
小麦TCP基因家族的全基因组鉴定和对热胁迫的响应
2018-12-13 04:59李梓彰党仁美刘媛媛闻珊珊
麦类作物学报 2018年10期
丁 宁,李梓彰,田 文,党仁美,刘媛媛,闻珊珊
(西北农林科技大学农学院,陕西杨凌 712100)
TCP蛋白作为植物特异性转录因子,分别以玉米中 teosinte branches1(tb1) 、金鱼草中cycloidea(cyc)和水稻中PCF三个基因的首字母命名,在植物细胞增殖和生长过程中发挥着重要作用[1]。许多物种的TCP基因家族已经被鉴定[2-5],它可以参与不同激素的信号通路,介导对赤霉酸、细胞分裂素、脱落酸、茉莉酸、油菜素内酯和吲哚-3-乙酸的活性调控[6]。
TCP蛋白由59个氨基酸组成的碱性螺旋I-环-螺旋II(bHLH)结构组成,有I类(也称为PCF或TCP-P类)和II类(也称为CYC/TB1和CIN或TCP-C)两个家族。I类亚家族中的PCF1和PCF2蛋白[7]可以同源或异源二聚体的形式与增殖细胞核抗原(PCNA)的水稻启动子元件特异性结合,而PCNA是与DNA复制和细胞周期控制相关的基因[8]。II类亚家族可进一步细分为CIN和CYC/TB1两个分支。CIN和CYC/TB1(或ECE)中的基因分别参与外侧器官发育和腋生分生组织的发育,从而分别产生花或侧枝[2]。
作为DNA结合蛋白,一些TCP基因对叶绿体或核基因的转录也有调控作用,它们可以自我激活或与其他基因的顺式元件相互作用的方式调节转录[9-10]。此外,植物TCP类转录因子还参与分枝、配子、叶和花的发育以及株高、昼夜节律的调控等生理活动[11]。黎飞飞等[12]在郑麦7698中通过电子克隆得到水稻 TCP-1的同源基因 TaTCP-1A,并通过基因枪介导的遗传转化方法证明该基因在一定程度上可以促进小麦愈伤组织的再生。Yuji Yamasaki等[13]研究发现,不管小麦种子是否处于休眠或者其存在大小、重量方面的差异,TCP可以在转录后水平调控种子胚的成熟,进一步预测得知SITEIIATCYTC被证明在种子发芽中与蛋白质合成相关基因的表达上调有关。Song等[14]通过对小麦幼穗进行0 ℃处理,鉴定到192个microRNAs,其中TCP和其他转录因子一起在冷胁迫中靶向特异的miRNA表达而导致异常的生殖器官发育。
高温会直接影响小麦的发育和籽粒品质。据报道,温度升高2 ℃以上可能导致小麦产量降低50%。为保证小麦在高温条件下稳定高产,开展小麦耐热基因的鉴定和应用研究有很重要的现实意义。小麦基因组数据的不断完善和释放,使得在全基因组水平对小麦基因进行鉴定和分析成为可能。目前,对小麦TCP基因在全基因组水平的研究尚未见报道,因此,本研究利用生物信息学方法和qRT-PCR技术在全基因组水平对TaTCP基因的结构、系统发育、进化压力选择指标和RAN-seq表达模式等进行研究,并对候选耐热基因进行荧光定量验证,以期为小麦TCP基因的进一步研究奠定基础。
1 材料与方法
本研究所用材料和方法详见田文等[15]的“普通小麦DREB基因家族的全基因组鉴定及热胁迫下的表达模式分析”一文。qRT-PCR分析中所用的特异性引物见表1。

表1 荧光定量PCR的引物Table 1 Primers for qRT-PCR
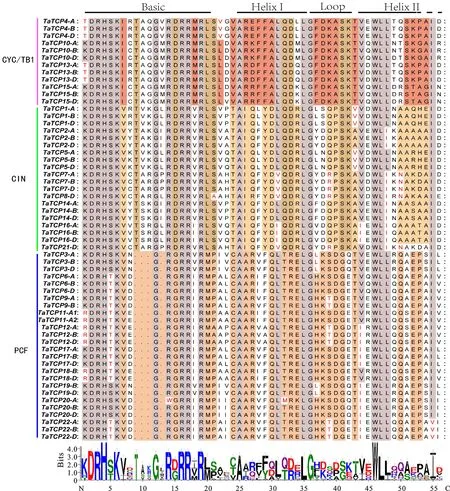
青色代表这三个亚家族中的保守残基;红色代表具有类似电荷或疏水性的氨基酸;棕色代表在CYC/TB1和CIN蛋白中保守的残基,酒红色表示只在CYC/TB1蛋白中,淡黄色表示仅在CIN蛋白中,浅红色表示在PCF蛋白质中。对应于序列比对结果下方的字母高度反映了该处氨基酸出现的相对频率。
Lilac colour boxes represent the conserved residues in these three classes; amino acids with similar charge or hydrophobicity are in red; residues conserved in CYC/TB1 and CIN proteins are shown in brown; residues only conserved in CYC/TB1 proteins are shown in wine red; residues only conserved in CIN proteins are shown in light yellow; residues conserved in PCF proteins are shown in light red. The height of the letter below the sequence alignment result reflects the relative occurrence frequency of the amino acid at that position.
图1TCP氨基酸序列的比对
Fig.1AlignmentofTCPmotifsinputativeaminoacidsequences
2 结果与分析
2.1 小麦中TCP基因家族的候选基因鉴定
本研究最终得到58个非冗余且包含完整结构域的小麦TCP候选基因。通过在线工具计算后统计,58个TaTCP的蛋白质序列长度介于177~577 aa,分子量为18.2~61.6 kD,等电点pI介于4.95~11.66之间,开放阅读框ORF长度为587~1 734 bp,这些都说明小麦TCP基因家族成员之间存在较大的组成差异。然后结合染色体位置和系统发育树将候选基因命名。发现58个TaTCP基因分布在除6A以外的其余20条染色体上,并且有20个基因位于5号染色体中,占基因总数的34.48%,说明5号染色体在TaTCP基因的进化中具有重要作用。然而由于小麦基因组的详细注释还没有完成,所以TaTCP基因在染色体中的确切位置还不清楚。
为了验证本研究对候选TaTCP基因的预测,根据Cubas(1999)提出的方法[1],对蛋白序列中的保守基序进行了进一步研究。结果显示(图1),在TCP-C亚家族中,bHLH中碱性区域含有一个双向核定位信号(NLS),该信号对于蛋白质向核内的转运是必需的,而TCP-P亚家族中的碱性区域只包含部分NLS,推测这两个亚家族之间的蛋白质序列差异可能导致其功能上的不同。TCP-C和TCP-P亚家族分别在碱性区域中都含有保守基序KDRHSK(I/V)-TG-RDRRΨ和(K/R)DRH-KV-G-RGRRΨ,其中Ψ代表任何疏水残基,如甲硫氨酸(M)、异亮氨酸(I)。在两个亚家族中,连接两个螺旋的区域具有相同的保守甘氨酸(G)、天冬氨酸(D)和丝氨酸(S)残基。
2.2 TaTCP的系统发育和基因结构分析
为研究小麦TCP基因的系统发育关系,采用N-J法构建系统发育树。结果(图2)显示,所有TCP成员分为TCP-C和TCP-P亚家族,TCP-C亚家族又分为两个小亚类。此外,发现系统发育树中有些数值不是很高,这可能是由于全长蛋白质序列的相似性低造成的。相当多的TaTCP成员(89.83%)不含内含子。只有6个基因( TaTCP6-D、 TaTCP8-D、 TaTCP17-B、 TaTCP20-A、 TaTCP20-B和 TaTCP22-A)在编码序列中含有一个内含子,可能是基因在进化过程中产生了结构上的变化。而两个亚家族之间具有相似的基因结构,表明小麦TCP基因家族在进化上存在一定的保守性,这种结构上的稳定性可能与TCP参与植物多种重要的生物学过程密切相关。为了显示进化关系,根据58个TaTCP、24个AtTCP和22个OsTCP的全长蛋白质序列构建了系统发育树(图3)。58个TaTCP中有26个在PCF亚家族,20个在CIN亚家族,12个在CYC/TB1亚家族,预测TaTCP基因家族可能与相同分类的直系同源基因具有相似的生物学功能。
2.3 TaTCP的达尔文压力选择分析
达尔文进化选择的类型通常通过计算Ka/Ks(非同义替换率/同义替换率)之间的比值来衡量,其中Ka/Ks=1用于中性选择;Ka/Ks<1表示纯化选择;Ka/Ks>1表示正向选择[16]。乌拉尔图小麦(T.urartuas)和节节麦(A.tauschii)分别作为小麦基因组中A和D基因组的供体物种,大麦也被认为与小麦在起源上存在一定的关联,它们都因为与小麦在进化上关系密切而被作为直系同源物种。通过序列比对的方法寻找小麦TCP的直系同源基因通过Ensembl Plants数据库最终确认其功能的相似性。通过命名的22组(每组含有1、2或3个等位基因)TaTCP基因分别在大麦、乌拉尔图小麦和节节麦基因组库中寻找对应的直系同源序列(orthologous)。结果显示,分别有19、3和2组TaTCP在A、B、D三个亚基因组中与大麦、乌拉尔图小麦和节节麦中存在直系同源基因(表2)。据统计,在小麦和大麦(MLOC_开头的ID号)组成的19组直系同源基因对中,Ka/Ks的比值范围为0.0613~0.9474,说明小麦基因相对于大麦进行了纯化选择。在小麦和节节麦(F775_开头的ID号)组成的2组基因对的Ka/Ks比值范围为0~0.3221,说明小麦基因相对于节节麦也进行了纯化选择。同样,小麦与乌拉尔图小麦(TRIUR3_开头的ID号)之间所组成的3组直系同源基因对的Ka/Ks值介于0.1660~1.2225之间,说明它们之间存在不同的选择压力,特别是 TaTCP16-A和 TRIUR3_19119的Ka/Ks比值为1.2225,反映了 TaTCP16-A在进化过程中的正向选择,这对小麦物种的进化是非常有利的。从寻找的直系同源基因个数看,乌拉尔图小麦或节节麦的数量小于大麦,这可能是因为两个物种(AA和DD)的基因组小于大麦基因组造成的。
2.4 TaTCP基因在小麦不同组织和不同逆境胁迫下的表达分析
用小麦(品种为中国春)5种组织或器官(穗、芽、根、叶和籽粒)和非生物胁迫反应(热和干旱)的RNA-seq数据来探究TCP基因在小麦生长和发育中的潜在生物学功能,发现大部分同源基因对表现出类似的表达模式。图4所示,在58个候选基因中,35个TaTCP在至少一种组织中检测到表达。总共有1、1、4和3个基因分别在籽粒、茎、根和穗中特异性表达。有些基因缺乏表达数据可能涉及特殊的时间和空间表达事件,所以没有在数据库中检测到。此外,58个TaTCP基因中没有一个基因在5种组织中同时检测到表达值。 TaTCP6-A在叶、根和茎中表达量最高,表明该基因可能参与植物特异性生长调节。对获取的表达数据分析发现, TaTCP5-D作为一个上调基因,对热和干旱胁迫的反应明显高于其他TaTCP基因。与对照相比,大部分基因表达值在两种胁迫条件下没有太大的变化,这可能是因为TaTCP基因位于调控网络中的上游位置,与非生物胁迫的应答有复杂的调控关系。

图2 TaTCP的系统发育关系(A)和基因结构(B)Fig.2 Phylogenetic relationships(A) and gene structure(B) of TaTCP

图3 小麦、水稻和拟南芥TCP的系统发育分析Fig.3 Phylogenetic analysis TCPs of wheat, rice and Arabidopsis表2 用大麦、乌拉尔图小麦和节节麦作为TaTCP直系同源基因计算的Ka/Ks比率Table 2 Ka/Ks ratio of TaTCP orthologous genes calculated with barley, T.urartuas, and A.tauschii as outgroups

小麦IDWheat gene ID直系同源基因IDOrthologous IDA基因组A-genome Ka/KsB基因组B-genome Ka/KsD基因组D-genome Ka/Ks TaTCP1 MLOC_702660.199 10.190 00.206 3 TaTCP2 MLOC_102140.472 10.490 90.466 6 TaTCP3 MLOC_723930.130 50.212 70.163 1 TaTCP4 MLOC_581420.217 70.238 10.251 6 F775_019230.287 20.322 10.000 0 TaTCP5 MLOC_147850.119 20.167 60.129 5 TaTCP7 MLOC_682850.096 90.104 40.093 9 TaTCP8 MLOC_12004--0.159 7 F775_07590--0.173 9 TaTCP10 MLOC_701160.265 40.290 60.258 5 TRIUR3_317950.471 70.321 50.324 8 TaTCP13 MLOC_46140.230 80.205 10.229 7 TRIUR3_207280.315 80.166 00.206 7 TaTCP14 MLOC_639890.947 40.925 00.945 8 TaTCP16 MLOC_725810.157 40.216 60.132 1 TRIUR3_191191.222 50.740 70.799 7 TaTCP18 MLOC_49381-0.424 20.346 1 TaTCP19 MLOC_23537-0.582 10.598 7 TaTCP22 MLOC_605770.061 30.068 70.066 7

红色表示基因的表达较强,白色表示中间表达量,蓝色表示没有表达信号。1和2:37 ℃下热胁迫处理1 h和6 h;3和4:干旱胁迫处理1 h和6 h。
The red color represents the higher expression of relative gene; white indicates intermediate expression value and the blue color indicates no expression signal. 1 and 2:Treatment for 1 h and 6 h under 37 ℃ heat stress,respectively; 3 and 4:Treatment for 1 h and 6 h under drought stress, respectively.
图4TaTCP基因在五种不同器官或组织中以及在非生物胁迫下的表达谱
Fig.4ExpressionprofilesofTaTCPgenesinfivedifferentorgansortissuesandunderabioticstresses
基于基因表达热图,选择 TaTCP5-A、 TaTCP5-B和 TaTCP5-D三个基因作为研究对象,使用qRT-PCR技术在35 ℃和42 ℃条件下进一步验证热图的结果。结果显示,在小麦不同的生长发育时期内,三个部分同源基因在不同温度下的响应是不同的。从图5中看出, TaTCP5-A在12个不同时期和两种热胁迫处理下的表达量基本都是下调的,这与热图中的结果一致;但在分蘖期35 ℃的热胁迫下和在开花期42 ℃的热胁迫处理下,该基因都有明显的的上调,可能在两个不同时期, TaTCP5-A在抵御热胁迫中发挥作用。 TaTCP5-B在灌浆早期35 ℃的胁迫下表达量上升,说明在灌浆早期该基因是发挥主要的耐热功能的因子;而其表达量在42 ℃胁迫下是下降的,说明即使处于同一生育时期,在不同的温度下基因的表达量都是不同的。而在热图中表现明显的 TaTCP5-D,在分蘖-起身期、拔节期、拔节后期、孕穗前期和抽穗期,在35 ℃的处理下表现尤为明显,特别是在抽穗期间比对照表现出更高的表达水平,表明在这些发育时期, TaTCP5-D是发挥耐热功能的主要因子。通过上述分析,qRT-PCR结果与RNA-seq数据在趋势上基本一致,也证明了使用RNA-seq数据评估小麦在不同处理下的转录表达水平是合理且可行的(图5)。
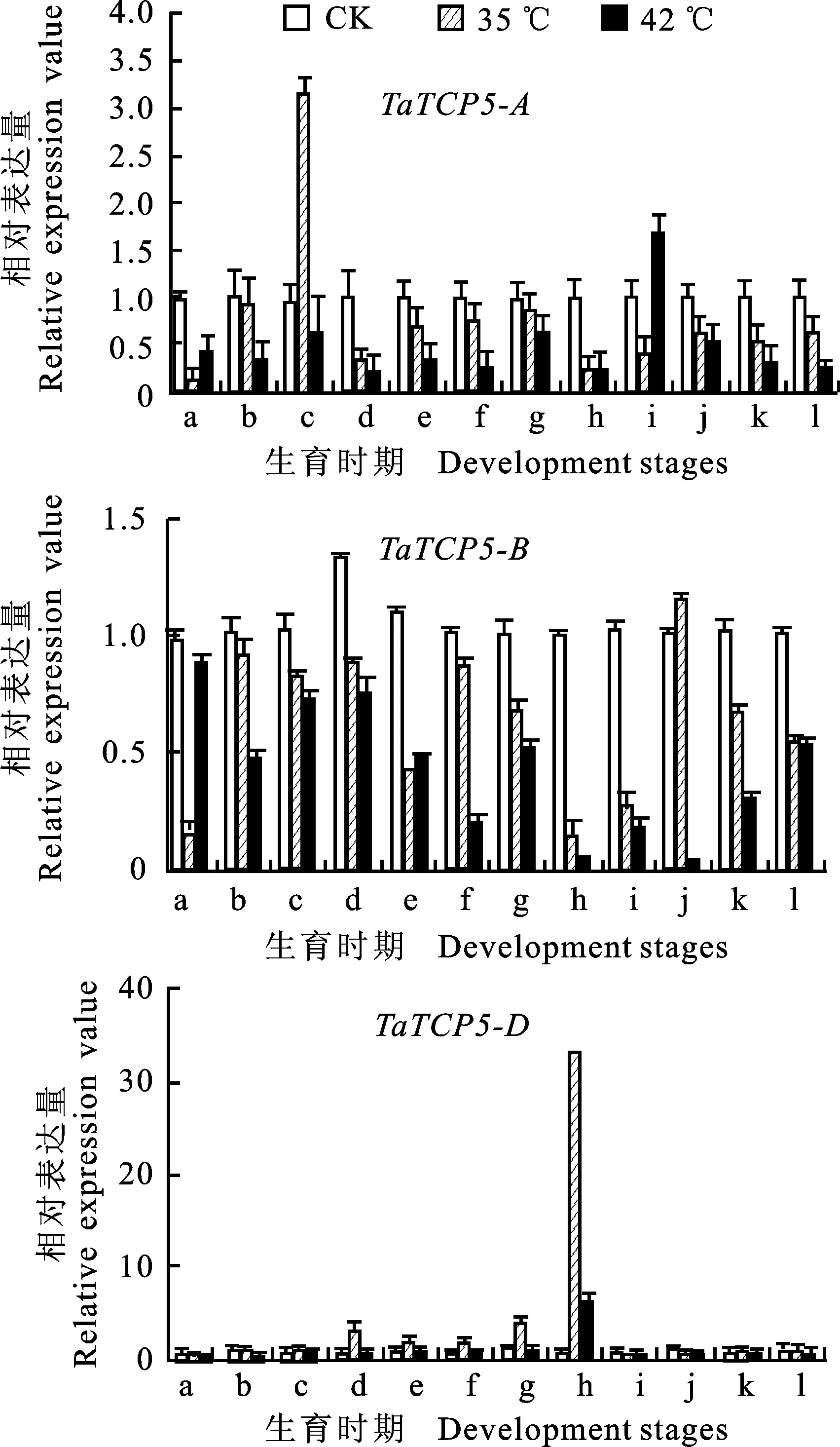
A:出苗3天;b:出苗7天;c:分蘖期;d:分蘖-起身期;e:拔节期;f:拔节后期;g:孕穗前期;h:抽穗期;i:开花期;j:灌浆早期;k:灌浆期;l:灌浆后期。
A:3 days after germination; b:7 days after germination; c:Tillering; d:Erecting; e:Early jointing; f:Late jointing; g:Early booting; h:Heading stage; i:Flowering; j:Early stage of grain filling; k:Filling; l:Late grain filling stage.
图5TaTCP5-A/B/D基因在小麦不同发育时期热胁迫下的表达水平
Fig.5ExpressionlevelsofTaTCP5-A/B/Datdifferentgrowthstagesunderheatstress
3 讨 论
根据PlantTFDB数据库得知,T.urartu和Aegilopstauschii分别只有4个和5个鉴定到的TCP基因,而本研究中小麦被鉴定到58个非冗余的候选基因。这58个候选基因组可以划分成22组,除 TaTCP8-D和 TaTCP21-D外,其余20组均含有两个或三个等位基因。由于它的供体物种都只有少量的TCP基因,所以推断现在小麦中的TCP基因是随着其进化经历了基因组的杂交和融合才形成的。有趣的是,其中 TaTCP11-A1和 TaTCP11-A2这两个基因位于A基因组的同一条染色体上,除了区段不同外,序列和基因结构完全相同,这种基因组中的串联复制(segmental duplication,SD)导致数量上的扩增也间接证明了TCP基因在进化过程中的扩展。除了基因组复制中涉及的SD现象,还有区段复制(tandem duplication,TD)、染色体片段的交换、基因的加倍等情况也会导致基因组的持续扩张,而这些必然会促进表型和功能上的多样化。除此之外,根据本文对达尔文进化压力选择的分析可知,与小麦构成直系同源基因对的大麦基因数量比T.urartu和Aegilopstauschii这两个二倍体供体物种中的基因要多。除了已知大麦的基因组比它们的大之外,还可能因为小麦在经历全基因组复制(WGD)事件后导致TCP基因的数量增加,而两个二倍体物种的基因组变化不大。
相似的蛋白结构预示着其功能可能具有相似性,通过与其他相近物种的序列分析可以预测小麦TCP基因的功能。比如:miR319通过与ClassⅡ类TCP中的CIN亚类基因结合,以转录后负调控的方式调控基因的表达,在拟南芥和水稻中的TCP基因中均有该小RNA的靶向序列[17]。将位于同一CIN家族的TaTCP分析后发现, TaTCP16-A、 TaTCP16-B和 TaTCP16-D具有小麦tae-miR319的结合位点,而这三个部分同源基因是否存在功能上的冗余还有待进一步验证。不过从进化压力指标计算中发现, TaTCP16-A是经过正向选择的结果,说明这一基因在进化中起着积极的作用;或从热图中也可以看出, TaTCP16-A在非生物胁迫中的表达量变化比较显著,这些证据都间接证明该基因可以在以后研究的过程中优先考虑。
对于TCP基因参与非生物胁迫下的调控将是研究的热点之一。Wang等[18]研究发现,miR319可以负向调控水稻中 OsPCF6和 OsTCP21的表达来增加植株对低温的防御。Mukhopadhyay等[19]发现,过表达 OsTCP19可以通过调控多种激素途径提高水稻的耐盐和耐旱性。本研究发现,TCP的三个部分同源基因在小麦不同生育时期的表达量是不同的,每个基因都在特定生育时期中对耐热胁迫发挥主要作用,暗示 TaTCP5基因在抵御热胁迫中具有不同的生物学功能。结合环形进化树得知, OsTCP19和 TaTCP5都位于CIN亚家族,推测 TaTCP5基因可以通过与不同的激素信号途径如ABA、JA通路中的关键基因结合从而提高热响应,而由于该基因没有预测到有miRNA的直接靶标,所以是否有转录水平的调控难以阐明。 TaTCP5在同一生育时期,35 ℃的胁迫下要比42 ℃下的表达量高,可能是该基因对热胁迫有一定的防御限度,或者在42 ℃胁迫下, TaTCP5不是主要参与热响应的基因。而TCP基因通过对耐热的响应是否可以提高其他胁迫下的交叉适应性,或者TCP不同亚家族基因之间是否对耐热性存在交叉调控的机制,这些都需要进一步验证。
猜你喜欢
军事文摘(2022年16期)2022-08-24
基层中医药(2022年4期)2022-07-22
作文小学高年级(2022年5期)2022-06-16
环境卫生工程(2021年4期)2021-10-13
汉字汉语研究(2021年2期)2021-08-30
今日农业(2021年11期)2021-08-13
疯狂英语·新读写(2021年2期)2021-02-25
中国生殖健康(2020年4期)2020-12-09
中西医结合肝病杂志(2020年2期)2020-10-27
汉字汉语研究(2019年2期)2019-08-27
