和演化[3],植物种间杂交后代的鉴定和外源染色体检测[4],构建DNA物理图谱分析染色体的易位、倒位、互换及重组等结构变异等研究。此外,FISH在转基因植株的细胞学鉴定、胚挽救、体细胞融合植株的早期鉴定、遗传转化材料分析、基因组的结构及其在细胞中的空间排列与功能等研究中也发挥着重要作用,展示了广阔的应用前景。
现代月季为蔷薇属(RosaL.)植物,是世界第一大切花,是由蔷薇属内很多种或品种反复杂交而成的一大类栽培复合体[5-6]。蔷薇属植物是最先吸引了细胞学家注意力的庭园植物之一[7],从林奈时期即对其进行了染色体研究[8]。从1904年Strasburger对蔷薇属染色体基数的报道至今,全世界对其胞核学的研究已有很多[8-9],但鉴定蔷薇属的每一条染色体仍很困难。这是因为多种蔷薇属植物的种子萌发和枝条扦插较困难,常用嫁接方法进行繁殖,植株根系实为砧木的根系,因此一般选用茎尖为制片材料,而茎尖相对于根尖细胞分裂率低、细胞内含物较多、细胞壁较厚[10],更为主要的原因是蔷薇属植物的染色体为小染色体(平均长度 2.81 μm)[11-13],导致难以稳定地得到高质量的荧光原位杂交图谱[14],使荧光原位杂交技术在蔷薇属中的应用受到一定的限制。此外,由于可利用的蔷薇属染色体特异标记有限,利用荧光原位杂交技术进行染色体研究、使每一条染色体有特定的标记而便于识别的工作还无法展开。
笔者综述了染色体荧光原位杂交技术在蔷薇属上的应用情况,展望了其在该属植物研究中的应用前景,旨在为蔷薇属植物的分子细胞遗学研究提供参考。
1 特定DNA序列定位
通过rDNA在染色体上的定位,能够清楚地识别形态相近的染色体和核型相似的物种。根据rDNA位点数目、位置和拷贝数的变化,可以揭示染色体结构的变异,有利于分析物种亲缘关系、多倍体的形成等问题,并将细胞遗传学研究深入到分子水平[1]。蔷薇属中已经定位的序列只有高度重复序列,包括45S rDNA和5S rDNA。
1.1 45S rDNA
45S rDNA在细胞分裂间期大量转录表达形成特殊的区域,参与核仁形成。每个45S rDNA转录单位包括18S rDNA、5.8S rDNA、26S rDNA 3个亚单位和中间的间隔区序列(internal transcribe spacer,ITS),呈高度保守状态,在基因组中为串联重复序列,不同物种中的拷贝数为500~40 000。至今,45S rDNA在25个蔷薇属野生种/变种、12个古老月季品种及6个现代月季品种染色体上的位点数目和位置已有报道(表1)。
Ma等于1997年首先将荧光原位杂交技术应用到蔷薇属植物上。她用从大豆(Glycinemax)中克隆到的18-26S rDNA做探针,进行了月季花(R.chinensis)、香水月季(R.odorata)、Rosa×fortuniana、金樱子(R.laevigata)、刺梨(R.roxburgii)和1个现代品种(Rosa‘Angel Face’)的FISH分析,首次提出用18-26S rDNA重复序列进行蔷薇属植物的染色体荧光原位杂交是可行的,并指出rDNA杂交位点对应于核仁组织区(NOR),具有相对保守的数量和位置,即1个染色体组有1个NOR,且NOR都位于亚中着丝点染色体的短臂近端部[10]。随后,Fernández-Romero 等也用来自大豆的 18-25S rDNA做探针,研究了5个2倍体蔷薇野生种大花香水月季(R.gigantea)、麝香蔷薇(R.moschata)、野蔷薇(R.multiflora)、玫瑰(R.rugosa)、R.sempervirens和2个古老品种(R.chinensis‘Semperflorens’,R.gallica‘Versicolor’)的rDNA定位[14]。5个2倍体蔷薇野生种的结论与Ma等的一致,但古老品种R.chinensis‘Semperflorens’有3个杂交位点,分布于3条近似的染色体上,而R.gallica‘Versicolor’有6个杂交位点,位于3对不同的染色体上。Akasaka等以从小麦(Triticumaestivum)中获得的45S rDNA做探针,分别对属于A型染色体组[21-22]的单瓣月季花(R.chinensisvar.spontanea)、大花香水月季、麝香蔷薇、野蔷薇、光叶蔷薇(R.wichuriana),B型染色体组的小叶蔷薇(R.willmottiae),C型染色体组的玫瑰和D型染色体组的R.marretii、R.foliolosa进行了45 rDNA定位[11-12],其中A型染色体组的蔷薇最为重要,创造现代月季的8个野生种染色体全部属于该染色体组[23]。A型染色体组的这5个蔷薇野生种的45S rDNA杂交位点的数目和位置都比较固定,都有2个杂交位点,都位于随体染色体也是最短染色体(chromosome 7)的短臂端部,显示了在该染色体上的45S rDNA等位基因座。其他3个染色体组的蔷薇野生种的45S rDNA的位置也与A型染色体组的种类近似,位于最短染色体的短臂端部;但R.foliolosa(D型染色体型)的45S rDNA杂交位点数为6,且与同染色体组的R.marretii核型差异较大。Flory认为蔷薇属植物起源于亚洲东部,其中某些种逐渐迁移到北美洲[24],因此Akasaka等推测可能在R.foliolosa迁移到北美的过程中其染色体发生了重组,形成了因地理分布造成的核型变异[12]。Lim等用从小麦中克隆的 18-5.8-26S rDNA为探针检测到5倍体的狗蔷薇(R.canina)有5个45S rDNA杂交位点,但未明确位点的准确位置[17]。2014年张婷等用来自番茄(Solanumlycopersicum)的45S rDNA为探针进行了桂味组(Section Cinnamomeae)的多苞蔷薇(R.multibracteata)、合柱组(Section Synstylae)的川滇蔷薇(R.soulieana)及金樱子组(Section Laevigatae)的金樱子(R.laevigata)的原位杂交研究,结果表明45S rDNA的位点数相对保守,这3个2倍体的蔷薇品种都只有1对45S rDNA位点,且大都位于1对异型同源sm染色体的短臂上,其中多苞蔷薇和川滇蔷薇的2个45S rDNA位点信号强弱有区别,而金樱子的杂交信号强弱一致[16]。
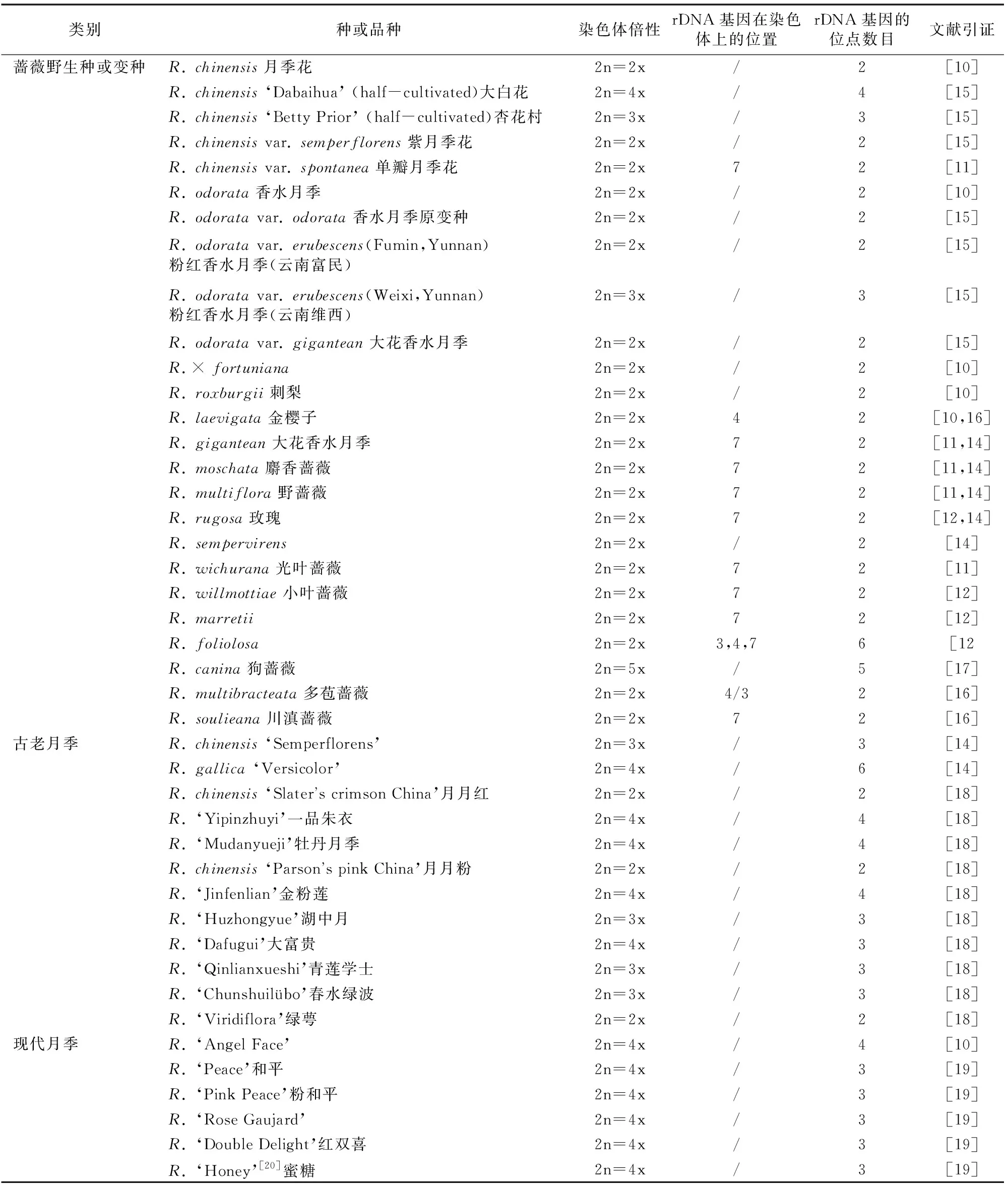
表1 45S rDNA基因在蔷薇属43个种或品种染色体上的定位及位点数目
田敏等分别于2012年和2013年用来自蕃茄的45S rDNA对紫月季花(R.chinensisvar.semperflorens)、大白花(R.chinensis‘Dabaihua’)和杏花村(R.chinensis‘Betty Prior’),香水月季复合群(R.odoratacomplex)的香水月季原变种(R.odoratavar.odorata)、大花香水月季(R.gigantea)、来自云南富民的2倍体粉花香水月季(R.odoratavar.erubescens)和来自云南维西的3倍体粉花香水月季(R.odoratavar.erubescens)[15],月月红(R.chinensis‘Slater’s crimson China’)、一品朱衣(R. ‘Yipinzhuyi’ )、牡丹月季(R. ‘Mudanyueji’ )、月月粉(R.chinensis‘Parson’s pink China’)、金粉莲(R. ‘Jinfenlian’)、 湖中月(R. ‘Huzhongyue’ )、大富贵(R. ‘Dafugui’ )、青莲学士(R. ‘Qinglianxueshi’)、春水绿波(R. ‘Chunshuilübo’)、绿萼(R. ‘Viridiflora’)等10个中国古老月季品种[18],以及和平(R. ‘Peace’)、粉和平(R. ‘Pink Peace’)、R. ‘Rose Gaujard’ 、红双喜(R. ‘Double Delight’)、蜜糖(R. ‘Honey’)等5个现代月季品种[19]处于细胞分裂间期和前中期的核进行了荧光原位杂交研究。结果表明,从月季组的蔷薇野生种到中国古老月季品种的形成过程中,45S rDNA位点数与多倍化过程紧密相关且与倍性成正比,中国古老月季的45S rDNA的数量和位点与原产中国的野生种一致;但在现代品种中的45S rDNA位点数减少了,均只有3个。该研究在一定程度上表明中国古老月季是由原产中国的一些野生种经长期的杂交或自然突变,经由人工选育固定下来而形成的,相关的核型分析[25]和SSR分子标记[26]也支持这一观点;而现代月季渗入了欧洲的一些蔷薇属种质,这种远缘杂交引起了染色体更大的变异,导致rDNA的数量发生较大的变化。此外,原产中国的月季花复合群、香水月季复合群及古老品种,不论是在染色体的倍性和形态上[25,27],还是在45S rDNA位点的拷贝数和间期组织模式上,都表现出多样性,而现代栽培品种却表现出一致性。
总体来说,虽然目前对蔷薇属植物的45 rDNA定位研究主要还是集中在数量上,对其在染色体组上的精确定位较少,但从已有的研究可知,除R.foliolosa、古老品种R.gallica‘Vesicolor’和大富贵外,45S rDNA的位点数在蔷薇野生种和古老品种中均对应于核仁组织区,具有比较保守的数量和位置,即1个染色体组对应1个45S rDNA位点,且都位于亚中部着丝点染色体的短臂近端部。现代品种的45S rDNA位点数则表现出与野生种或变种和古老品种不同的变化规律:除R. ‘Angel Face’外,其他5个现代品种的45S rDNA少于其对应的染色体组数,且在拷贝数和间期组织模式上都表现出一致性,表明反复的品种间杂交和自交已使染色体结构渐渐趋于同质。因此,要进一步提高现代月季的品质需要从野生资源[28]和古老月季[29]中引入新的种质。
1.2 5S rDNA
5S rDNA是绝大多数生物的核糖体大亚基的一个组成部分,其功能在真核生物中还不太清楚。5S rDNA基因家族通常以高拷贝串联重复形式存在于基因组中,有2 000~5 000 个拷贝,且常不与其他重复序列(如26S-5.8S-18S)串联在一起。每个5S rDNA重复单位由基因编码区和非编码区(间隔区)组成,长度为200~1 000 bp。编码区长度为 120 bp,非编码区长度为100~700 bp。120 bp的编码区相对间隔区序列比较保守[30],间隔区序列差异很大,位于间隔区中部的序列变化最丰富[31]。相对于45S rDNA,5S rDNA应用于蔷薇属植物的荧光原位杂交有一定的困难,到目前为止仅对13个蔷薇野生种进行了5S rDNA的定位(表2)。
Ma等首先研究了5S rDNA应用于蔷薇属植物染色体荧光原位杂交的可行性。她用克隆自黑木相思树(Acaciamelanoxylon)的5S rDNA做探针没有获得成功,但指出来自黑木相思树的5S rDNA既能杂交到与豆科植物亲缘关系较远的单子叶植物,也能杂交到与之亲缘关系相对较近的梨属植物上[10],此次试验未成功不是由于原位杂交的程序有误或是5S rDNA序列的差异性,而是由于染色体制片的问题。不过自此后,蔷薇属植物的5S rDNA原位杂交的探针都是克隆自靶染色体材料的基因组本身。
Akasaka等从野蔷薇中克隆了5S rDNA,制备成探针对属于A型染色体组的单瓣月季花、大花香水月季、麝香蔷薇、光叶蔷薇,B型染色体组的小叶蔷薇,C型染色体组的玫瑰和D型染色体组的R.marretii、R.foliolosa进行了rDNA定位[11-12]。A型染色体组的5种蔷薇的5S rDNA杂交位点数目和位置的变化较大,反应了染色体重组现象。Wu 等认为这5个种的同源性较高[32-33],而它们最短染色体上都同时有45S和5S rDNA位点,这可以作为A型染色体组蔷薇的染色体特征标记;5S rDNA杂交位点除位于最短染色体以外,其余位点都位于其他染色体长臂的相近区域。在B、C、D染色体组的蔷薇中,5S rDNA的杂交位点数目比较固定,位于染色体长臂上,且与45S rDNA不在同一条染色体上。综合来看,这9个2倍体蔷薇中,5S rDNA位点数有2个(1对)、3个和4个(2对),其中具1对5S rDNA位点的种,其5S rDNA位点位于非45S rDNA染色体长臂的近着丝点处;具3个或2对的种有1对5S rDNA位点与45S rDNA位于同一染色体的长臂或短臂上,其余位点则位于另外染色体的长臂的近着丝点处。Lim 等用来自狗蔷薇的5S rDNA为探针与自身的染色体进行杂交[17],得到8个杂交位点,均位于染色体长臂的近着丝点处,其中3个位点与45S rDNA位于同一染色体,另外5个位点位于别的染色体上。
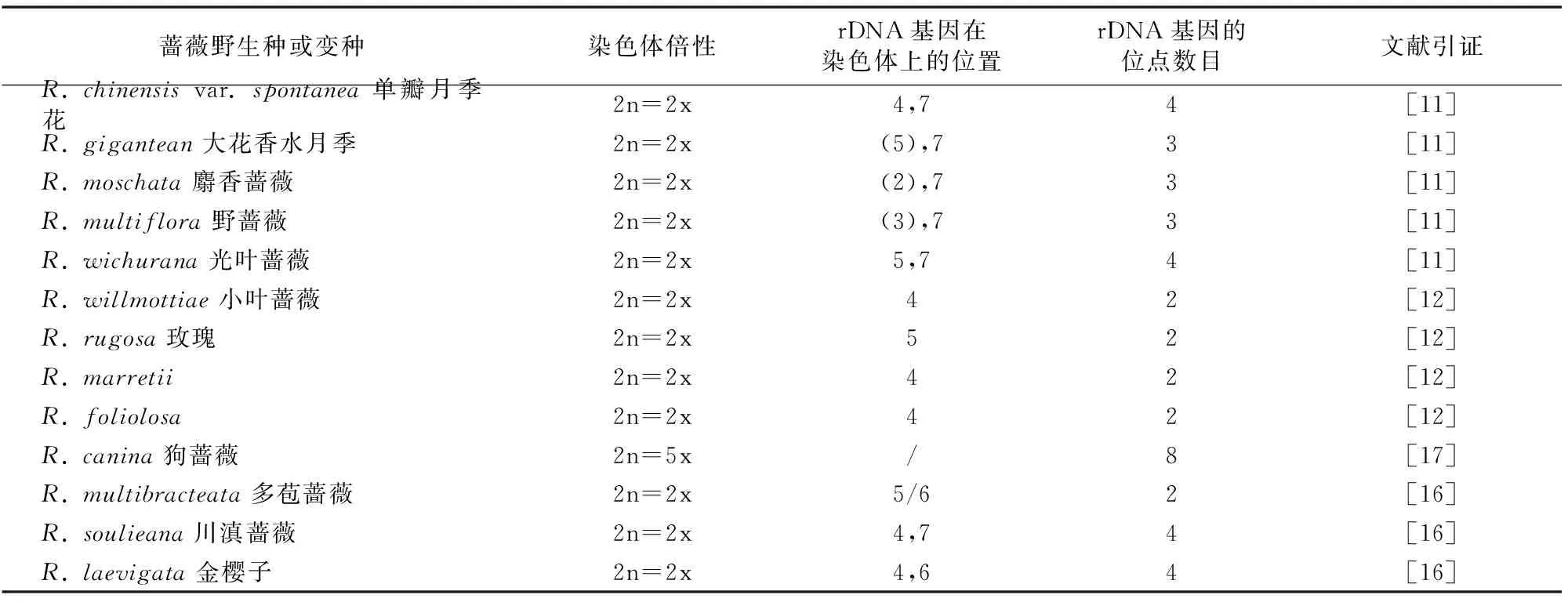
表2 5S rDNA基因在蔷薇属13个种染色体上的定位及位点数目
张婷等参照Akasaka等的方法[11-12],以PCR方法制备5S rDNA,用于桂味组的多苞蔷薇、合柱组的川滇蔷薇及金樱子组的金樱子的染色体荧光原位杂交研究,得出3个2倍体野生种的5S rDNA位点数量不等,但都位于染色体长臂的近着丝点处,杂交信号的强弱有差异[16]。具体来看,多苞蔷薇的1对5S rDNA位于非45S rDNA染色体的长臂的近着丝点处;川滇蔷薇和金樱子有2对5S rDNA位点,其中1对位于45S rDNA染色体的长臂的近着丝点处,另1对位于其他染色体长臂的近着丝点处。由此得出,蔷薇属植物中5S rDNA位点按是否与45S rDNA有共线性分为2种:只有1对5S rDNA位点,与45S rDNA没有共线性,且位于染色体长臂的近着丝点处;有3个、2对或更多5S rDNA位点,与45S rDNA共线性的5S rDNA位点位于染色体短臂或长臂的近着丝点处,其余位点位于其他染色体长臂的近着丝点处。该结论与Akasaka等的[11-12]基本一致。
因此,不同于45S rDNA,5S rDNA的位点数在蔷薇属不同种中变异较大,有2、3、4、8个不等,且没有一定的规律,在染色体上也没有固定的分布模式。目前尚未见有对蔷薇属古老品种和现代品种的5S rDNA定位的报道。
2 染色体同源性分析
荧光原位杂交方法已在很多植物中普遍应用,但由于蔷薇属植物的染色体较小,荧光原位杂交技术的普遍应用有一定难度,因此,应用其进行分子细胞遗传学的深入研究较少,目前主要应用于染色体的同源性及亲缘关系的初步推测上。
在Ma等的研究中,R.×fortuniana分裂间期时2个NOR位点在大小上是相同的,但其中1个NOR位点较分散也较活跃[10],这与Shepherd认为这个种为杂交起源(R.banksiae×R.laevigata)[34]的观点相吻合,因此推测其为杂种起源。同时,现代品种R. ‘Angel Face’(2n=4x)的染色体组有4个 18-26S rDNA 杂交位点,但其中2个位点信号强度较强,表明其由AABB 2种异源染色体组成,且比例为2 ∶2。
Fernández-Romero等用18-25S rDNA的原位杂交结合小孢子母细胞(pollen mother cells,PMCs)减数分裂染色体制片、花粉育性观察和实际结实率统计的方法,分析了古老品种R.chinensis‘Semperflorens’和R.gallica‘Versicolor’各自的染色体同源性[14]。R.chinensis‘Semperflorens’的3个18-25S rDNA杂交位点分布于3条近似的染色体上;PMCs减数分裂时最多形成6个3价体和3个1价体,这两方面的证据表明R.chinensis‘semperflorens’的染色体之间具有很高的同源性,为自发形成的同源3倍体,这种同源性会导致在减Ⅰ后期时染色体分配出现较多的错误从而导致不育;花粉育性观察和结实率统计支持此结论。而R.gallica‘Versicolor’的6个18-25S rDNA杂交位点位于3对不同的染色体上,其中位于1对较小的染色体上的信号较强,结合nrDNA的ITS1序列研究的结果[35],表明R.gallica‘Versicolor’为异源起源,其中一个染色体组含有1个NOR,另一个染色体组含有2个NOR。PMCs减数分裂时最多形成14个2价体,也证明其异源性,在减Ⅰ后期时染色体能正常分配使其可育;花粉育性观察和结实率数据符合此结论。
狗蔷薇为5倍体,它减数分裂时特殊的配子不均等分裂方式很早引起了学者的极大兴趣[8,21]。Lim等用FISH技术辅助分析狗蔷薇不均等减数分裂中的染色体分配情况。他用18-5.8-26S rDNA和5S rDNA为探针,分别与体细胞和PMCs进行双色FISH,证明了狗蔷薇中只有1对同源染色体,在形成配子时此同源染色体均匀分配到雌雄配子中,其他一直处于单价体形式的染色体都分配到雌配子中[17]。该研究为利用FISH深入研究蔷薇属植物的染色体同源性及其配子形成时的分配方式提供了参考。
张婷等基于rDNA-FISH的核型分析研究发现多苞蔷薇有2种核型类型[16],一种与Jian等常规核型分析的结果相同,属较为原始的1A型,另一种为更进化的2A型[37]。前者没有异型同源染色体,且5S rDNA位点染色体不是sm染色体;后者的2对rDNA染色体都是异型同源染色体。这可能是由于研究材料来自不同居群,它们之间存在核型差异,这也从细胞水平上反映了其种内存在丰富的形态变异和遗传多样性。川滇蔷薇的核型及核型公式与Jian等基于常规核型分析的结果[36]相同,但2对sm染色体的序号相差较大,这一方面可能是由于研究材料来源不同所致,另一方面也可能是由于常规核型分析的精确度不高导致染色体排序有误。金樱子在核型和核型公式上与Crane等基于常规核型分析的结果相同[37],但在染色体排序上稍有差别。这3种蔷薇野生种均存在异型同源染色体,异型同源染色体由于较大的长度差异使研究人员在常规染色体核型分析时很容易将其错配。因此,基于rDNA-FISH的蔷薇属植物染色体核型分析更加准确可靠,应采用类似的较精确的方法对一些分布较广、种内存在丰富形态变异的蔷薇进行进一步的核型研究。由此可见,荧光原位杂交技术不仅能准确地识别各自染色体组中的同源染色体,而且还能通过杂交位点数量、位置和强弱体现各自染色体的结构特征,为染色体识别提供了明确有效的标记。
3 原位杂交技术在蔷薇属植物研究中的应用前景
作为分子遗传学与细胞遗传学结合而产生的一门新兴技术,原位杂交具有上述2个学科所不具备的许多优点,它不仅能够将核酸序列直接定位在染色体或间期核中的特定位置,以构建染色体的物理图谱,分析染色体和基因组的结构,而且能对伴随物种形成过程中的基因组内及基因组间的结构重排进行探测[38]。在其他栽培作物中,利用特定DNA序列、大片段克隆(BAC/YAC)和整个基因组DNA(gDNA)作为探针,研究多倍体的起源、不同基因组之间的关系、DNA物理图谱、染色体结构变异等的报道已有很多,但在蔷薇属植物的染色体结构变异检测、基因定位、外源染色体鉴定及亲缘关系研究上,FISH技术都还没有得到充分应用。蔷薇属植物分布广泛、种类众多、分类复杂,如何应用成熟的FISH技术来解决复杂的遗传和进化问题,也就成为了蔷薇属植物分子细胞遗传学研究的方向。针对目前的研究现状,可将FISH应用于以下3个方面。
3.1 现代月季形成过程中的染色体变异
作物驯化可以看作是植物进化的一种模型,在驯化过程中随着农业需求性状的选择育种,染色体、DNA结构会随之改变[39]。蔷薇属植物有着悠久的栽培历史,经过长期自然和人工栽培的变异、杂交、选择,经历了从蔷薇野生种(wild roses or species roses,WR or SR)演化到古老月季(old garden roses,OGR),进而到现代月季(modern roses,MR)的过程。我国的蔷薇属资源是创造现代月季的重要原始亲本。原产我国的月季花复合群(R.chinensiscomplex)和香水月季复合群(R.odoratacomplex)是现代月季最重要的两大性状即连续开花和芳香怡人(茶香)的来源[23]。我国古老月季如月月红、月月粉、彩晕香水和淡黄香水等,对现代月季的育成起到了决定性的作用[40],是野生蔷薇和现代品种之间的过渡类型[6,41]。现代月季来自我国的原始野生亲本均是2倍体[34,42],而现代月季品种通常是4倍体[43];Heinisch等还推测在现代月季的杂交育种中出现过更高倍(大于4倍)的杂种[44]。那么在从蔷薇野生种演化到古老月季,进而演化到现代月季的过程中,染色体在结构上发生了怎样的变化?对现代月季及其重要历史亲本进行系统的以染色体荧光原位杂交为基础的分子细胞遗传学研究,是回答这些问题的前提和必要途径。
3.2 蔷薇属植物的染色体变异及分布规律研究
蔷薇属约有200多个种[5],由于缺乏遗传背景的详尽研究,制约了其在现代月季遗传改良中的深入应用。早期的研究认为自然界中野生种存在2x至8x的倍性[21-22],最近在分布于我国云南的野生种中发现了6x和10x[36,45]却没有发现4x和8x,并且多倍化主要集中在分布于高海拔地区的桂味组植物中,总体比例不到10%。根据已有的关于蔷薇属植物的染色体报道可知,除分布在欧洲的狗蔷薇组(Sect.Caninae)和蔷薇组(Sect.Rosa)、高海拔和极地周围的桂味组以及北美的卡罗莱纳组(Sect.Carolinae)中多倍体种占一定比例外,绝大部分蔷薇野生种均为2倍体。因此,蔷薇属植物进化和演化的细胞学动力主要还是来自染色体的重组和结构变异。那么,蔷薇属植物是否存在比10x更高的倍性?组间和组内各种间基于FISH的亲缘关系如何?多倍体和染色体结构变异与生境特别是海拔的关系如何?系统演化过程中染色体重组和结构变异各自扮演了怎样的角色?利用荧光原位杂交技术对这些问题进行深入研究,是研究蔷薇属植物的遗传多样性和系统演化的必要细胞学手段,也可为利用这些野生资源对现代月季品种进行种质创新和遗传改良提供细胞学数据。
3.3 蔷薇属植物多倍体的起源研究
植物多倍化是一种普遍存在的生物学现象,多倍化是植物进化和多样化的重要原动力[46-47]。对于植物多倍体,FISH和GISH(基因组荧光原位杂交技术)是探测祖先基因组来源、理清种间进化关系、构建遗传图谱、分析基因组杂交渐渗和促进遗传育种的重要手段[48]。常用的方法有单亲本GISH[49]、双亲本GISH[50]、基因组特异探针[51]并结合减数分裂时染色体的配对行为[14]等方法来判断多倍体基因组的起源和组成,特别是GISH为鉴别外源染色体带来了方法上的革命[52]。虽然整个蔷薇属植物系统演化的主要细胞学动力可能不是多倍化,但多倍化现象在狗蔷薇组、桂味组及卡罗莱纳组中却十分显著。那么,这些多倍体是如何形成的?是多倍化(同源多倍化和异源多倍化)还是由原始染色体断裂产生?哪些种参与了它的形成并承担了什么角色?要解答这些科学问题,需要用到分子细胞遗传学的研究方法,可以利用与多倍体蔷薇有较近亲缘关系且分布区重叠的其他蔷薇的基因组为探针进行GISH杂交,并结合减数分裂时染色体rDNA-FISH信号进行分析,阐明这些多倍体蔷薇的起源和形成过程,为研究蔷薇属植物的系统演化提供分子细胞遗传学证据。
猜你喜欢
作物学报(2022年2期)2022-11-06
中国烟草学报(2021年3期)2021-08-04
基层中医药(2020年6期)2020-09-11
人大建设(2018年7期)2018-09-19
上海农业学报(2017年3期)2017-04-10
特产研究(2016年3期)2016-04-12
中国当代医药(2015年17期)2015-03-01
郑州大学学报(医学版)(2015年1期)2015-02-27
火花(2015年7期)2015-02-27
现代检验医学杂志(2014年4期)2014-02-02
