
党参CpSUC4基因全长克隆及表达分析
2018-12-12 09:36:00郑庆红李晶吉姣姣李建宽高建平
山西大学学报(自然科学版) 2018年4期
郑庆红,李晶,吉姣姣,李建宽,2,高建平,2*
(1.山西医科大学 药学院,山西 太原 030001;2.山西省道地药材资源开发工程技术研究中心,山西 太原 030001)
0 引言
党参(Codonopsis Radix)为桔梗科植物党参Codonopsispilosula(Franch.) Nannf.、素花党参C.pilosulaNannf. Var.modesta(Nannf.) L. T. Shen或川党参C.tangshenOliv.的干燥根,具有健脾益肺,养血生津等功效,是我国传统的补气药(《中国药典》2015版)。现代药理研究表明党参有效成分党参多糖具有增加红细胞含量,增强淋巴细胞免疫作用[1],治疗慢性阻塞性肺疾病[2],保护胃黏膜、抗胃溃疡[3]等功效。
蔗糖是植物多糖合成途径中重要的前体物质[4],其合成主要由植物“源端”,即叶组织经光合作用完成,然后装载入韧皮部的筛管伴胞复合体,通过共质体或质外体途径到达花、果实、根系等“库”器官[5-6]。在缺少胞间连丝的情况下,植物体中蔗糖由“源”到“库”的运输主要通过韧皮部质外体途径进行[7],在质外体途径中存在一种关键性的载体蛋白,即蔗糖转运蛋白(sucrose/H+cotransporter,SUC或sucrose transporter,SUT),其可利用ATP在质膜H+/ATPase所建立的质子动力势作用下产生跨膜质子梯度,使蔗糖与H+跨膜共转运[8-9]。
双子叶植物中蔗糖转运蛋白分为SUT1、SUT2、SUT4这3个亚族,均属寡聚糖/H+共转运家族[10]。SUT1亚族为双子叶植物所特有,主要定位于韧皮部筛分子细胞质膜或库组织的细胞质膜上,在源库组织中均有表达,主要参与蔗糖的装载和卸载[11-12]。SUT2亚族属于低亲和力-高转运能力的蔗糖转运蛋白,多不具有转运蔗糖的活性,且不能有效定位到质膜[13],牡丹PsSUT2基因主要在库组织,如根、老茎、叶、鳞芽(花瓣)中表达,尤以盛花期花瓣中表达量最高[7]。SUT4亚族属于低亲和-高转运能力的蔗糖转运蛋白,在蔗糖较高的条件下,能以较大速率跨膜转运蔗糖分子[14]。植物SUT4类蛋白主要参与调控蔗糖在细胞液泡中的积累。拟南芥中AtSUC4基因超量表达后叶片蔗糖含量降低,与AtSUC4促进液泡中蔗糖外排的作用相一致[15];烟草NtSUT4的超量表达可促进纤维素的合成[16],苹果MdSUT4定位于苹果愈伤组织原生质体的液泡膜上,在各组织中均有较高表达,且在果实发育中的表达水平与蔗糖含量呈负相关,过表达MdSUT4能降低蔗糖含量,提高类黄酮含量[10],推测其可能参与蔗糖从液泡膜中的外排。但关于党参中SUT4类蛋白的研究,目前尚未见报道。
鉴于蔗糖转运蛋白在糖生理中的重要作用,推测党参蔗糖转运蛋白基因可能在党参多糖代谢与调控中起重要作用。本文以陵川潞党参为试材,克隆党参蔗糖转运蛋白基因CpSUC4,进行生物信息学及原核表达分析,并通过基因组织表达模式分析,结合党参多糖含量分析,探讨其与党参多糖代谢的相关性,为进一步揭示其在党参多糖合成中的调控机制奠定基础。
1 材料与方法
1.1 材料
样品采自山西陵川,经山西医科大学药学院高建平教授鉴定为党参,于盛花期取党参根、茎、叶、花组织样品,液氮速冻后于-80℃超低温冰箱中保存备用。
1.2 CpSUC4基因全长cDNA的克隆
采用改良Trizol法提取党参根、茎、叶、花四组织总RNA[16],质量分数1%琼脂糖凝胶电泳检测其完整性。cDNA第一链合成按照Takara反转录试剂盒说明书进行。
根据转录组数据库中Nr功能注释为SUC4的Unigene序列,利用Primer5.0设计引物,以党参根系cDNA为模板,进行PCR扩增,上下游引物序列见表1。
PCR产物经质量分数为1.2%琼脂糖凝胶电泳分离后,采用胶回收试剂盒对目标片段进行回收,并连接至pMDTM18-T载体(Takara,Japan),重组质粒转化大肠杆菌DH5α感受态细胞,阳性重组质粒进行双向测序(北京六合华大基因科技股份有限公司)。

表1 试验所用引物序列
1.3 党参CpSUC4基因生物信息学分析
根据测序得到的党参CpSUC4基因cDNA序列,进行蛋白质和核苷酸数据库Blast搜索(NCBI:http:∥www.ncbi.nlm.nih.gov/),采用“ORF Finder”查找开放阅读框(open reading frame,ORF,http∥www.ncbi.nlm.nih.gov/gorf/gorf.html)。编码蛋白的理化性质预测、蛋白保守结构域、蛋白跨膜区预测、亚细胞定位预测、氨基酸序列相似性分析等均为在线分析,具体方法参照课题组前期研究[17]。
利用DNAMAN6.0软件进行氨基酸多重序列比对。通过MEGA5.1重建Neighbor-joining系统进化树,设置Bootstrap=1000。
1.4 党参CpSUC4基因的原核表达
根据测序所得CpSUC4基因序列设计带酶切位点的特异引物,扩增CpSUC4基因完整cDNA序列,引物序列见表1。
将纯化的PCR产物与pET-28a(+)分别进行BamHⅠ、HindⅢ双酶切,T4DNA ligase连接,热激法转化DH5α感受态细胞,涂布于含100 mg/L卡那霉素的LB固体培养基上,挑取阳性克隆,摇菌,提取质粒并酶切鉴定,获得重组质粒pET-28a-CpSUC4。转化重组质粒pET-28a-CpSUC4至大肠杆菌BL21(DE3)感受态细胞,以pET-28a空载体为对照。具体方法见文献[18]。
1.5 党参多糖含量测定
于党参盛花期分别取根、茎、叶、花四组织样品,低温(<60℃)烘干后,粉碎,进行多糖含量测定。多糖含量测定方法参照课题组前期研究[19]。
1.6 党参CpSUC4基因不同组织表达分析
以党参GAPDH基因为内参基因,采用qRT-PCR进行党参CpSUC4基因的组织表达模式分析。试验方法见文献[4]。qRT-PCR引物序列见表1。
2 结果与分析
2.1 党参CpSUC4基因全长克隆
根据转录组数据库中Nr注释为SUC4的Unigene序列,设计特异引物,扩增产生长度约为1 600 bp的片段(图1),测序获得1 832 bp的序列,该cDNA序列包含的开放阅读框(ORF)常1 488 bp,编码495个氨基酸,BLASTx分析表明其与多个物种中SUT相似性很高,可达74%~77%,因此将其定名为CpSUC4。
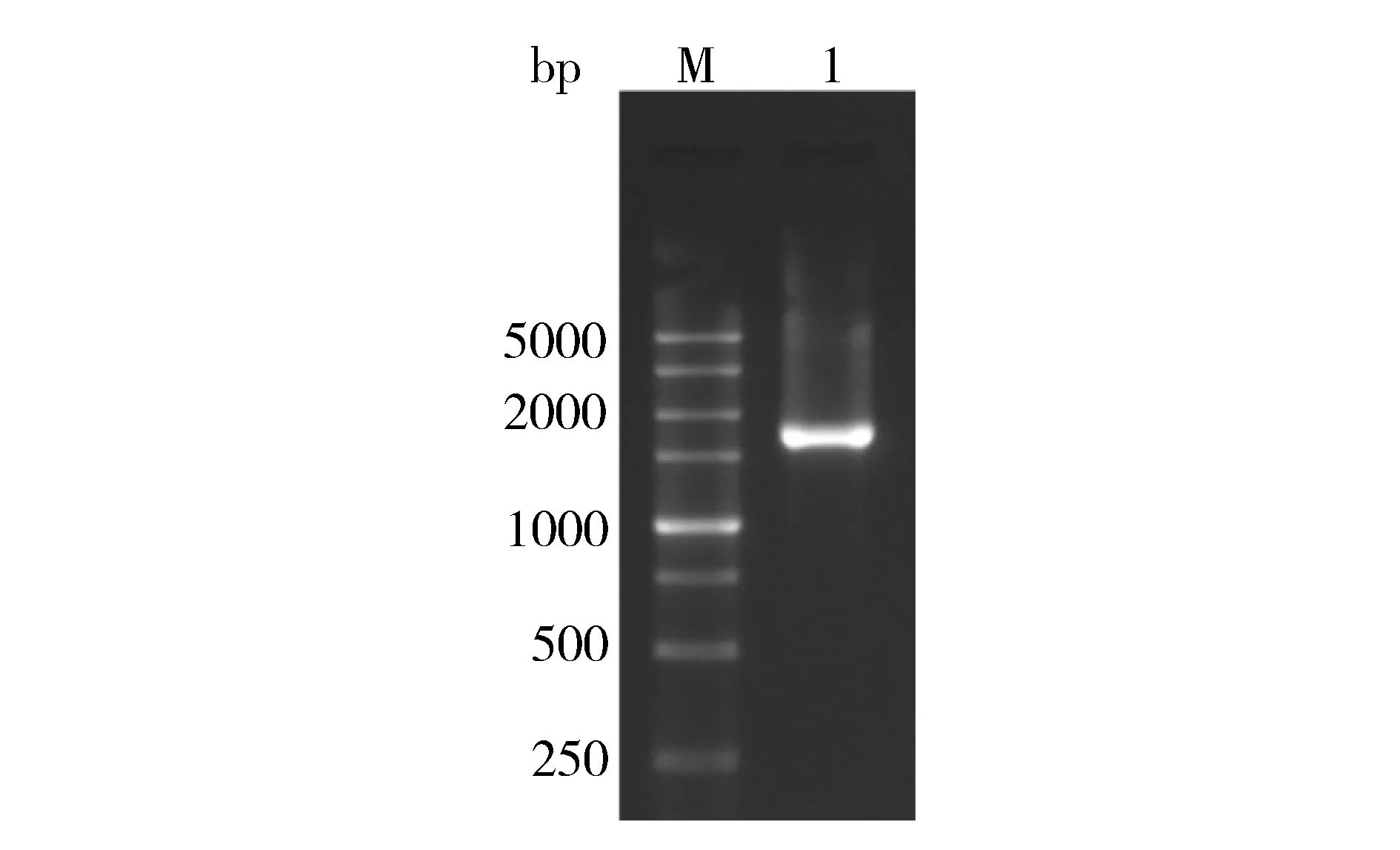
M-marker 1-CpSUC4gene M-DNA marker 1-CpSUC4基因Fig.1 Agarose gel electrophoresis analysis of the full length of the CpSUC4gene图1 党参CpSUC4基因全长cDNA电泳分析
2.2 党参CpSUC4生物信息学分析
NCBI中CpSUC4氨基酸序列Blast结果显示其与向日葵(Helianthusannuus)SUC4相似性达77%,与中粒咖啡Coffeacanephora、番茄(Lycopersiconesculentum)、北美假大麻(Datiscaglomerata)等的SUC4相似性达75%,并利用DNAMAN6.0软件对CpSUC4和其它物种的SUC4进行氨基酸序列的多重比对分析,结果表明CpSUC4与已知SUC4蛋白的氨基酸序列高度保守,相似性平均为73.35%,其中仅有5′端氨基酸序列同源性较低,第28-88位、第409-464位氨基酸序列高度一致(图2)。
保守结构域分析结果显示CpSUC4蛋白含有一个MFS(Major facilitator superfamily)功能域,属于GPH超家族(GPH family sucrose/H+symporter)和MFS蛋白家族,具备高等植物蔗糖转运蛋白的典型结构域特征(图3)。

Fig.2 Multiple alignment of the CpSUC4 deduced amino acid sequence with SUC4 in other plants图2 党参CpSUC4与其他植物SUC4氨基酸序列比对

Fig.3 Prediction of the conserved region of the CpSUC4deduced amino acid sequence图3 党参CpSUC4氨基酸保守结构域分析
为分析党参CpSUC4基因所编码蛋白的进化关系,从GenBank数据库中选取不同科共19个物种的SUT/SUC氨基酸序列,构建党参CpSUC4蛋白系统进化树。结果显示,党参CpSUC4与菊科植物向日葵、莴苣、菊苣蔗糖转运蛋白亲缘关系较近,可聚为一支(图4)。
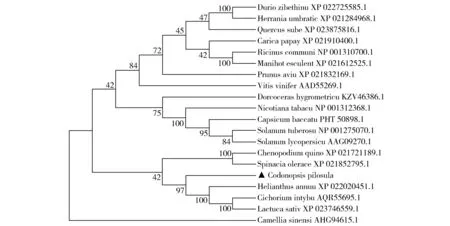
Fig.4 Clustering tree of SUC4s in plants图4 植物SUC4聚类树
亚细胞定位预测结果表明,内质网膜定位系数为0.850,过氧化物酶体定位系数为0.539,细胞质膜定位系数为0.440,线粒体内膜定位系数为0.100,说明CpSUC4蛋白主要定位在膜结构上,这与其主要参与蔗糖-质子的跨膜转运一致。TMHMM Server v.2.0跨膜结构预测结果(图5)表明,CpSUC4蛋白含有蔗糖转运蛋白所具有的典型的12次跨膜结构域(33-55位、65-87位、100-119位、134-153位、174-196位、221-240位、276-298位、321-343位、356-375位、390-412位、424-446位、461-483位氨基酸),N-端和C-端均位于细胞质的一侧。
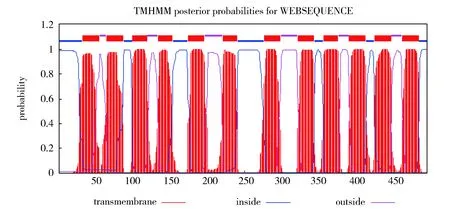
Fig.5 Prediction of transmembrane domain of CpSUC4 protein图5 CpSUC4蛋白跨膜结构域预测
蛋白理化性质预测分析发现党参CpSUC4分子式为C2487H3833N637O654S22,总原子数为7633。相对分子质量为53.83 kDa;理论等电点(pI)为8.55,带正电残基(Arg+Lys)为34,带负电残基(Asp+Glu)为30。该蛋白的不稳定系数为38.70,表明该蛋白质稳定。脂肪系数为107.74,亲水性系数为0.556。
2.3 党参CpSUC4基因表达与党参多糖合成相关性分析
为了分析党参CpSUC4基因表达与党参多糖合成的相关性,对党参不同组织党参多糖含量及CpSUC4基因表达量进行了测定,结果发现党参根系中多糖含量显著高于其他组织(P<0.05),花次之,叶中多糖含量最低,各组织间多糖含量差异显著(P<0.05)。而基因表达分析显示,党参根和花中CpSUC4基因表达量最高,二者差异不显著(P<0.05),而茎和叶中基因表达量最低,显著低于花中基因表达量(P<0.05)(图6)。
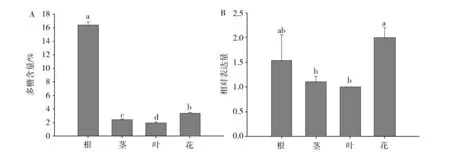
Fig.6 Analysis of the polysaccharide content (A) and the CpSUC4 gene expression (B) in Codonopsispilosula Data represent the means ± SD from three independent experiments. Statistically significant differences between means were determined using Fisher’s LSD test (P<0.05)图6 党参不同组织多糖含量(A)及CpSUC4基因表达(B)分析不同小写字母表示0.05水平差异显著
2.4 党参CpSUC4基因原核表达分析
为了分析CpSUC4基因是否能够编码活性蛋白,构建pET-28a-CpSUC4原核重组表达载体,转化大肠杆菌BL21(DE3)后进行IPTG诱导表达,结果显示重组CpSUC4表达载体可表达产生相对分子质量约为60 kDa特异性融合蛋白,而对照(不带外源基因的载体pET-28a)则不表达该蛋白(图7)。
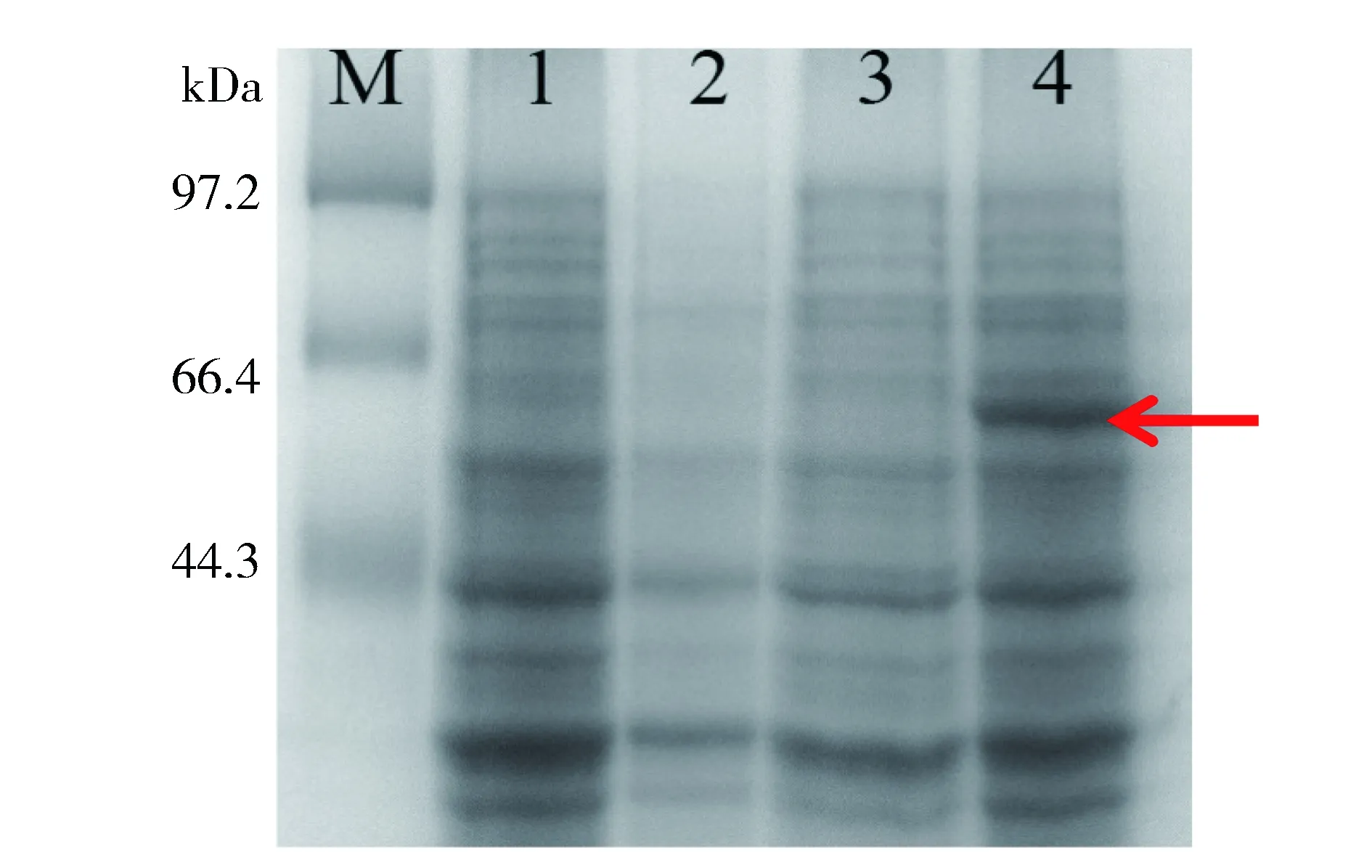
M-Marker;1-含pET-28a的菌未诱导;2-含pET-28a的菌诱导; 3-含pET-28a-CpSUC4的菌未诱导;4-含pET-28a-CpSUC4的菌诱导M-Marker; 1-pET-28a without induction; 2-pET-28a induced; 3-pET-28a-CpSUC4 without induction; 4-pET-28a-CpSUC4inducedFig.7 SDS-PAGE analysis of pET-28a-CpSUC4 expressed products图7 pET-28a-CpSUC4重组表达蛋白的SDS-PAGE分析
3 讨论
党参为中国传统补气药,而党参多糖作为党参有效成分之一,具有多种药理活性,因此通过分子水平研究其生物合成机制具有重要意义。本文根据克隆得到的党参SUC4基因序列,进行核苷酸和预测氨基酸序列分析,结果显示CpSUC4与其他植物SUC4具有很高的相似性,CpSUC4具有SUC/SUT所共有的MFS功能域,而MFS超家族常含有12个高度保守的跨膜结构域,只在N端和C端存在高变异区,最保守的跨膜区域位于第1、2和11跨膜区[6],而CpSUC4氨基酸序列高度一致的区域位于第28~88位、409~464位氨基酸,分别位于第1(33~55位)、2(65~87位)、11(424~446位)跨膜区。由于蔗糖转运蛋白基因在高等植物中具有高度保守性,亲缘关系越近,蔗糖转运蛋白的功能越相似[20]。党参薄壁细胞中含有大量菊糖[21],其为菊科植物特征性成分,有研究者推测其与菊科有亲缘关系,本试验系统进化树分析发现CpSUC4与菊科向日葵、菊苣、莴苣等SUC/SUT亲缘关系最近,这也为进一步研究菊科与桔梗科的亲缘关系提供了理论依据。蛋白理化性质预测CpSUC4大小为53.83 kDa,而原核表达分析显示CpSUC4融合表达蛋白大小为60kDa左右,与预测目标大小基本一致。
亚细胞定位预测显示CpSUC4属于液泡膜蔗糖转运蛋白,为植物所特有的载体蛋白,主要在蔗糖进出韧皮部和库器官、蔗糖的供给与贮藏、转运与调控等生理过程中发挥重要作用[10]。拟南芥中AtSUC4定位于液泡膜,参与液泡膜蔗糖的外运[13],同样,桃液泡膜PpSUT4也参与蔗糖从液泡中的外排[21],因此推测党参CpSUC4可能参与蔗糖从液泡中的外排。
CpSUC4基因在党参根、茎、叶、花中均有表达,说明CpSUC4基因可能具有双重功能,即韧皮部蔗糖的卸载及装载。杏PaSUC4基因同样在源库组织中均表达,但是在源器官如成龄叶、新梢中表达量最高[22],赵婷婷[23]对甘蔗的SUC4进行了定量分析,结果是在叶中表达量最高,在根中最低。本文的研究结果表明,CpSUC4在“源”器官叶中表达量最低,显著低于“库”器官花,推测CpSUC4主要参与“库”器官韧皮部蔗糖的卸载。
本课题组及以往研究显示,党参多糖合成的前体物质包括葡萄糖、果糖、阿拉伯糖、甘露糖等单糖,而这些单糖尤其是葡萄糖和果糖主要由蔗糖分解产生[4],因此蔗糖的代谢活动是党参多糖及其他以蔗糖为前体物质的有效成分合成的主要影响因子之一。苹果MdSUT4的超量表达可能促进蔗糖从液泡中外排至细胞质中,为类黄酮的合成提供了大量的前体物质,从而促进了类黄酮的合成,而降低了蔗糖含量[10]。多糖类物质如果聚糖主要在液泡中合成,且果聚糖合成关键酶如1-SST、1-FEH等均定位于液泡中[24],而蔗糖的外排会导致多糖合成前体物质的减少,从而降低多糖含量。本试验表明:盛花期党参多糖在根系中含量显著高于其他组织,而CpSUC4基因表达量在“源”器官叶中表达量最低,而蔗糖的代谢活动主要在细胞质中进行[25],因此推测CpSUC4表达量高,则会促进蔗糖向“库”器官如根、花等组织运输,而CpSUC4的液泡外排作用会使得韧皮部细胞质中蔗糖的大量积累,从而促进多糖合成,这也正与党参根、花等“库”器官多糖含量显著高于叶等“源”器官相符合。鉴于党参因此通过党参CpSUC4在糖类物质代谢及运输过程中的重要性,本文CpSUC4基因的克隆,可为进一步研究其在党参多糖生物合成中的调控机制提供物质基础,为完善党参多糖代谢途径奠定理论基础。
猜你喜欢
Natural Products and Bioprospecting(2021年2期)2021-04-10 01:42:56
广西糖业(2020年3期)2020-09-25 08:32:44
中国农业科技导报(2020年1期)2020-03-15 00:45:26
中国真菌学杂志(2019年1期)2019-01-23 09:04:20
中成药(2018年4期)2018-04-26 07:12:58
中成药(2018年1期)2018-02-02 07:20:31
浙江工业大学学报(2017年5期)2018-01-22 02:03:33
中成药(2017年10期)2017-11-16 00:50:06
中国糖料(2016年1期)2016-12-01 06:49:06
中国果业信息(2015年11期)2015-01-23 19:19:58
